

Доклады Российской академии наук. Науки о жизни, 2022, T. 505, № 1, стр. 288-293
ИММУНОГЕМАТОЛОГИЧЕСКИЕ ПАРАМЕТРЫ ЭКТОТЕРМНЫХ АМФИБИОНТОВ ФАУНЫ СРЕДНЕГО УРАЛА: СИБИРСКОГО УГЛОЗУБА (SALAMANDRELLA KEYSERLINGII DYBOVSKY, 1870, CAUDATA) И ОЗЕРНОЙ ЛЯГУШКИ (PELOPHYLAX RIDIBUNDUS PALLAS, 1771, ANURA)
Л. А. Ковальчук 1, *, Л. В. Черная 1, В. А. Мищенко 1, Д. Л. Берзин 1, академик РАН В. Н. Большаков 1
1 Федеральное государственное бюджетное учреждение науки Институт экологии растений и животных Уральского отделения Российской академии наук
Екатеринбург, Россия
* E-mail: kovalchuk@ipae.uran.ru
Поступила в редакцию 03.03.2022
После доработки 09.04.2022
Принята к публикации 11.04.2022
- EDN: HQXNHJ
- DOI: 10.31857/S2686738922040096
Аннотация
Впервые представлены результаты сравнительного анализа гематологических параметров эктотермных амфибионтов фауны Среднего Урала – сибирского углозуба (S. keyserlingii) и озерной лягушки (P. ridibundиs). Показана видовая специфика иммунной защиты изученных видов по содержанию гранулоцитов и агранулоцитов (р < 0.001). Высокий лимфоцитарный профиль (73.3–76.1%) теплоустойчивой озерной лягушки обеспечивает активацию механизмов приобретенного адаптивного иммунитета. У сибирских углозубов, адаптированных к условиям низко отрицательных температур, присутствует набор неспецифических лейкоцитов (39.3–44.4%). В сравнении с озерными лягушками сибирские углозубы характеризуются более высоким развитием врожденного иммунитета.
Сокращающаяся численность популяций и видового разнообразия земноводных нарушает устойчивость сообществ водных и наземных экосистем в различных регионах планеты [1–3]. На территории горного Урала в его естественных и антропогенных ландшафтах обитают экологически контрастные по температурной толерантности виды амфибий из отряда Anura – озерная лягушка (Pelophylax ridibundus Pallas, 1771) и из отряда Caudata – сибирский углозуб (Salamandrella keyserlingii Dybovsky, 1870) [4]. При исследовании механизмов гомеостаза сибирского углозуба и озерной лягушки нами показана видовая специфика термостабильного аминокислотного спектра плазмы крови изученных амфибий, поддерживающая их выживание и эврибионтность в широком температурном диапазоне [5]. Также отмечено присутствие в крови свободных аминокислот, обладающих иммуномодулирующими свойствами и участвующих в регуляции гемопоэза [5]. Известно, что в эволюционном аспекте роль системы крови позвоночных определяется как решающая при формировании резистентности организма к воздействию биотических и абиотических факторов среды (температура, гипоксия, токсичные химические вещества, паразитарные инфекции, стресс) [6–8]. Помимо этого, система крови у представителей герпетофауны достаточно высоко организована, а состав и морфология лимфоидной системы не отличаются от данных имеющихся научных источников по исследованию позвоночных [9–14]. Это позволяет считать, что адаптивная стратегия длительного пребывания сибирского углозуба (СУ) в условиях низко отрицательных температур (–30°С…–45°С) и толерантность озерной лягушки (ОЛ) к экстремально положительным температурам (+28°С…+40°С) несомненно обусловливает выживание амфибионтов [5, 15, 16]. Однако основная информация о механизмах иммунной защиты природных популяций хвостатых и бесхвостых амфибий весьма ограничена, а реакции адаптивного иммунитета и участие неспецифических иммунных ответов в регуляции физиологических процессов практически не исследованы [6].
Цель работы – сравнительный анализ иммуногематологических параметров периферической крови эктотермных амфибионтов природных популяций фауны Среднего Урала: сибирского углозуба (S. кeyserlingii) и озерной лягушки (P. ridibundиs).
Экспериментальные группы животных представлены половозрелыми самцами S. кeyserlingii (n = 20) и P. ridibundиs (n = 20), отловленными на территории Свердловской области (56°42′ с.ш. 61°20′ в.д.) в весенний (первая декада мая) и летний (последняя декада июля) сезоны 2019–2020 гг. Отлов амфибионтов в нерестовых водоемах проводился с помощью водного сачка; сбор особей СУ в летний период осуществляли вручную в наземных укрытиях. Животных без признаков заболеваний доставляли в лабораторию в отдельных контейнерах с влажным мхом в день отлова. Сибирский углозуб S. кeyserlingii включен в Красную Книгу Среднего Урала [17]. Статус – III категория. Редкий, малочисленный вид на периферии ареала. Встречается повсеместно, но спорадически. Экспериментальные особи СУ отловлены на затененных участках прибрежной части Калиновского пруда, расположенного в особо охраняемой лесопарковой зоне городской агломерации. Температура воды в весенний период составляла +11°С, в летний +17°С при среднесуточной температуре воздуха +15°С и +21°С соответственно. Взрослые особи СУ всю жизнь проводят на суше в прибрежной полосе нерестового водоема, за исключением короткого периода размножения. Длительного пребывания на солнце не переносят и при температуре в тени около +27°С погибают. Следует отметить, что большинство видов амфибий не способны адаптироваться к обитанию в теплых водоемах, тем не менее исследователи отмечают обитание озерной лягушки (инвазивный вид на Урале) в термальных водоемах, где температура воды достигает и до +30°С [4]. Опытные особи ОЛ отловлены в неглубоких заводях р. Тагил, где температура воды весной составляла в среднем +21°С, а летом +27°С и выше, при среднесуточной температуре воздуха +14°С и +19°С соответственно. Зимой температура воды исследуемого водотока не опускается ниже +10°С…+12°С, и обитающие в нем ОЛ характеризуются отсутствием зимней спячки. Показатели периферической крови определяли с помощью гематологического анализатора “BC-5800” (Mindray, Китай). Лейкоцитарную формулу рассчитывали на 100 лейкоцитов в мазках крови, окрашенных по Романовскому-Гимзе. На основании лейкоцитарной формулы определяли интегральный лейкоцитарный индекс (ИСЛ – соотношение гранулоцитов и агранулоцитов в отн. ед.), позволяющий оценить модуляцию эффекторных механизмов иммунной системы и степень напряженности компенсаторных процессов в организме. Результаты обработаны с использованием пакета лицензионных прикладных программ “Statistica v. 10.0”. Метод главных компонент (PCA) реализован посредством статистической среды R (R 3.1.2, пакеты “Vegan и “Ade4”) [18].
Исследования периферической крови амфибионтов показали статистически значимое повышенное содержание гемоглобина в 1.5 раза у СУ (59.8 ± 3.3 г/л) в сравнении с ОЛ (40.2 ± 2.5 г/л) (р = 0.04). Показатели содержания гемоглобина соответствуют значениям, приведенным в работах отечественных исследователей [10, 14, 19]. Превышение содержания тромбоцитов в 3.5 раза (41.7 ± 5.8 Г/л) (р = 0.01) и двукратное повышение тромбокрита (РСТ = 0.02%) связаны с возросшей долей объема цельной крови, занимаемой тромбоцитами, участвующими в иммунных реакциях у ОЛ в сравнении с СУ. У сибирского углозуба в весенний и летний периоды отмечено высокое содержание лейкоцитов (77.0 ± 7.5 Г/л), что в 2.4 раза больше, чем у ОЛ (р = 0.001). Лейкоцитарный состав крови ОЛ и СУ, как и у всех позвоночных, представлен двумя группами клеток: гранулоцитами (нейтрофилы, эозинофилы, базофилы), определяющими реакции врожденного иммунитета, и агранулоцитами (моноциты, лимфоциты), ответственными за реакции адаптивного иммунного ответа [6, 20]. В весенний и летний периоды лейкоцитарная формула периферической крови исследуемых амфибий имеет лимфоидный профиль: у ОЛ (69.1–69.8%) и у СУ (52.4–56.7%) (табл. 1). Выявлены видовые различия по количественному содержанию лимфоцитов, гранулоцитов и агранулоцитов, отражающих взаимосвязь эффекторных механизмов иммунной системы генетически детерминированных видов амфибий (р < 0.001) (табл. 1).
Таблица 1.
Параметры лейкоцитарной формулы самцов озерной лягушки (Pelophylax ridibundus) и сибирского углозуба (Salamandrella keyserlingii). Среднее арифметическое (X M boot), ошибка среднего (SEboot) и доверительный интервал ([95% CIboot]) бутстреп-распределения
| Показатели, % | Вид | I. Весна ОЛ (n = 8) СУ (n = 11) |
II. Лето ОЛ (n = 12) СУ (n = 9) |
Tukey’s test (p-value) | |
|---|---|---|---|---|---|
| Нейтрофилы | ОЛ | 24.1 ± 1.4 [21.1–26.8] |
17.1 ± 0.5* [16.2–18.0] |
I–II (< 0.001) | Вид I: < 0.001 II: < 0.001 |
| СУ | 40.6 ± 1.0@ [38.8–42.6] |
33.4 ± 0.6*@ [32.3–34.4] |
I–II (< 0.001) | ||
| – юные | ОЛ | 3.3 ± 0.3 [2.6–4.0] |
5.8 ± 0.4* [5.2–6.5] |
I–II (< 0.001) | Вид I: < 0.001 II: < 0.001 |
| СУ | 6.2 ± 0.3@ [5.7–6.7] |
9.3 ± 0.4*@ [8.6–10.0] |
I–II (< 0.001) | ||
| – палочкоядерные | ОЛ | 9.2 ± 0.7 [7.9–10.5] |
6.9 ± 0.6* [5.8–8.0] |
I–II (0.05) | Вид I: 0.28 II: < 0.001 |
| СУ | 10.1 ± 0.4 [9.3–10.9] |
17.1 ± 0.3*@ [16.4–17.8] |
I–II (< 0.001) | ||
| – сегментоядерные | ОЛ | 11.5 ± 1.0 [9.7–13.7] |
4.2 ± 0.2* [3.7–4.6] |
I–II (< 0.001) | Вид I: < 0.001 II: < 0.001 |
| СУ | 23.8 ± 0.6@ [22.7–24.9] |
7.1 ± 0.5*@ [6.1–8.0] |
I–II (< 0.001) | ||
| Эозинофилы | ОЛ | 2.7 ± 0.3 [2.1–3.2] |
6.9 ± 0.5* [5.9–7.9] |
I–II (< 0.001) | Вид I: 0.05 II: 0.10 |
| СУ | 3.8 ± 0.2@ [3.4–4.3] |
5.9 ± 0.3* [5.4–6.3] |
I–II (< 0.001) | ||
| Моноциты | ОЛ | 4.2 ± 0.6 [3.0–5.4] |
6.3 ± 0.5* [5.4–7.3] |
I–II (0.001) | Вид I: 0.15 II: < 0.001 |
| СУ | 3.3 ± 0.2 [2.9–3.7] |
3.6 ± 0.2@ [3.3–3.9] |
I–II (0.60) | ||
| Лимфоциты | ОЛ | 69.1 ± 0.9 [67.5–71.1] |
69.8 ± 0.5 [68.8–70.8] |
I–II (0.60) | Вид I: < 0.001 II: < 0.001 |
| СУ | 52.4 ± 1.0@ [50.4–54.4] |
56.7 ± 0.8*@ [55.2–58.1] |
I–II (0.001) | ||
* – статистически значимые сезонные различия: I и II (p < 0.05); @ – видовые различия (p < 0.05); ОЛ – озерная лягушка, СУ – сибирский углoзуб.
Note: * – statistically significant seasonal differences: I and II (p < 0.05); @ – species differences (p < 0.05); LF – Lake frog, SS – Siberian salamander.
Периферическая кровь исследованных видов характеризуется преобладанием агранулоцитов, обеспечивающих иммунный надзор и избирательную реактивность организма в весенний и летний сезоны: у ОЛ – 73.3–76.1% и у СУ – 55.7–60.3% (р < 0.001). Лимфоцитарно-гранулоцитарный состав периферической крови амфибий характеризуется невысокой долей гранулоцитов у ОЛ (24.0–26.7%) в сравнении с СУ (39.3–44.4%) (р < 0.001). В весенний и летний периоды в периферической крови СУ доля гранулоцитов в 1.7 раза выше, чем у ОЛ (p < 0.001), а доля агранулоцитов на 30% выше у ОЛ (табл. 1). Повышенный уровень нейтрофилов (р < 0.001) периферической крови СУ обеспечивает неспецифическую защиту организма как от токсических воздействий, так и от патогенных инфекций. Отмечена также разнонаправленность сезонной динамики палочкоядерных нейтрофилов в крови ОЛ и СУ (p < 0.001) (табл. 1). У летних особей ОЛ снижено содержание сегментоядерных нейтрофилов в 2.7 раза и у СУ в 3.3 раза.
Активация эозинофильного гранулоцитопоэза в крови самцов ОЛ в 2.6 раза (р < 0.001) и у СУ в 1.6 раза (р < 0.001) в летний период свидетельствует о единых механизмах, определяющих способность животных к избирательной адаптивной стратегии как по антимикробной, так и по противогельминтной иммунной защите [13, 14]. В периферической крови половозрелых самцов СУ в весенний и летний периоды отмечено значимое содержание участников клеточных воспалительных реакций: базофильных лейкоцитов (0.08–0.09%) и их отсутствие в крови ОЛ. Межвидовые различия по содержанию моноцитов у исследуемых гидробионтов весной отсутствуют (р = 0.15) и наблюдаются в летний период (р < 0.001). Известно, что повышенная активация моноцитов, выполняя фагоцитарную роль, продуцирует провоспалительные цитокины – эндогенные регуляторы гемопоэза и клеточно-опосредованного иммунного ответа [7, 11, 20].
По-видимому, в ответ на действие эндотоксинов среды обитания моноциты ОЛ (в отличие от СУ) в летний период усиливают выработку провоспалительных цитокинов, активируя систему естественного иммунитета (p < 0.001). Проведенный расчет ИСЛ подтвердил межвидовые различия в лейкоцитарном профиле исследованных животных. Высокий показатель ИСЛ (0.65–0.80) для СУ, по сравнению с ИСЛ (0.32–0.36) для ОЛ подтверждает более выраженную реактивность системы врожденного иммунитета, обеспечивающей неспецифическую срочную защиту организма от патогенных антигенов среды обитания (p < 0.001). Наряду с едиными закономерностями, выявленными в путях поддержания гомеостаза у двух видов амфибий: озерной лягушки и сибирского углозуба, показана и определенная стратегия разнонаправленности в механизмах аварийного регулирования иммунитета.
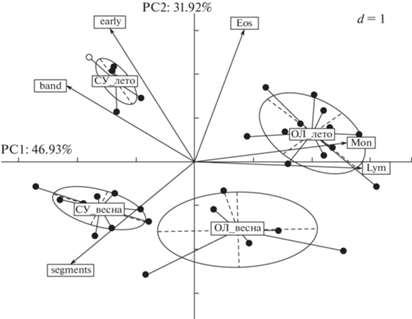
Использование метода главных компонент (РСА) позволило визуализировать видовую и сезонную специфику лейкоцитарного состава крови СУ и ОЛ, подтверждая результаты представленного выше статистического анализа. Показано, что 46.93% общей дисперсии параметров крови приходится на первую главную компоненту (РС1), 31.92% – на вторую главную компоненту (РС2) (рис. 1). Первая (РС1) и вторая (РС2) главные переменные обусловливают значимые различия амфибионтов по основным параметрам лейкоцитарной крови. Наибольший вклад в межвидовую изменчивость показателей крови по РС1 вносят лимфоциты (30.43%), моноциты (25.01%), палочкоядерные нейтрофилы (17.68%), сегментоядерные нейтрофилы (16.52%). Их коэффициенты корреляции с РС1 составили соответственно: 0.93, 0.84, –0.71, –0.68 (р < 0.001). Первая главная компонента по этим переменным четко выделяет в самостоятельные группы как особей резистентного вида к холоду (СУ), так и теплоустойчивого вида (ОЛ) (рис. 1). Со второй главной компонентой РС2 высоко коррелируют эозинофилы (0.80), юные нейтрофилы (0.81), сегментоядерные нейтрофилы (–0.62). Вклад этих показателей в сезонную изменчивость лейкоцитарной формулы изучаемых видов амфибий составил 33.67, 34.18 и 20.29%, соответственно, что также подтверждает неоднородность особей исследованных двух экологически контрастных видов амфибий по данным показателям. Из рисунка видно, что все представленные данные обособлены в четыре самостоятельные группы, причем их наибольшая пространственная дифференциация обусловлена альтерацией лейкоцитарного состава крови в зависимости от сезона.
Рис. 1.
Результаты компонентного анализа лейкограммы (%) сибирского углозуба (СУ) и озерной лягушки (ОЛ) в разные сезоны года. PC1, PC2 – оси главных компонент, % – процент дисперсии данных, объясненных главной компонентой; стрелки отражают корреляцию главных компонент с исходными показателями; эллипсы представляют собой 95% доверительные области.
Таким образом, результаты проведенных исследований впервые позволили оценить видовую специфику эффекторных механизмов иммунной системы в регуляции физиологических процессов амфибионтов: озерной лягушки и сибирского углозуба, что обеспечивает выживание и эврибионтность исследованных видов в широком температурном диапазоне естественных и антропогенных ландшафтов Среднего Урала. Знание о наследственно обусловленных иммуногематологических параметрах гомеостаза видов, помимо несомненного теоретического интереса, имеют и прикладное значение при разработке природоохранных мероприятий в условиях быстрых преобразований среды обитания.
Список литературы
Strayer D., Dudgeon D. Freshwater biodiversity conservation: recent progress and future challenges // Science. 2010. V. 29. № 1. P. 344–358.
Global amphibian assessment. International Union for Conservation of Nature and Natural Resources. 2008. Cambridge, UK. (Available from: http://www.globalamphibians.org).
Stuart S.N., Chanson J.S., Cox N.A., et al. Status and trend of amphibian decline and extinction worldwide // Science. 2004. V. 306. P. 1783–1786.
Большаков В.Н., Вершинин В.Л. Амфибии и рептилии Среднего Урала. Екатеринбург; 2005.
Ковальчук Л.А., Черная Л.В., Мищенко В.А., и др. Аминокислотный спектр плазмы крови эндемичного и инвазивного видов земноводных фауны Урала // Доклады Российской академии наук. Науки о жизни. 2021. Т. 500. № 1. С. 478–482.
Хаитов Р.М. Иммунология: Структура и функции иммунной системы. М.: ГЭОТАР – Медиа. 2013.
Davis F.R., Maney D.L., Maers J.C. The use of leukocyte profiles to measure stress in vertebrates: a review for ecologists // Functional Ecology. 2008. V. 22. P. 760–772.
Ковальчук Л.А., Мищенко В.А., Черная Л.В., и др. Особенности иммуногематологических параметров перелетного (Vespertilio murinus Linnaeus, 1758) и оседлого (Myotis dasycneme Boie, 1825) видов рукокрылых фауны Урала // Доклады РАН. Науки о жизни. 2021. Т. 501. № 6. С. 49–52.
Fournier M., Robert J., Salo H.M., et al. Immunotoxicology of Amphibians // Applied Herpetology. 2005. V. 2. P. 297–309.
Силс Е.А. Сравнительный анализ гематологических показателей остромордой (Rana arvalis, Nilsson, 1842) и озерной (Rana ridibunda, Pallas, 1771) лягушек городских популяций // Вестник ОГУ. 2008. № 10 (92). С. 230–235.
Zimmerman L.M., Vogel L.A., Bowden R.M. Understanding the vertebrate immune system: Insights from the reptilian perspective // Journal Experimental Biology. 2010. V. 213. P. 661–671.
Arikan H., Cicek K. Hematology of amphibians and reptiles: a review // North-West. J. Zool. 2014. V. 10. № 1. P. 190–209.
Романова Е.Б., Соломайкин Е.И., Бакиев А.Г. и др. Лейкоцитарный состав крови Elaphe dione (Serpentes: Colubridae) заповедника “Оренбургский” (Россия) // Nature Cons. Res. Заповедная наука. 2018. № 3. С. 28–35.
Романова Е.Б., Шаповалова К.В., Рябинина Е.С. Лейкоцитарный состав крови и микроядра в эритроцитах амфибий загрязненных водных объектов Нижегородской области // Принципы экологии. 2018. № 2. С. 125–139.
Берман Д.И., Мещерякова Е.Н., Булахова Н.А. Экстремальные отрицательные температуры и потери массы сибирским углозубом (Salamandrella keyserlingii, Amphibia, Hynobiidae) // Доклады Академии Наук. 2016. Т. 468. № 5. С. 590–593.
Ляпков С.М. Озерная лягушка (Pelophylax ridibundus) в термальных водоемах Камчатки // Зоол. журн. 2014. Т. 93. № 12. С. 1427–1432.
Красная книга Среднего Урала. Екатеринбург; Изд-во Урал. Ун-та.1996.
Chessel D., Dufour A.B., Thioulouse J. The ade 4 package-I: One-table methods // R News. 2004. № 4. P. 5–10.
Вершинин В.Л., Вершинина С.Д. Сравнительный анализ содержания гемоглобина у четырех видов бесхвостых амфибий Уральской горной страны // Доклады РАН. 2013. № 4. С. 488–491.
Coico R., Sunshine G., Benjamini E. Immunology. A Short Course. Hoboken. NJ: Wiley-Liss Publications. 2003.
Дополнительные материалы отсутствуют.
Инструменты
Доклады Российской академии наук. Науки о жизни