

Доклады Российской академии наук. Науки о жизни, 2022, T. 507, № 1, стр. 441-444
CHASMAPORTHETES LUNENSIS (HYAENIDAE, CARNIVORA) ИЗ РАННЕГО ПЛЕЙСТОЦЕНА КРЫМА
А. В. Лавров 1, Д. Р. Хантемиров 2, *, Д. О. Гимранов 3, академик РАН А. В. Лопатин 1
1 Палеонтологический институт им. А.А. Борисяка Российской академии наук
Москва, Россия
2 Уральский федеральный университет имени первого президента России Б.Н. Ельцина
Екатеринбург, Россия
3 Институт экологии растений и животных Уральского отделения Российской академии наук
Екатеринбург, Россия
* E-mail: hantemirov.d@mail.ru
Поступила в редакцию 28.06.2022
После доработки 25.08.2022
Принята к публикации 25.08.2022
- EDN: CZMWWY
- DOI: 10.31857/S2686738922060178
Аннотация
Из раннеплейстоценового местонахождения в пещере Таврида (Крым, поздний виллафранк, возраст 1.8–1.5 млн. лет) описан фрагмент верхнечелюстной кости вымершей гиены Chasmaporthetes lunensis (Del Campana, 1914), типичного представителя виллафранкских фаун Евразии. Это первая находка рода Chasmaporthetes в плейстоцене Крыма.
Род Chasmaporthetes Hay, 1921, объединяет неогеновые и плейстоценовые виды сравнительно крупных, но легко сложенных гиен, из-за длинных тонких конечностей получивших название “охотящихся гиен” (hunting hyenas). Предполагается, что в отличие от многих других гиен хасмапортетесы были активными охотниками, хорошо адаптированными к бегу.
Специализация к активному хищничеству также отражена в строении зубной системы этих гиен. Все щечные зубы представителей рода Chasmaporthetes режущего (гиперкарниворного) типа. Предкоренные зубы более тонкие, чем у других плио-плейстоценовых гиен, с режущими лезвиями главных бугорков и сильно развитыми задними добавочными базальными бугорками. Для Chasmaporthetes характерен узкий хищнический зуб m1 без метаконида; его короткий талонид не имеет бассейна (в отличие от других гиен) и состоит из одного режущего лезвия, которое сформировано гипоконидом [1–3].
Род Chasmaporthetes существовал с позднего миоцена (туролия) до раннего плейстоцена (1.4–0.8 млн. л.н.). Его ареал включал значительную часть Евразии (C. lunensis (Del Campana, 1914); C. borissiaki (Khomenko, 1932); C. bonisi Koufos, 1987; C. exitelus Kurtén et Werdelin, 1988; C. gangsriensis Tseng, Li et Wang, 2013), Африки (C. nitidula Ewer, 1955; C. australis Hendey, 1974) и Северной Америки (C. ossifragus Hay, 1921) [1, 4]. Это единственный род Hyaenidae, проникший в Северную Америку [4, 5].
Наибольшее распространение представители рода Chasmaporthetes получили в виллафранке. На территории Евразии с позднего плиоцена по ранний плейстоцен был широко распространен C. lunensis. В Испании этот вид был найден в плиоценовых местонахождениях Лайна и Вильяройя [6–8] и раннеплейстоценовом местонахождении Пуэбла-де-Вальверде [9]. На территории Франции C. lunensis известен из плиоценового местонахождения Этуэр и из нижнего плейстоцена местонахождений Сенез, Рокканейра, Сен-Валье и Пардин [10, 11]. В Италии раннеплейстоценовые остатки C. lunensis найдены в местонахождениях Оливола, Валь-ди-Магра и Инферно [10, 12], в Германии – в местонахождени Эрпфинген [13].
За пределами Европы C. lunensis был найден в Турции в плиоценовых местонахождениях Гулязи [14] и Чалта [15]. Материалы по C. lunensis из плиоцена и плейстоцена Евразии были детально описаны М.В. Сотниковой [2, 16] из местонахождений Береговая (Россия, Забайкалье), Одесские катакомбы (Украина, MN15; подвид C. l. odessanus Sotnikova, 1994), Шамар (Монголия, MN16), Куруксай (Таджикистан, MNQ17). С. lunensis также присутствует в нескольких китайских плиоценовых местонахождениях [1, 17]. Помимо C. lunensis в Азии выделялись также C. kani Galiano et Frailey, 1977, C. progressus Qiu, 1987 и C. bielawskyi (Schaub, 1941) [см. 17], но в настоящее время они обоснованно считаются синонимами С. lunensis [18].
Находки Chasmaporthetes в Восточной Европе редки. В связи с этим значительный интерес представляют новые материалы из пещеры Таврида в Крыму. Эта пещера расположена в 15 км восточнее Симферополя около п. Зуя Белогорского района. По составу фауны позвоночных костеносный слой датируется ранним плейстоценом (поздний виллафранк, псекупский комплекс, 1.8–1.5 млн. л.н.) [19]. Во время раскопок в пещере было найдено множество костей хищных млекопитающих, в том числе Hyaenidae. Подавляющее большинство остатков гиен (к настоящему времени их найдено более 100) принадлежит гигантской гиене Pachycrocuta brevirostris (Gervais, 1850), и лишь одна находка из сборов 2021 г. – описанный ниже верхнечелюстной фрагмент – относится к C. lunensis (рис. 1). Это первая находка Chasmaporthetes в плейстоцене Крыма см. [20].
Рис. 1.
Chasmaporthetes lunensis (Del Campana, 1914), экз. ПИН, № 5644/110, фрагмент левой верхнечелюстной кости с Р2–Р3: а – с окклюзиальной стороны, б – с лабиальной стороны, в – с лингвальной стороны; Крым, пещера Таврида; нижний плейстоцен. Обозначения: 1 – дистальный край альвеолы С1; 2 – дистальная часть альвеолы Р1; 3 – фасетка стирания от окклюзии с р2; 4 – фасетка стирания от окклюзии с р3; 5 – цингулюм.

Изученный образец хранится в Палеонтологическом институте им. А.А. Борисяка РАН (ПИН), Москва. Измерения проводились штангенциркулем с точностью до 0.1 мм. На зубах измерялись наибольшая переднезадняя длина (L) и наибольшая лабиолингвальная ширина (W).
На фрагменте левой верхнечелюстной кости (экз. ПИН, № 5644/110) сохранились целые двухкорневые премоляры P2–P3 и части альвеол однокорневых зубов – клыка C1 и переднего премоляра P1 (рис. 1). Дистальный край альвеолы С1 вплотную примыкает к альвеоле Р1. Ширина уцелевшей части альвеолы Р1 – 11.9 мм, реконструируемая ширина Р1 – 12.2–12.4 мм. Дистальный край альвеолы Р1 без диастемы примыкает к альвеоле Р2.
Премоляры P2–P3 имеют высокий конусовидный главный бугорок, редуцированный передний добавочный бугорок, смещенный лингвально, сильно развитый задний добавочный базальный бугорок, а также характерные для Chasmaporthetes мощный непрерывный цингулюм и базальный лингвальный выступ (выступ протокона), смещенный дистально.
Главные бугорки P2–P3 лабиолингвально сжатые, выпуклые с лабиальной стороны и почти плоские с лингвальной стороны, слегка наклонены назад. Главный бугорок на Р2 существенно короче и ниже, чем на P3. Передний и задний режущие гребни хорошо выражены. Основание переднего режущего гребня смещено лингвально от оси зуба. Передние добавочные бугорки сильно редуцированы и уплощены, смещены лингвально (различимы с лингвальной стороны P2 и P3 по наличию соответствующих складок в основании главных бугорков). Задние добавочные базальные бугорки высокие, сильно сжатые лабиолингвально, отделены от главных бугорков глубокими вырезками. Цингулюм особенно мощно выражен вдоль лингвальной стороны, а на лабиальной стороне – дистальнее главного бугорка. Выступ протокона не имеет собственной вершины, его поверхность ровная и наклонена лингвально. Край выступа протокона четко очерчен лингвальным цингулюмом.
Размеры зубов (в мм) экз. ПИН, № 5644/110: P2: L – 18.2; W – 12.0; P3: L – 23.2; W – 15.2.
Мезиальный край Р2 отчетливо смещен лингвально относительно осевой линии зубов Р1–Р3 и имеет фасетку стирания от окклюзии с р2. В дистальной части выступа протокона Р3 четко выражена фасетка стирания от окклюзии с р3. Это указывает на небольшую экскурсию (подвижность) нижней челюсти, необходимую для плотного примыкания премоляров при их окклюзии.
Экз. ПИН, № 5644/110 демонстрирует характерные признаки Chasmaporthetes, включая лабиолингвально сжатые главные бугорки с развитыми передними и задними режущими гребнями на верхних премолярах. Размеры и строение Р2–Р3 у экз. ПИН, № 5644/110 соответствуют таковым C. lunensis (см. рис. 2). Редукция передних добавочных бугорков характерна для C. lunensis в отличие от C. exitelus и С. nitidula [1, 2]. Строение премоляров соответствует режущей специализации пары хищнических зубов Р4/m1.
Рис. 2.
Сравнение размеров верхних премоляров Р2 и Р3 Chasmaporthetes и Pachycrocuta (L – длина, W – ширина, в мм): а – P2, б – P3. Обозначения: звезда – С. lunensis (Del Campana, 1914), экз. ПИН, № 5644/110, Таврида, Крым, Россия; квадрат – С. lunensis, различные местонахождения Евразии [6, 11]; косой крест – C. borissiaki (Khomenko, 1932), Молдавия и Франция [1]; круг – P. brevirostris (Gervais, 1850), Таврида, Крым, Россия: без заливки – экз. ПИН, № 5644/104; с заливкой – экз. ПИН, № 5644/162.
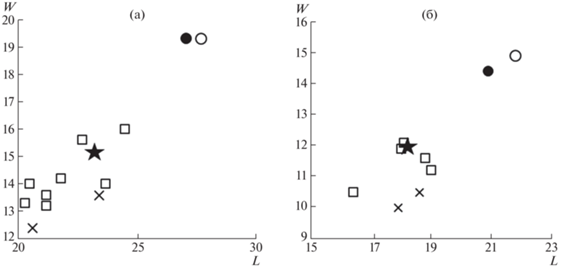
Соотношение числа находок остатков Chasmaporthetes lunensis и Pachycrocuta brevirostris в пещере Таврида близко к 1:100. Можно полагать, что хасмапортетес был относительно редким элементом фауны млекопитающих в раннем плейстоцене Восточной Европы. Присутствие C. lunensis соответствует возрасту 1.8–1.5 млн. л.н., установленному для фаунистического комплекса Тавриды на основании анализа его состава [19]. Данная находка Chasmaporthetes является одной из самых поздних в Европе.
Список литературы
Kurtén B., Werdelin L. A review of the genus Chasmaporthetes Hay, 1921 (Carnivora, Hyaenidae) // Journal of Vertebrate Paleontology. 1988. V. 8. № 1. P. 46–66.
Сотникова М.В. Хищные млекопитающие плиоцена – раннего плейстоцена: стратиграфическое значение. Москва: Наука, 1989. 124 с. (Труды ГИН АН СССР. Вып. 440).
artstone-Rose A. Reconstructing the diets of extinct South African carnivorans from premolar ‘intercuspid notch’ morphology // Journal of Zoology. 2011. V. 285. № 2. P. 119–127
Berta A. The Plio-Pleistocene hyaena Chasmaporthetes ossifragus from Florida // Journal of Vertebrate Paleontology. 1981. V. 1. № 3–4. P. 341–356.
Tseng Z.J., Li Q., Wang X. A new cursorial hyena from Tibet, and analysis of biostratigraphy, paleozoogeography, and dental morphology of Chasmaporthetes (Mammalia, Carnivora) // Journal of Vertebrate Paleontology. 2013. V. 33. № 6. P. 1457–1471.
Villalta Comella J.F. de. Contribución al conocimiento de la fauna de mamíferos fósiles del Plioceno de Villarroya (Logroño) // Boletín del Instituto Geológico y Minero de España. 1952. № 64. P. 1–203.
Crusafont M., Aguirre E. Euryboas lunensis et Hyaena donnezani associées, en Espagne dans le gisement d’age Pliocène terminal de Layna (Soria) // Comptes Rendus de l’Académie des sciences, Paris. Sèr. D. 1971. V. 273. P. 2476–2478.
Kurtén B., Crusafont Pairó M. Villafranchian carnivores (Mammalia) from La Puebla de Valverde (Teruel, Spain). Helsinki: Societas Scientiarum Fennica, 1977. 39 p.
Antón M., Turner A., Salesa M.J., Morales J. A complete skull of Chasmaporthetes lunensis (Carnivora, Hyaenidae) from the Spanish Pliocene site of La Puebla de Valverde (Teruel) // Estudios Geológicos. 2006. V. 62. № 1. P. 375–388.
Schaub S. Ein neues Hyaenidengenus von der Montagne de Perrier // Eclogae Geologicae Helvetiae. 1941. V. 34. № 2. P. 279–286.
Viret J. Le loess à bancs durcis de Saint-Vallier (Drôme), et sa faune de mammifères villafranchiens // Nouvelles Archives de Museum d’Histoire Naturelle de Lyon. 1954. Fasc. 4. P. 1–200.
Ficcarelli G., Torre D. Una mandibola di Euryboas lunensis (Del Campana) nel giacimento Villafranchiano di Olivola (Val di Magra) // Memorie della Societa Toscana di Scienze Naturali. Ser. A. 1967. V. 74. P. 193–198.
Lehmann U. Weitere Fossilfunde aus dem ältesten Pleistozän der Erpfinger Höhle (Schwäbische Alb) // Mitteilungen aus dem Geologischen Staatsinstitut in Hamburg. 1957. V. 26. P. 60–99.
Schütt G. Hyaenidenfunde aus dem Villafranchium der Türkei // Mitteilungen aus dem Geologischen Institut der Universität Hannover. 1971. V. 10. P. 119–140.
Sen S., Saraç G. Hyaenidae (Carnivora, Mammalia) from late Miocene and Pliocene of Çalta (Ankara, Turkey) // Revue de Paléobiologie. 2018. V. 37. № 2. C. 561–575.
Сотникова М.В. Род Chasmaporthetes Нау, 1921 из плиоцена России, Украины, Монголии и Таджикистана // Палеотериология / Ред. Л.П. Татаринов. Москва: Наука, 1994. С. 113–139.
Qiu Z., Deng T., Wang B. Early Pleistocene mammalian fauna from Longdan, Dongxiang, Gansu, China // Palaeontologia Sinica. New ser. C. 2004. № 191. P. 1–198.
Werdelin L., Solounias N. The Hyaenidae: taxonomy, systematics and evolution // Fossils and Strata. 1991. № 30. P. 1–104.
Лопатин А.В., Вислобокова И.А., Лавров А.В., и др. Пещера Таврида – новое местонахождение раннеплейстоценовых позвоночных в Крыму // Доклады Академии наук. 2019. Т. 485. № 3. С. 381–385.
Родионов В.В., Титов В.В. Раннеплиоценовая фауна млекопитающих с юго-западного побережья Крыма // 100-летие Палеонтологического общества России. Проблемы и перспективы палеонтологических исследований. Материалы LXII сессии Палеонтологического общества при РАН (4–8 апреля 2016 г., Санкт-Петербург). Санкт-Петербург: ВСЕГЕИ, 2016. С. 269.
Дополнительные материалы отсутствуют.
Инструменты
Доклады Российской академии наук. Науки о жизни