

Доклады Российской академии наук. Науки о жизни, 2022, T. 507, № 1, стр. 530-535
ПАРАФИЛИЯ НЕТОПЫРЕЙ (PIPISTRELLUS; VESPERTILIONIDAE) ПОДТВЕРЖДАЕТСЯ АНАЛИЗОМ ЯДЕРНЫХ ГЕННЫХ МАРКЕРОВ
С. С. Жукова 1, Е. Н. Соловьева 2, И. В. Артюшин 1, С. В. Крускоп 2, 3, *
1 Биологический факультет Московского государственного университета имени М.В. Ломоносова
Москва, Россия
2 Зоологический музей Московского государственного университета имени М.В. Ломоносова
Москва, Россия
3 Совместный Российско-Вьетнамский Тропический научно-исследовательский и технологический центр
Ханой, Вьетнам
* E-mail: selysius@mail.ru
Поступила в редакцию 15.08.2022
После доработки 01.09.2022
Принята к публикации 08.09.2022
- EDN: ECGUWI
- DOI: 10.31857/S2686738922060361
Аннотация
Существуют свидетельства того, что и в ныне принятом составе род настоящих нетопырей, Pipistrellus, может быть парафилетичным. Нами впервые изучены филогенетические отношения между большинством надвидовых таксонов подтрибы Pipistrellina на основании анализа последовательностей трех ядерных генов (RAG2, APOB и DMP1). Наши данные свидетельствуют в пользу монофилии самой подтрибы, однако определенно показывают, что род Pipistrellus в современном понимании парафилетичен относительно вечерниц, Nyctalus, и толстопалых нетопырей, Glischropus (оба таксона хорошо обособлены морфологически). Более того, вечерницы оказались крайне близки к “западной” кладе Pipistrellus. Вероятно, Pipistrellus должен быть разделен на два или более самостоятельных рода, либо Nyctalus и Glischropus должны быть включены в его состав в качестве подродов.
ВВЕДЕНИЕ
На протяжении десятилетий род нетопыри, Pipistrellus, был своего рода таксономической “мусорной корзиной”, объединяя мелких гладконосых летучих мышей с одной парой малых предкоренных зубов в верхней челюсти [1, 2]. Однако в результате морфологических, кариологических и молекулярно-генетических исследований [3–5] многие виды были исключены из его состава и перенесены в самостоятельные рода, зачастую неродственные собственно Pipistrellus (такие как Hypsugo, Falsistrellus, Neoromicia, Arielulus, Perimyotis и др. [6]). В принятом ныне объеме, род Pipistrellus несомненно объединяет формы, связанные сравнительно близким родством [4, 7], однако вопрос его монофилии остается открытым. Группа видов “rueppeli”, очевидно делавшая Pipistrellus парафилетичным, была выделена в самостоятельный род Vansonia [8]. Однако монофилия остальных Pipistrellus всерьез не обсуждалась, хотя отдельные опубликованные данные указывали на то, что настоящие нетопыри, вероятно, парафилетичны относительно таких признанных родов, как Nyctalus и Glischropus [7–9].
Объем подтрибы Pipistrellina здесь принят в понимании [10] и соответствует трибе Pipistrellini согласно [6, 11]. Чтобы проверить гипотезу о парафилии Pipistrellus, нами были получены последовательности трех ядерных генов для ряда видов этого рода, а также родственных родов, и на их основе осуществлен филогенетический анализ.
МАТЕРИАЛЫ И МЕТОДЫ
Геномную ДНК выделяли из фиксированных в этаноле проб тканей стандартным методом фенол-хлороформной депротеинизации [12]. Были получены последовательности трех ядерных генов (RAG2, 1234 bp; DMP1, 886 bp; и APOB, 889 bp). Выбор этих ядерных маркеров определен их вариабельностью и представленностью в ГенБанке, что позволило расширить анализируемую выборку. Последовательности праймеров взяты из опубликованных работ [13, 14]. Реакция амплификации осуществлялась при следующих условиях: первичная денатурация – 94°C в течение 3 мин; затем 35 циклов, включающих денатурацию – 94°C в течение 45 с, отжиг c функцией touch down – 68–65° в течение 1 мин, синтез – 72°C в течение 1 мин. Конечный синтез осуществлялся при 72°C в течение 7 мин. Опыты выполнены на приборе My Cycler BioRAD. Контроль амплификации проводили в 1% агарозном геле. Очистку продуктов ПЦР проводили с помощью набора Diatom DNA Clean-Up. Автоматическое секвенирование проведено на секвенаторе ABI PRISM 3500xl. Всего получено 119 последовательностей ядерных генов для 11 видов; еще 66 последовательностей 10 видов были получены из ГенБанка. Виды родов Eptesicus, Vespertilio и Hypsugo взяты в качестве внешней группы. Номера использованных музейных экземпляров (обозначены литерой S) и номера ГенБанка указаны на рис. 1–2.
Рис. 1.
Дерево филогенетических отношений подтрибы Pipistrellina по расширенному набору последовательностей ядерных генов APOB, DMP1, RAG2, построенное методом Байеса (апостериорные вероятности получены для цепи в 5 000 000 генераций), поддержки над или под узлами обозначают апостериорные вероятности/значения бутстрепа, полученные анализом ML. Цветом выделены четыре основные обсуждаемые клады; группы видов нетопырей “западной” клады: “nath” – nathusii, “pip” – pipistrellus, “kuhl” – kuhlii.
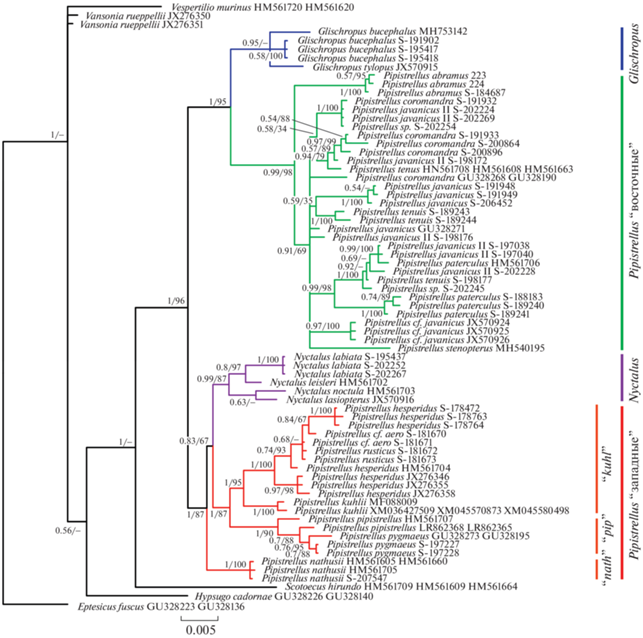
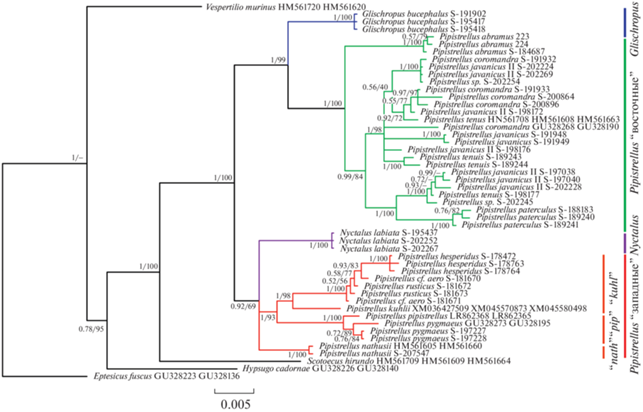
Рис. 2.
Дерево филогенетических отношений подтрибы Pipistrellina по сокращенному набору последовательностей ядерных генов APOB, DMP1, RAG2, построенное методом Байеса (апостериорные вероятности получены для цепи в 5 000 000 генераций), цифры над узлами обозначают апостериорные вероятности/значения бутстрепа, полученные анализом ML. Обозначения таксонов как в рис. 1.
Последовательности выровнены с помощью программ BioEdit v7.1.3. и SeqMan Pro v.7.1. [15, 16] и скорректированы вручную. Анализ конкатенированных последовательностей выполнен в двух вариантах: “расширенном”, с максимальным таксономическим покрытием, и “сокращенном”, куда включены только экземпляры, для которых получены последовательности хотя бы двух генов из трех.
Реконструкция филогенетических деревьев выполнена методами максимального правдоподобия (ML) при помощи программы IQ-Tree и методом байесова анализа (BA) с помощью программы MrBayes 3.1.2. Для определения достоверности полученной топологии ML использовали процедуру “bootstrap” с 1000 репликами. Некорректированные меж- и внутригрупповые p-дистанции посчитаны в программе MEGA.
Таблица 1.
Некорректированные p-дистанции (%) по расширенному набору конкатенатов генов яДНК: межгрупповые дистанции под диагональю, внутригрупповые – на диагонали, над диагональю – стандартная ошибка. Дистанции для P. abramus рассчитаны отдельно от остальных “восточных” Pipistrellus
| P. abramus | Glischropus | “восточные” Pipistrellus | группа видов “kuhlii” | P. nathusii | Nyctalus | группа видов “pipistrellus” | |
|---|---|---|---|---|---|---|---|
| P. abramus | 0.08 | 0.39 | 0.22 | 0.35 | 0.40 | 0.36 | 0.38 |
| Glischropus | 2.53 | 0 | 0.33 | 0.37 | 0.57 | 0.38 | 0.43 |
| “восточные” Pipistrellus | 1.99 | 2.40 | 1.17 | 0.31 | 0.36 | 0.30 | 0.34 |
| группа видов “kuhlii” | 3.70 | 2.80 | 3.64 | 0.63 | 0.29 | 0.26 | 0.26 |
| P. nathusii | 3.69 | 2.71 | 3.49 | 1.96 | 0 | 0.28 | 0.41 |
| Nyctalus | 3.55 | 2.76 | 3.26 | 2.17 | 1.63 | 0 | 0.28 |
| группа видов “pipistrellus” | 3.71 | 3.11 | 3.51 | 2.15 | 2.71 | 2.17 | 0.48 |
Таблица 2.
Некорректированные p-дистанции (%) по сокращенному набору конкатенатов генов яДНК: межгрупповые дистанции под диагональю, внутригрупповые – на диагонали, над диагональю – стандартная ошибка
| “восточные” Pipistrellus | Glischropus | Nyctalus | “западные” Pipistrellus | |
|---|---|---|---|---|
| “восточные” Pipistrellus | 1.33 | 0.33 | 0.31 | 0.29 |
| Glischropus | 2.41 | 0.00 | 0.39 | 0.37 |
| Nyctalus | 3.29 | 2.76 | 0.00 | 0.23 |
| “западные” Pipistrellus | 3.59 | 2.88 | 2.13 | 1.44 |
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Наши данные как по каждому отдельному гену, так и по конкатенированным последовательностям в целом согласуются с ранее опубликованными (напр., [7, 9]). Топологии деревьев, построенных методами Байеса и ML, не имели существенных различий. Монофилия подтрибы Pipistrellina относительно Eptesicus и Vespertilio поддержана на дереве, построенном по конкатенату методом Байеса (рис. 1, 2). Положение рода Hypsugo (представленного видом H. cadornae) спорно, в ряде реконструкций он занимает сестринское положение к Pipistrellina либо корневое положение внутри нее. Роды Vansonia (V. rueppeli) и Scotoecus (S. hirundo), судя по полученным реконструкциям, принадлежат к кладе Pipistrellina, но их положение относительно друг друга неоднозначно.
Монофилия клады Pipistrellina в узком смысле (Pipistrellus+Nyctalus+Glischropus) имеет высокие поддержки на всех полученных реконструкциях. Она разделяется на две клады следующего порядка, одна из которых включает “западные” виды нетопырей (группы видов “pipistrellus”, “kuhlii” и “nathusii”) и род Nyctalus, а вторая – “восточные” виды нетопырей (группы видов “javanicus” и “stenopterus”) и род Glischropus.
В “западной” кладе все три группы видов нетопырей монофилетичны и имеют максимальные поддержки. Положение Nyctalus неоднозначно: на некоторых деревьях, построенных по отдельным генам, вечерницы объединяются с группой видов “nathusii”; при “сокращенном” наборе таксонов формируется неразрешимая трихотомия Nyctalus – группа “nathusii” – группы “pipistrellus” и “kuhlii”.
В “восточной” кладе на всех реконструкциях род Glischropus занимает базальное положение. Все включенные в анализ “восточные” нетопыри формируют хорошо поддержанную монофилетическую группу. При этом базальное положение в ней занимает P. abramus, а не морфологически обособленный P. stenopterus.
Анализ каждого из ядерных генов дает в среднем более низкие поддержки большинства узлов, однако в целом топология этих деревьев не противоречит вышеописанной. Все три гена поддерживают монофилию подтрибы Pipistrellina, при этом во всех случаях род Scotoecus занимает в ней базальное положение. Разделение нетопырей на “западных” и “восточных” также поддержано каждым из генов, однако по гену RAG2 монофилия “западной” клады имеет низкую поддержку, а данные по гену DMP1 исключают Glischropus из “восточной” клады, делая равноценным членом неразрешенной трихотомии. Род Nyctalus на всех реконструкциях объединяется с “западной” кладой нетопырей.
Стоит отметить, что значение генетических p-дистанций между “восточными” и “западными” нетопырями выше, чем между этими кладами (и входящими в них группами видов) и родами Glishropus и Nyctalus (табл. 1, 2). Аналогичную ситуацию демонстрируют опубликованные ранее дистанции по митохондриальным генам, хоть и посчитанные для меньшей таксономической выборки [9].
Таким образом, по нашим данным, род Pipistrellus в современном понимании безусловно парафилетичен относительно родов Nyctalus и Glischropus. Более того, генетическая линия морфологически обособленного рода Nyctalus с высокой вероятностью располагается внутри “западной” клады Pipistrellus, будучи обособленной от групп видов “pipistrellus” и “kuhlii” не в большей степени, чем группа “nathusii”. Все это говорит о необходимости пересмотра границ рода Pipisterllus и в целом таксономических границ в подтрибе Pipistrellina.
ЗАКЛЮЧЕНИЕ
Наши данные полностью подтверждают предположение о парафилии рода Pipistrellus в современном его понимании. Из ныне признанных надвидовых таксонов подтрибы Pipistrellina в нашем анализе отсутствует только род Scotozous (который еще не был изучен генетически). Однако его добавление в филогенетическую реконструкцию будет ставить вопрос только о его собственном статусе и не должно повлиять на относительное положение Pipistrellus, Nyctalus и Glischropus. Таким образом, “восточная” клада нетопырей должна быть выделена в самостоятельный таксон группы рода (род или подрод). При этом абсолютное большинство названий таксонов группы рода в пределах Pipistrellus s. lato было предложено для представителей “западной” клады (напр., [17]); единственное исключение – название Alionoctula, предложенное для морфологически обособленной группы видов “stenopterus” [9]. Таким образом, выделение “восточной” клады в самостоятельный род должно быть сопряжено с введением нового названия. Однако описание такого таксона требует уточнения его границ (за счет увеличения числа проанализированных видов) и составления его морфологического диагноза. Такая работа, определенно, должна быть сделана в ближайшем будущем.
Трактовка уровня таксономических различий между полученными кладами неоднозначна. Вечерниц традиционно рассматривают как хорошо обособленный самостоятельный род, однако его положение на филогенетическом дереве предполагает, что не только группа видов “javanicus”, но, возможно, и группа видов “nathusii” должны рассматриваться как самостоятельные роды. Альтернативным вариантом может быть рассмотрение всех групп видов нетопырей, а также Nyctalus и Glischropus как подродов в составе Pipistrellus, что противоречит предполагаемому времени дивергенции этих таксонов (средний миоцен [18, 19]). Аналогичные ситуации можно наблюдать и в других таксонах рукокрылых, например, в Molossinae (см. [20]). Таким образом, наше исследование, помимо решения конкретной таксономической проблемы, приобретает и методологическое значение.
Список литературы
Hill J.E., Harrison D.L. The baculum in the Vespertilioninae (Chiroptera: Vespertilionidae) with a systematic review, a synopsis of Pipistrellus and Eptesicus, and the descriptions of a new genus and subgenus // Bulletin of the British Museum (Natural History), 1987. V. 52. P. 225–305.
Koopman K.F. Chiroptera: Systematics, VIII. Mammalia, part 60. New York: Walter de Gruyter, 1994. 217 p.
Horaček I., Hanak V. Generic status of Pipistrellus savii and comments on classification of the genus Pipistrellus (Chiroptera, Vespertilionidae) // Myotis, 1986. V. 23–24. P. 9–16.
Volleth M., Bronner G., Göpfert M.C., et al. Karyotype comparison and phylogenetic relationships of Pipistrellus-like bats (Vespertilionidae; Chiroptera; Mammalia) // Chromosome Research, 2001. V. 9. № 1. P. 25–46.
Kearney T.C., Volleth M., Contrafatto G., Taylor P.J. Systematic implications of chromosome GTG-band and bacula morphology for southern African Eptesicus and Pipistrellus and several other species of Vespertilioninae (Chiroptera: Vespertilionidae) // Acta Chiropterologica, 2002. V. 4. № 1. P. 55–76.
Moratelli R., Burgin C., Cláudio V., et al. Family Vespertilionidae (Vesper Bats) / Eds Wilson D.E., Mittermeier R.A. Handbook of the Mammals of the World, Volume 9: Bats. Barcelona: Lynx Edicions, 2019. 1008 p.
Roehrs Z.P., Lack J.B., Van Den Bussche R.A. Tribal phylogenetic relationships within Vespertilioninae (Chiroptera: Vespertilionidae) based on mitochondrial and nuclear sequence data // Journal of Mammalogy, 2010. V. 91. № 5. P. 1073–1092.
Koubínová D., Irwin N., Hulva P., et al. Hidden diversity in Senegalese bats and associated findings in the systematics of the family Vespertilionidae // Frontiers in Zoology, 2013. V. 10. № 48. P. 1–16.
Kruskop S.V., Solovyeva E.N., Kaznadzey A.D. Unusual Pipistrelle: taxonomic position of the Malayan Noctule (Pipistrellus stenopterus; Vespertilionidae; Chiroptera) // Zoological Studies, 2018. V. 57. № 60. P. 1–15.
Крускоп С.В. Отряд Chiroptera / Ред Павлинов И.Я., Лисовский А.А. Млекопитающие России: систематико-географический справочник. Москва: Т-во научных изданий КМК, 2012. 604 с.
Hoofer S.R., Van den Bussche R.A. Molecular phylogenetics of the chiropteran family Vespertilionidae // Acta Chiropterologica, 2003. V. 5. Suppl. P. 1–63.
Sambrook J., Fritsch E.F., Maniatis T. Molecular cloning – a laboratory manual. Sec. ed. N.Y.: Cold Spring Harbour Lab. Press, 1989. 385 p.
Baker R.J., Porter C.A., Patton J.C., Van Den Bussche T.A. Systematics of bats of the family Phyllostomidae based on Rag2 DNA sequences // Occasional Papers of Museum of the Texas Technical University, 2000. V. 202. P. 1–16.
Artyushin I.V., Kruskop S.V., Lebedev V.S., Bannikova A.A. Molecular phylogeny of Serotines (Mammalia, Chiroptera, Eptesicus): evolutionary and taxonomical aspects of the E. serotinus species group // Biology Bulletin, 2018. V. 45. № 5. P. 469–477.
Hall T.A. BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT // Nucleotide, 1999. V. 41. P. 95–98.
Burland T.G. DNASTAR’s lasergene sequence analysis software // Methods in Molecular Biology, 1999. V. 132. P. 71–91.
Ellerman J.R., Morrison-Scott T.C.S. Checklist of Palaearctic and Indian Mammals 1758 to 1946. Second ed. London: British Museum (Natural History), 1966. 810 p.
Lack J.B., Van Den Bussche R.A. Identifying the confounding factors in resolving phylogenetic relationships in Vespertilionidae // Journal of Mammalogy, 2010. V. 91. № 6. P. 1435–1448.
Rosina V.V., Sinitsa M.V. Bats (Chiroptera, Mammalia) from the Turolian of Ukraine: phylogenetic and biostratigraphic considerations // Neues Jahrbuch für Geologie und Paläontologie, 2014. V. 272. № 2. P. 147–166.
Ammerman L.K., Lee D.N., Tipps T.M. First molecular phylogenetic insights into the evolution of free-tailed bats in the subfamily Molossinae (Molossidae, Chiroptera) // Journal of Mammalogy, 2012. V. 93. № 1. P. 12–28.
Дополнительные материалы отсутствуют.
Инструменты
Доклады Российской академии наук. Науки о жизни