

Доклады Российской академии наук. Науки о жизни, 2022, T. 506, № 1, стр. 396-405
РАННЕПЛЕЙСТОЦЕНОВЫЙ ПОДКОВОНОС RHINOLOPHUS MACRORHINUS CIMMERIUS SUBSP. NOV. (RHINOLOPHIDAE, CHIROPTERA) ИЗ ПЕЩЕРЫ ТАВРИДА В КРЫМУ
Академик РАН А. В. Лопатин 1, *
1 Палеонтологический институт им. А.А. Борисяка Российской академии наук
Москва, Россия
* E-mail: alopat@paleo.ru
Поступила в редакцию 15.05.2022
После доработки 27.05.2022
Принята к публикации 01.06.2022
- EDN: STDVNY
- DOI: 10.31857/S2686738922050171
Аннотация
Из нижнеплейстоценовых отложений пещеры Таврида в центральном Крыму описаны многочисленные остатки (неполный череп, черепные и нижнечелюстные фрагменты, изолированные зубы) крупного подковоноса из группы Rhinolophus ferrumequinum, отнесенного к новому подвиду Rhino-lophus macrorhinus cimmerius subsp. nov. По зубным признакам новый подвид менее специализирован, чем R. m. anomalidens Topál, 1979 из позднего виллафранка Центральной Европы, что подразумевает его происхождение от более древней формы, морфологически близкой R. m. macrorhinus Topál, 1963. Хорошая сохранность черепных структур позволила изучить следы небных валиков и строение носовых раковин R. macrorhinus cimmerius subsp. nov.
Раннеплейстоценовые мелкие млекопитающие из пещеры Таврида в центральном Крыму (Белогорский район, пос. Зуя) включают насекомоядных, зайцеобразных, грызунов [1, 2] и рукокрылых. Наиболее многочисленны остатки Rodentia и Chiroptera. Проведенное нами предварительное изучение показало присутствие в составе ассоциации летучих мышей из этого местонахождения представителей родов Rhinolophus (Rhinolophidae), Eptesicus, Plecotus и Myotis (Vespertilionidae).
В настоящей статье описываются черепные, нижнечелюстные и зубные остатки крупных подковоносов Rhinolophus macrorhinus Topál, 1963 из нижнеплейстоценовых отложений пещеры Таврида, собранные в 2018, 2020 и 2021 г. Хорошая сохранность черепных материалов позволила изучить следы небных валиков и строение носовых раковин; обычно эти структуры не попадают в область внимания исследователей ископаемых рукокрылых.
Возраст фаунистического комплекса пещеры Таврида определен в пределах позднего виллафранка, около 1.8–1.5 млн л.н. [1, 2].
Изученный материал хранится в Палеонтологическом институте им. А.А. Борисяка РАН (ПИН) в г. Москве. Размеры приведены в мм. Методика измерений черепа по [3]. Терминология строения зубов по [4]. О составе и эволюции группы Rhinolophus ferrumequinum в плейстоцене Европы см. [5–8].
Отряд Chiroptera Blumenbach, 1779
Семейство Rhinolophidae Gray, 1825
Подсемейство Rhinolophinae Gray, 1825
Род Rhinolophus Lacépède, 1799
Rhinolophus macrorhinus Topál, 1963
Rhinolophus macrorhinus cimmerius Lopatin, subsp. nov.
Название подвида – лат. киммерийский, от Киммерии, античного названия Крыма и всего Северного Причерноморья.
Голотип – ПИН, № 5644/940, неполный череп с правыми и левыми C1, P4–M3 (рис. 1а–1г, 4а); Россия, Республика Крым, Белогорский район, местонахождение Таврида (точка “Логово гиен”, шурф 3, глубина 130–140 см; сборы 2020 г.); нижний плейстоцен.
Рис. 1.
Rhinolophus macrorhinus cimmerius Lopatin, subsp. nov.: а–г – голотип ПИН, № 5644/940, неполный череп с правыми и левыми C1, P4–M3: а – с дорсальной стороны, б – с вентральной стороны, в – с правой латеральной стороны, г – с ростральной стороны; д – экз. ПИН, № 5644/936, лицевая часть черепа с правыми и левыми C1, P4–M3 с вентральной стороны; е – экз. ПИН, № 5644/803, лицевая часть черепа с левыми C1, P4–M2 и правыми P4–M2 с вентральной стороны; ж – экз. ПИН, № 5644/801, лицевая часть черепа с правыми C1, P2, P4–M3 и левыми C1, P4–M3 с вентральной стороны; з – экз. ПИН, № 5644/938, лицевая часть черепа с правыми и левыми P4–M3 с вентральной стороны; и – экз. ПИН, № 5644/810, левая верхнечелюстная кость с C1–M3 с окклюзиальной стороны; к–м – экз. ПИН, № 5644/822, правая нижнечелюстная кость с C1–M3: к – с лабиальной стороны, л – с окклюзиальной стороны, м – с лингвальной стороны; н–п – экз. ПИН, № 5644/821, правая нижнечелюстная кость с C1–M3: н – с лабиальной стороны, о – с окклюзиальной стороны, п – с лингвальной стороны; р – экз. ПИН, № 5644/851, левая нижнечелюстная кость без зубов с лабиальной стороны; Россия, Крым, пещера Таврида; нижний плейстоцен.

Описание (рис. 1–4). Размеры сравнительно крупные, как у современного R. ferrumequinum (Schreber, 1774). Череп относительно низкий, мозговая коробка широкая, отчетливо выпуклая латерально. Межглазничный промежуток узкий. Ширина черепа в скуловых дугах немного превосходит ширину в мастоидных отростках. Суставные впадины сравнительно небольшие.
Рис. 2.
Rhinolophus macrorhinus cimmerius Lopatin, subsp. nov., экз. ПИН, № 5644/936, лицевая часть черепа с правыми и левыми C1, P4–M3 с вентральной стороны (СЭМ): а – общий вид; б – костное небо; в – увеличенный фрагмент правой части небного гребня IV; г – увеличенный фрагмент правой части небного гребня IV в области разрушения (видна поверхность кости); Россия, Крым, пещера Таврида; нижний плейстоцен.

Рис. 3.
Rhinolophus macrorhinus cimmerius Lopatin, subsp. nov., экз. ПИН, № 5644/939, лицевая часть черепа с правыми P4–M3 и левыми M1–M3: а – с вентральной стороны; б – томограмма носовой полости, аксиальная проекция (медиальный и латеральный тяжи правого комплекса турбиналий удалены); в – томограмма левой половины носовой полости, сагиттальная проекция (в антеровентральной части комплекса турбиналий видны крупные костные тяжи, более длинный латеральный тяж сильно выступает дистально); г, д – изъятый из носовой полости правый латеральный тяж (СЭМ): г – с латеральной стороны, д – с медиальной стороны; Россия, Крым, пещера Таврида; нижний плейстоцен.
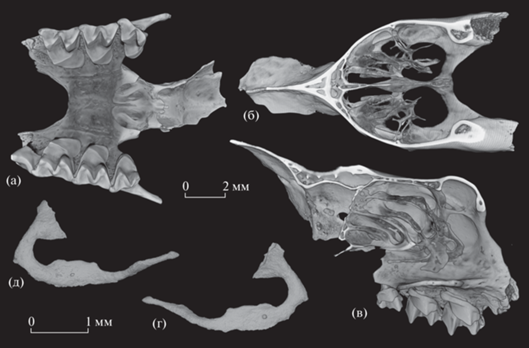
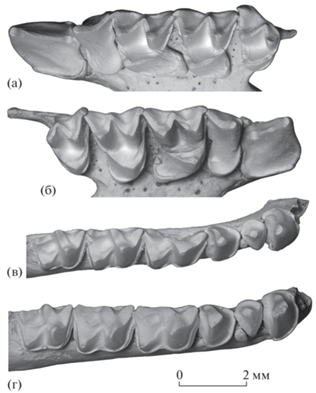
Рис. 4.
Rhinolophus macrorhinus cimmerius Lopatin, subsp. nov.: а – голотип ПИН, № 5644/940, левые С1 и P4–M3 с окклюзиальной стороны; б – экз. ПИН, № 5644/801, правые С1–M3 с окклюзиальной стороны; в – экз. ПИН, № 5644/822, правые C1–M3 с окклюзиальной стороны; г – экз. ПИН, № 5644/821, правые C1–M3 с окклюзиальной стороны; Россия, Крым, пещера Таврида; нижний плейстоцен.
Ростральный отдел черепа массивный (рис. 1а, 1б). Клыки широко расставлены. Носовое вздутие умеренно высокое, короткое, спереди не выступает за край носовой вырезки. Парные срединные вздутия хорошо развиты, но слабо обособлены друг от друга, парные боковые вздутия слабо обозначены. Надглазничные гребни четко выражены, ограниченная ими дистально и латерально ростральная (или лобная) впадина (расположенная позади срединных вздутий) умеренно углубленная. Сагиттальный гребень, тянущийся от места слияния надглазничных гребней к затылку, хорошо развит на всем протяжении и особенно высок на уровне передней части мозговой коробки (рис. 1в, 1г).
Подглазничное отверстие крупное, треугольной формы. Костный мостик, отделяющий подглазничное отверстие от глазницы, очень узкий и тонкий, палочковидный. Мелкое щелевидное слезное отверстие находится в дорсальном основании этого костного мостика, на уровне середины M2. Скуловая дуга тонкая, низкая, с очень легким дорсальным изгибом (рис. 1в).
Наименьшее расстояние между наиболее выступающим постеролингвальным краем полки гипокона M2 и задним краем верхнечелюстной кости примерно равняется ширине полки гипокона M2 или в полтора-два раза превосходит ее.
Небный мостик сравнительно короткий (длина без переднего выступа и заднего шипа составляет 31–36.5% от длины верхнечелюстного зубного ряда C1–M3), его передний край находится на уровне паракона, протокона или мезостиля M1, а задний край расположен на уровне мезостиля, метакона или метастиля M2 либо парастиля M3. Хоаны заметно расширены постеролатерально.
На некоторых экземплярах в пределах костного неба сохранились следы мягких тканей, позволяющие реконструировать наличие пяти слегка изогнутых дистально поперечных небных складок (или небных валиков, plicae palatinae transversae vel rugae palatinae). Эти следы имеют вид легких субпараллельных углублений, часто подчеркнутых осветлениями на поверхности кости в форме широких субпоперечных полос и более четких тонких линий на месте осевых частей небных валиков (экз. ПИН, №№ 5644/801, 803, 806, 816, 937, 938, 954, 956, 959, 963 и голотип).
На экз. ПИН, № 5644/936 остатки оснований небных складок сохранились в виде светлоокрашенного корковидного рыхловатого непрочного образования на костном небе, с шевронообразными поперечными утолщениями по осевым гребням отдельных валиков (рис. 1д, 2а, 2б). Латеральные края наиболее дистального из идентифицированных небных валиков располагаются у полок гипоконов левого и правого M2. Более мезиальные небные складки находятся соответственно на уровне середины M2, лингвального промежутка M1/M2, середины M1 и заднего края P4. Очевидно, еще два крупных передних небных валика располагались в пределах твердого неба спереди от небного мостика (валик I между клыками, валик II между противолежащими P4 или промежутками C1/P4), как у других представителей рода Rhinolophus [9, 10]. Таким образом, сохранившиеся на экз. ПИН, № 5644/936 следы относятся к небным валикам III (между промежутками P4/M1), IV (между средними частями противолежащих M1), V (между промежутками M1/M2), VI (между средними частями противолежащих M2) и VII (между полками гипоконов M2, у заднего края небного мостика). На экз. ПИН, №№ 5644/803, 816, 938 сохранились следы небных валиков III–VI, на экз. ПИН, № 5644/806 – небных валиков IV–VII, на прочих – только небных валиков V и VI (рис. 1б, 1е–1з).
Изучение корковидных образований, слагающих остатки небных валиков на экз. ПИН, № 5644/936, на сканирующем электронном микроскопе Tescan Vega II XMU в ПИН показало их слоисто-комковатую неупорядоченную ячеистую структуру (рис. 2в, 2г). Исследование данного образца методом энергодисперсионной рентгеновской спектроскопии на указанном СЭМ (программное обеспечение INCA 4.14, Oxford Instruments) продемонстрировало отсутствие качественных и количественных различий в химическом составе (при преобладании Ca, P и O) между веществом остатков небных валиков и костным веществом других частей черепа. Следует полагать, что описанные остатки небных валиков представляют собой фоссилизованные фрагменты соединительной ткани, слагавшей основания небных валиков при жизни животного [см. 9]. Внешне они соответствуют высохшим морфологическим структурам соединительных тканей твердого неба на черепах современных рукокрылых из остеологических коллекций.
Носовые раковины (турбиналии) внутри носовой полости имеют характерное для Rhinolophoidea (и уникальное среди Mammalia) строение, связанное с назофонацией (испусканием эхолокационных сигналов через ноздри), свойственной этим летучим мышам [11–13]. В антеровентральной части комплекса турбиналий каждой стороны обособлены два резко изогнутых и вытянутых дистально тонких костных тяжа – медиальный и латеральный (рис. 1г, 3). Более длинный латеральный тяж достигает уровня заднего края небного мостика (экз. ПИН, №№ 5644/801, 803, 937, 954), слегка выступает за него (экз. ПИН, №№ 5644/802, 938, 939; рис. 3а) или заметно выходит за пределы носовой полости и достигает уровня парастиля – паракона M3 (голотип; рис. 1б) или даже метакона M3 (экз. ПИН, № 5644/936; рис. 1д, 2а, 2б). Средняя часть горизонтальной порции латерального тяжа расширена дорсовентрально и имеет по одному миниатюрному отверстию на латеральной и медиальной сторонах (рис. 3г, 3д). Медиальный тяж существенно короче латерального и не выходит за край небного мостика (рис. 3в).
Ряд C1–P4 по длине в 1.5–1.7 раза уступает ряду M1–M3 (рис. 1б, 1д, 1ж, 4а, 4б).
Верхний клык С1 мощный, высокий, трехгранный, с резкими ребрами и слегка загнутой дистально вершиной (рис. 1в, 1г). Основание коронки трапециевидных очертаний. Лингвальная сторона длиннее лабиальной, прямая, с заметным постеролингвальным выступом. Цингулюм непрерывный, его высота положения и толщина изменчивы. На антеролабиальной стороне цингулюм резко изогнут в апикальном направлении. Постеролабиальный край основания коронки с глубокой вырезкой, в которой помещается коронка P2. Постеролингвальный край основания коронки слегка вогнутый (рис. 4а, 4б).
Рудиментарный верхний малый премоляр P2 однокорневой, с округлой плоской коронкой (рис. 1ж, 1и, 4б). Он смещен к лабиальному краю зубного ряда и не препятствует прямому контакту C1 и P4 (хотя обычно между этими зубами сохраняется небольшой промежуток).
Верхний большой премоляр P4 короткий и широкий, с мелким парастилем, развитым талоном и узким цингулюмом, который существенно расширяется в лингвальной части. Цингулюм прерывается в середине задней стороны основания коронки, на месте отчетливой задней вырезки (рис. 4а, 4б).
Верхние моляры M1 и M2 с широкой полкой гипокона, особенно мощной и сильно выступающей постеролингвально на M1 (рис. 1б, 1д–1з, 2а, 4а, 4б). Прецингулюм короткий и узкий, постцингулюм хорошо развитый. Задняя вырезка неглубокая, но отчетливая. M3 сравнительно слабо редуцированный, субтреугольный, с крупным парастилярным выступом. Метастиль отсутствует, метакон хорошо развит. Прецингулюм и постцингулюм рудиментарные, короткие и узкие.
Горизонтальная ветвь нижнечелюстной кости относительно длинная и низкая, с вогнутым вентральным краем и хорошо развитым подбородочным выступом (рис. 1к–1р). Подбородочное отверстие округлое, расположено под P2, задней частью C1 или границей C1/P2. Симфиз достигает уровня задней части C1. Нижнечелюстное отверстие крупное, овальное, продольно вытянутое. Восходящая ветвь длинная и низкая. Венечный отросток субтреугольный, по высоте приблизительно равен C1. Передний и задний края венечного отростка пологие, вершина слегка загнута назад. Сочленовный отросток длинный, суставный мыщелок широкий. Постдентальная длина нижнечелюстной кости (расстояние от M3 до заднего края суставного мыщелка) приблизительно равна длине ряда M1–M3. Угловой отросток мощный, длинный и сравнительно широкий, с тупым концом, направлен дистолатерально (рис. 1л, 1о).
Альвеолы двух нижних резцов тесно сближенные (рис. 4в).
Длина ряда C1–P4 приблизительно в 1.8–1.9 раза меньше длины ряда M1–M3 (рис. 4в, 4г).
Нижний клык C1 высокий, вертикальный. Основание коронки полукруглое, задний край слегка вогнутый. Цингулид непрерывный, узкий, формирует заметный постеролингвальный выступ.
Нижние малые премоляры представлены P2 и рудиментарным P3 (рис. 1к, 1л, 1н, 1о, 4в, 4г). В редких случаях P3 не развит, его альвеола отсутствует (рис. 1р).
P2 низкий, приблизительно вчетверо ниже C1. Коронка короткая и широкая, с округло-треугольной окклюзиальной поверхностью (лингвальная сторона заметно длиннее лабиальной) и узким цингулидом по всему основанию коронки.
P3 очень мелкий, однокорневой, столбиковидный, смещен лабиально от оси зубного ряда и не препятствует контакту P2 и P4. Дистально P3 прилегает к антеролабиальной стороне коронки P4, а антеродорсально частично перекрывается задней частью коронки P2. Коронка P3 овальная, поперечно вытянутая. Этот рудиментарный зуб участвует в окклюзии на поздних онтогенетических стадиях, при очень сильном стирании P2 (рис. 4г).
Нижний большой премоляр P4 приблизительно вдвое ниже C1, но заметно выше остальных нижних зубов. Коронка P4 трехгранная, с непрерывным узким цингулидом, слабо утолщенным у дистального края.
Нижние моляры никталодонтного типа, с узким непрерывным лабиальным цингулидом. M1 немного длиннее M2, протокристид на M1 короче и сильнее скошен назад, чем на M2. M3 немного короче M2. На M3 талонид сравнительно узкий, гипоконулид мельче, чем на M1 и M2.
Размеры в мм представлены в табл. 1, 2.
Таблица 1.
Размеры (в мм) черепа и верхних зубов Rhinolophus macrorhinus Topál, 1963 (n – число экземпляров)
| Параметры | R. macrorhinus cimmerius subsp. nov. | R. macrorhinus macrorhinus [5, 6] | R. macrorhinus anomalidens [6] | ||
|---|---|---|---|---|---|
| n | пределы | среднее | голотип | пределы | |
| Длина небного мостика | 22 | 2.45–3.1 | 2.68 | 2.8 | 2.45–2.95 |
| Высота носовой выпуклости | 19 | 3.9–4.3 | 4.1 | 4.7 | 4.4–4.45 |
| Межглазничная ширина | 10 | 2.4–2.9 | 2.79 | 3.2 | – |
| Ширина на уровне C1–C1 | 4 | 6.2–6.7 | 6.5 | 6.7 | 7.1–7.3 |
| Ширина на уровне M2–M2 | 10 | 7.9–8.4 | 8.13 | – | – |
| Ширина на уровне M3–M3 | 6 | 8.2–8.7 | 8.45 | 8.7 | 8.6–8.75 |
| Длина C1–M3 | 6 | 8.2–8.5 | 8.37 | 9.2 | 8.3–8.85 |
| Длина C1–P4 | 8 | 3.15–3.4 | 3.29 | 3.77 | 3.68–3.91 |
| Длина P2–M3 | 2 | 6.65–6.7 | 6.68 | – | 6.6–7.0 |
| Длина P4–M3 | 18 | 6.05–6.5 | 6.29 | 6.75 | 6.15–6.7 |
| Длина M1–M3 | 26 | 4.8–5.4 | 5.19 | 5.65 | 4.68–5.49 |
| Наименьшее расстояние между C1 и P4 | 7 | 0.1–0.15 | 0.11 | – | 0.14–0.23 |
| Длина C1 | 28 | 2.0–2.2 | 2.11 | 2.4 | 2.08–2.45 |
| Ширина C1 | 28 | 1.6–1.9 | 1.75 | 1.8 | 1.72–2.08 |
| Длина P2 | 3 | 0.45–0.55 | 0.48 | – | 0.25–0.5 |
| Ширина P2 | 3 | 0.4–0.5 | 0.47 | – | 0.41–0.59 |
| Длина P4 | 29 | 1.2–1.4 | 1.3 | – | 1.22–1.5 |
| Ширина P4 | 28 | 2.1–2.5 | 2.31 | – | 2.13–2.54 |
| Длина M1 | 62 | 1.9–2.2 | 2.02 | 2.24 | 1.9–2.23 |
| Ширина M1 | 60 | 2.35–2.65 | 2.49 | 3.25 | 2.72–3.32 |
| Длина M2 | 52 | 1.8–2.15 | 1.95 | 2.16 | 1.72–2.08 |
| Ширина M2 | 51 | 2.25–2.6 | 2.46 | 2.88 | 2.41–3.0 |
| Длина M3 | 38 | 1.35–1.6 | 1.45 | 1.44 | 1.18–1.4 |
| Ширина M3 | 38 | 2.0–2.25 | 2.11 | 2.12 | 2.03–2.31 |
Таблица 2.
Размеры (в мм) нижней челюсти и нижних зубов Rhinolophus macrorhinus Topál, 1963 (n – число экземпляров)
| Параметры | R. macrorhinus cimmerius subsp. nov. | R. macrorhinus anomalidens [6] | R. cf. macrorhinus [7, 8] | ||||
|---|---|---|---|---|---|---|---|
| n | пределы | среднее | пределы | n | пределы | среднее | |
| Длина C1–M3 | 4 | 8.7–9.2 | 8.85 | 9.05–9.6 | – | – | – |
| Длина C1–P4 | 7 | 3.0–3.45 | 3.24 | 3.36–3.6 | 2 | 3.36–3.38 | 3.37 |
| Длина P2–M3 | 7 | 7.6–8.1 | 7.84 | – | – | – | – |
| Длина P2–P4 | 10 | 2.0–2.35 | 2.15 | – | – | – | 2.36 |
| Длина P4–M3 | 28 | 6.8–7.3 | 7.03 | 7.1–7.6 | – | – | – |
| Длина M1–M3 | 41 | 5.7–6.15 | 5.9 | 5.71–6.2 | 2 | 6.04–6.2 | 6.12 |
| Длина C1 | 7 | 1.2–1.3 | 1.26 | 1.5 (голотип) | 2 | 1.3 | 1.3 |
| Ширина C1 | 7 | 1.4–1.6 | 1.5 | 1.72 (голотип) | – | – | – |
| Длина P2 | 10 | 0.85–1.0 | 0.91 | 0.86–1.0 | – | – | – |
| Ширина P2 | 9 | 0.95–1.3 | 1.13 | 1.04–1.27 | – | – | – |
| Длина P3 | 7 | 0.25–0.38 | 0.31 | – | – | – | – |
| Ширина P3 | 7 | 0.3–0.65 | 0.42 | – | – | – | – |
| Длина P4 | 43 | 1.15–1.4 | 1.26 | 1.18–1.45 | 6 | 1.36–1.43 | 1.39 |
| Ширина P4 | 43 | 1.15–1.45 | 1.29 | 1.13–1.4 | 6 | 1.21–1.49 | 1.34 |
| Длина M1 | 75 | 2.05–2.25 | 2.16 | 2.0–2.27 | 5 | 2.07–2.21 | 2.17 |
| Ширина талонида M1 | 71 | 1.3–1.6 | 1.45 | 1.31–1.54 | 5 | 1.38–1.58 | 1.5 |
| Длина M2 | 68 | 1.9–2.2 | 2.0 | 1.9–2.13 | 4 | 2.08–2.1 | 2.09 |
| Ширина талонида M2 | 66 | 1.3–1.6 | 1.46 | 1.27–1.54 | 4 | 1.41–1.64 | 1.52 |
| Длина M3 | 56 | 1.75–1.95 | 1.88 | 1.81–2.03 | 3 | 1.93–2.0 | 1.96 |
| Ширина тригонида M3 | 57 | 1.2–1.5 | 1.35 | – | 3 | 1.29–1.41 | 1.36 |
| Ширина талонида M3 | 56 | 1.05–1.3 | 1.18 | 1.04–1.27 | 2 | 1.14–1.15 | 1.15 |
| Постдентальная длина | 43 | 5.35–6.15 | 5.8 | – | – | – | – |
| Высота в венечном отростке | 46 | 3.9–4.4 | 4.14 | 4.05–4.36 | – | – | – |
Голотип ПИН, № 5644/940: длина небного мостика – 2.7; высота носового вздутия (от переднего края небного мостика до вершины) – 4.1; межглазничная ширина (в межглазничном сужении) – 2.6; ширина на уровне C1–C1 – 6.7; ширина на уровне M2–M2 – 8.1; ширина на уровне M3–M3 – 8.5; расстояние от середины сагиттального гребня до латерального выступа правой скуловой дуги – 6.4 (реконструированная ширина черепа в скуловых дугах – около 12.5–12.8); ширина черепа в мастоидных отростках – 11.6; длина C1–M3 – 8.45 (dex – правый, sin – левый); длина C1–P4 – 3.35 (dex, sin); длина P4–M3 – 6.3 (dex, sin); длина M1–M3 – 5.2 (dex, sin); наименьшее расстояние между C1 и P4 – 0.1 (dex), 0.15 (sin). Отношение длины небного мостика к длине C1–M3 – 0.32; отношение длины M1–M3 к длине C1–P4 – 1.55. Размеры зубов (L – длина, W – ширина, H – высота): C1 (dex): L – 2.1; W – 1.75; H – 3.5; C1 (sin): L – 2.1; W – 1.8; H – 3.5; P4 (dex, sin): L – 1.25, W – 2.35; M1 (dex): L – 2.0, W – 2.45; M1 (sin): L – 2.05, W – 2.45; M2 (dex): L – 1.95, W – 2.4; M2 (sin): L – 2.0, W – 2.4; M3 (dex): L – 1.5, W – 2.15; M3 (sin): L – 1.35, W – 2.15.
Экз. ПИН, № 5644/801: длина небного мостика – 2.7; высота носового вздутия – 3.9; межглазничная ширина – 2.9; ширина на уровне C1–C1 – 6.6; ширина на уровне M2–M2 – 8.15; ширина на уровне M3–M3 – 8.6; длина C1–M3 – 8.3 (dex, sin); длина C1–P4 – 3.15 (dex, sin); длина P2–M3 – 6.7 (dex); длина P4–M3 – 6.4 (dex, sin); длина M1–M3 – 5.3 (dex, sin). Отношение длины небного мостика к длине C1–M3 – 0.325; отношение длины M1–M3 к длине C1–P4 – 1.68.
Экз. ПИН, № 5644/936: длина небного мостика – 3.1; высота носового вздутия – 4.1; межглазничная ширина – 2.9; ширина на уровне C1–C1 – 6.2; ширина на уровне M2–M2 – 8.4; ширина на уровне M3–M3 – 8.7; длина C1–M3 – 8.5 (dex); длина C1–P4 – 3.3 (dex); длина P4–M3 – 6.45 (dex); длина M1–M3 – 5.3 (dex); наименьшее расстояние между C1 и P4 – 0.1 (dex). Отношение длины небного мостика к длине C1–M3 – 0.365; отношение длины M1–M3 к длине C1–P4 – 1.61.
Экз. ПИН, № 5644/939: длина небного мостика – 2.7; высота носового вздутия – 4.2; ширина на уровне M2–M2 – 8.0; длина P4–M3 – 6.35 (dex); длина M1–M3 – 5.3 (dex); длина латерального тяжа антеровентральной турбиналии – 3.1, высота горизонтальной порции латерального тяжа антеровентральной турбиналии – 0.5.
Экз. ПИН, № 5644/821: длина C1–M3 – 8.8; длина C1–P4 – 3.1; длина P2–M3 – 7.6; длина P2–P4 – 2.0; длина P4–M3 – 6.9; длина M1–M3 – 5.7; постдентальная длина – 6.0; высота в венечном отростке – 4.2.
Экз. ПИН, № 5644/822: длина C1–M3 – 8.7; длина C1–P4 – 3.2; длина P2–M3 – 7.7; длина P2–P4 – 2.15; длина P4–M3 – 6.8; длина M1–M3 – 5.8; постдентальная длина – 5.7; высота в венечном отростке – 4.1.
Изменчивость. Положение небного мостика относительно верхнего зубного ряда у описываемой формы изменчиво – его передний и задний края находятся соответственно на уровне паракона M1 и мезостиля M2 (экз. ПИН, №№ 5644/815, 938), паракона M1 и метакона M2 (экз. ПИН, № 5644/810), протокона M1 и метакона M2 (голотип; экз. ПИН, №№ 5644/801–803, 805, 939, 950), мезостиля M1 и метастиля M2 (экз. ПИН, №№ 5644/804, 952), протокона M1 и парастиля M3 (экз. ПИН, № 5644/936). У современных крупных подковоносов R. ferrumequinum (Schreber, 1774) и R. nippon Temminck, 1835 передний край небного мостика характеризуется более постоянным положением – немного дистальнее уровня вершины метакона M1 у R. ferrumequinum, на уровне или немного мезиальнее мезостиля M1 у большинства R. nippon [14].
P3 полностью редуцирован (нет альвеолы P3 между альвеолами P2 и P4) в двух случаях (экз. ПИН, №№ 5644/849, 851; рис. 1р), тогда как его присутствие наблюдается в 82 случаях (семь экземпляров с сохранившимся P3: экз. ПИН, №№ 5644/775, 776, 821, 822, 852, 853, 974; 75 экземпляров – с альвеолой P3), т.е. у 97.6% всех изученных экземпляров.
В материале имеется несколько лишенных корней и не вполне сформированных в основании нестертых коронок нижних моляров (M1, экз. ПИН, №№ 5644/916, 945; M2, экз. ПИН, №№ 5644/917, 918), которые принадлежали ювенильным особям с еще не прорезавшимися коренными зубами. Обусловленная стиранием онтогенетическая изменчивость зубной системы отражена увеличением площади фасеток стирания и уменьшением высоты зубных бугорков. У наиболее старых особей из-за сильного стирания жевательная поверхность моляров выровнена, высота главных бугорков клыков и больших премоляров заметно уменьшена, малые премоляры также несут следы стирания (рис. 4г).
На экз. ПИН, № 5644/822 M1 частично разрушен – отколота лингвальная стенка коронки на участке от передней части метаконида до середины энтоконида, в результате чего вскрыта полость зуба в области метаконида. Видимо, это прижизненное повреждение, так как поверхность и границы слома заглажены и слегка затерты (рис. 1л, 1м, 4в), что может свидетельствовать о довольно продолжительном функционировании данного зуба после травмы.
Сравнение и замечания. Крупные верхние зубы, сильно развитые полки гипокона M1–M2, относительно длинный и узкий M3 и степень редукции малых премоляров P2 и P3 позволяют относить раннеплейстоценового крымского подковоноса к виду R. macrorhinus – широко распространенному в Центральной и Юго-Восточной Европе в начале плейстоцена представителю крупных подковоносов из группы R. ferrumequinum [5–7, 15–17].
Новый подвид отличается от R. m. macrorhinus Topál, 1963 из виллафранка Венгрии (Беременд 4) более мелкими размерами, существенно меньшей высотой носовой выпуклости (табл. 1), наличием постеролингвального углубления в основании коронки C1, а также заметно большим расстоянием между полкой гипокона M2 и задним краем верхнечелюстной кости.
Отличия нового подвида от поздневиллафранкского R. m. anomalidens Topál, 1979 включают слабее выраженную носовую выпуклость, относительно мелкие верхние и нижние клыки (при сходной величине моляров) и менее редуцированные по длине M3 (табл. 1, 2). Отношение длины M1–M3 к длине C1–P4 у R. m. cimmerius subsp. nov. (1.47–1.68) существенно выше, чем у R. m. anomalidens (~1.3–1.4, см. [6]), как и отношение длины M1–M3 к длине C1–P4 (1.8–1.9 у крымской формы против ~1.7 у R. m. anomalidens).
Описание R. macrorhinus cimmerius subsp. nov. является первой регистрацией Rhinolophus в раннем плейстоцене Крыма. По ряду зубных признаков (величина клыков, отношение длины M1–M3 к длине C1–P4, степень редукции M3) новый подвид менее специализирован, чем R. m. anomalidens, что подразумевает его происхождение от более древней формы, морфологически близкой R. m. macrorhinus.
Современные подковоносы обитают в тропиках, субтропиках и на юге умеренного пояса Восточного полушария, в разнообразных ландшафтах от пустынь до горных лесов, охотятся на насекомых и других мелких членистоногих поблизости от убежищ. Большой подковонос R. ferrumequinum населяет пустынные предгорья и лиственные леса, часто селится в пещерах. Обилие остатков R. macrorhinus cimmerius subsp. nov. в нижнеплейстоценовых отложениях пещеры Таврида свидетельствует о теплом климате и наличии как открытых, так и лесных биотопов во времена формирования данного местонахождения.
Материал. Помимо голотипа, из пещеры Таврида (из разных точек и уровней) 227 экземпляров, в том числе фрагменты лицевой части черепов: с правыми C1, P2, P4–M3 и левыми C1, P4–M3 (экз. ПИН, № 5644/801), с правыми и левыми C1, P4–M3 (экз. ПИН, № 5644/936), с левыми C1, P4–M3 и правыми C1, P4–M2 (экз. ПИН, № 5644/950), с правыми и левыми P4–M3 (экз. ПИН, №№ 5644/802, 937, 938), с правыми P4–M3 и левыми M1–M3 (экз. ПИН, № 5644/939), с левыми C1, P4–M2 и правыми P4–M2 (экз. ПИН, № 5644/803), без зубов (экз. ПИН, № 5644/951); ростральные фрагменты черепов: с правыми и левыми P4–M2 (экз. ПИН, № 5644/952), с правым M1 и левыми M2–M3 (экз. ПИН, №№ 5644/804, 805), с правыми M1–M3 (экз. ПИН, № 5644/953), с правыми M1–M2 и фрагментом левого P4 (экз. ПИН, № 5644/954), с правым M1 (экз. ПИН, №№ 5644/806, 955), без зубов (экз. ПИН, №№ 5644/807, 808); фрагменты межглазничной части черепов (экз. ПИН, №№ 5644/809, 949); фрагменты верхнечелюстных костей: с обломанным C1 и целыми P2–M3 (экз. ПИН, № 5644/810), с C1 и P2–M2 (экз. ПИН, № 5644/956), с P4–M3 (экз. ПИН, №№ 5644/811–813, 941, 942, 957–959), с P4–M2 (экз. ПИН, №№ 5644/814–816), с P4–M1 (экз. ПИН, № 5644/960), с P4 (экз. ПИН, №№ 5644/961, 962), с обломанным P4 (экз. ПИН, № 5644/963), с M1–M3 и обломанным P4 (экз. ПИН, № 5644/771), с M1–M2 и обломанным P4 (экз. ПИН, № 5644/772), с M1 и обломанным P4 (экз. ПИН, № 5644/818), с M3 и обломанным P4 (экз. ПИН, № 5644/773), с M1–M3 (экз. ПИН, №№ 5644/817, 964), с M1–M2 (экз. ПИН, № 5644/965), с M1 (экз. ПИН, № 5644/966), с M2–M3 (экз. ПИН, № 5644/967–969), с M2 (экз. ПИН, №№ 5644/819, 970), с M3 (экз. ПИН, № 5644/880), без зубов (экз. ПИН, №№ 5644/820, 971–973, 983, 997); полные или почти полные нижнечелюстные кости: с C1, P2, P3 и P4–M3 (экз. ПИН, №№ 5644/821, 822, 974), с C1, P2 и P4–M3 (экз. ПИН, № 5644/823), с P2 и P4–M3 (экз. ПИН, №№ 5644/774, 824, 825), с P3 и P4–M3 (экз. ПИН, №№ 5644/775, 776), с P4–M3 (экз. ПИН, №№ 5644/777, 826–831, 833–839, 975–978), с M1–M3 (экз. ПИН, №№ 5644/778, 840–846, 979, 980), с M1 (экз. ПИН, № 5644/981), с M2–M3 (экз. ПИН, № 5644/779), без зубов (экз. ПИН, №№ 5644/780, 847–851, 913, 982); фрагменты нижнечелюстных костей: с C1, P2, P3 и P4–M2 (экз. ПИН, № 5644/852), с C1, P2, P3 и P4–M1 (экз. ПИН, № 5644/853), с C1, P2 и P4 (экз. ПИН, № 5644/984), с P4–M3 (экз. ПИН, № 5644/855), с P4–M2 (экз. ПИН, № 5644/985), с P4–M1 и обломанным M2 (экз. ПИН, № 5644/904), с P4–M1 (экз. ПИН, №№ 5644/781, 856–861, 905, 986, 987), с P4 (экз. ПИН, №№ 5644/854, 862), с M1–M3 и обломанным P4 (экз. ПИН, № 5644/988), с M1–M3 (экз. ПИН, №№ 5644/782, 863), с M1–M2 (экз. ПИН, №№ 5644/864–868, 989, 995), с M1 (экз. ПИН, №№ 5644/783, 869), с M1 и фрагментом M2 (экз. ПИН, № 5644/921), с M2–M3 (экз. ПИН, №№ 5644/870–875, 877, 878, 990–992), с M2 (экз. ПИН, №№ 5644/879), с M3 (экз. ПИН, №№ 5644/881, 993, 994), с основаниями M1–M3 (экз. ПИН, № 5644/882), с основаниями M2–M3 (экз. ПИН, № 5644/920), без зубов (экз. ПИН, №№ 5644/791–799, 883–888, 912, 996); изолированные зубы (экз. ПИН, №№ 5644/784–790, 800, 832, 847, 876, 889–903, 906–911, 914–919, 922–935, 943–948, 998).
Список литературы
Лопатин А.В., Вислобокова И.А., Лавров А.В., и др. Пещера Таврида – новое местонахождение раннеплейстоценовых позвоночных в Крыму // Доклады Академии наук. 2019. Т. 485. № 3. С. 381–385.
Лопатин А.В., Тесаков А.С. Раннеплейстоценовая белозубка Crocidura kornfeldi (Lipotyphla, Soricidae) из Крыма // Доклады Российской академии наук. Науки о жизни. 2021. Т. 501. № 1. С. 499–504.
Topál G., Csorba G. The subspecific division of Rhino-lophus luctus Temminck, 1835, and the taxonomic status of R. beddomei Andersen, 1905 (Mammalia, Chiroptera) // Miscellanea Zoologica Hungarica. 1992. V. 7. P. 101–116.
Gunnell G.F., Eiting T.P., Geraads D. New late Pliocene bats (Chiroptera) from Ahl al Oughlam, Morocco // Neues Jahrbuch für Geologie und Paläontologie. Abh. 2011. V. 260. № 1. P. 55–71.
Topál G. Description of a new bat, Rhinolophus macrorhinus sp. n. from the Lower Pleistocene of Hungary // Vertebrata Hungarica. 1963. V. 5. № 1–2. P. 219–226.
Topál G. Fossil bats of the Rhinolophus ferrumequinum group in Hungary (Mammalia: Chiroptera) // Fragmenta Mineralogica et Palaeontologica. 1979. V. 9. P. 61–101.
Wołoszyn B.W. Pliocene and Pleistocene bats of Poland // Acta Palaeontologica Polonica. 1987. V. 32. № 3–4. P. 207–325.
Wołoszyn B.W. Microevolutionary trends in Western Palaearctic bats. Case study: microevolutionary trends among bats of Rhinolophus “ferrumequinum” group (Mammalia: Chiroptera) // Travaux du Muséum National d’Histoire Naturelle “Grigore Antipa”. 2010. V. 53. P. 413–421.
Shimizu M., Miyao T. A study of bats. II. The palatal ridges of the bat // Journal of the Faculty of Liberal Arts, Shinshu University. 1967. Pt 2. № 1. P. 1–25.
Eisentraut M. Das Gaumenfaltenmuster der Säugetiere und seine Bedeutung für stammesgeschichtliche und taxonomische Untersuchungen // Bonner Zoologische Monographien. 1976. № 8. P. 1–214.
Curtis A.A., Simmons N.B. Unique turbinal morphology in horseshoe bats (Chiroptera: Rhinolophidae) // The Anatomical Record. 2017. V. 300. № 2. P. 309–325.
Curtis A.A., Smith T.D., Bhatnagar K.P., et al. Maxilloturbinal aids in nasophonation in horseshoe bats (Chiroptera: Rhinolophidae) // The Anatomical Record. 2020. V. 303. № 1. P. 110–128.
Ito K., Tu V.T., Eiting T.P., et al. On the embryonic development of the nasal turbinals and their homology in bats // Frontiers in Cell and Developmental Biology. 2021. V. 9. Art. 613545. P. 1–19.
Ikeda Y., Jiang T., Oh H., et al. Geographic variations of skull morphology in the Rhinolophus ferrumequinum species complex (Mammalia: Chiroptera) // Zoologischer Anzeiger. 2020. V. 288. P. 125–138.
Topál G. Bats from the Lowermost Pleistocene Locality 15 at Beremend, Hungary (Mammalia: Chiroptera) // Fragmenta Mineralogica et Palaeontologica. 1985. V. 12. P. 51–57.
Stormark J.G. Fossil bat fauna (Mammalia: Chiroptera) from Pliocene/Pleistocene localities in Southern Poland // Karst Waters Institute Special Publication. 1996. № 2. P. 43–44.
Rosina V.V., Agadjanian A.K., Shunkov M.V. Bats (Chiroptera) and other small mammals (Rodentia, Lagomorpha, Carnivora) from the Pleistocene site Trlica (Montenegro): biostratigraphical and paleoecological implications // Quaternaire. 2016. V. 27. № 4. P. 353–367.
Дополнительные материалы отсутствуют.
Инструменты
Доклады Российской академии наук. Науки о жизни