

Доклады Российской академии наук. Науки о жизни, 2023, T. 508, № 1, стр. 95-104
РАННЕПЛЕЙСТОЦЕНОВЫЙ КОЖАН Eptesicus praeglacialis (Vespertilionidae, Chiroptera) ИЗ ПЕЩЕРЫ ТАВРИДА В КРЫМУ
Академик РАН А. В. Лопатин 1, *
1 Палеонтологический институт им. А.А. Борисяка Российской академии наук
Москва, Россия
* E-mail: alopat@paleo.ru
Поступила в редакцию 05.07.2022
После доработки 09.08.2022
Принята к публикации 10.08.2022
- EDN: TKTYSB
- DOI: 10.31857/S268673892206018X
Аннотация
Из нижнеплейстоценовых отложений пещеры Таврида в центральном Крыму описаны черепные и нижнечелюстные остатки крупного кожана Eptesicus praeglacialis Kormos, 1930. Это первая находка черепов E. praeglacialis и первое свидетельство обитания данного вида в Крыму. Судя по степени стирания зубов, в тафоценозе представлены остатки как молодых, так и взрослых особей. Следы зубов мелких млекопитающих на костях (связанные с поеданием остатков мягких тканей) при отсутствии признаков дигестии, характерной для материалов из погадок хищных птиц, указывают, что в тафоценоз попали остатки погибших в пещере особей E. praeglacialis, которые использовали ее в качестве укрытия. Это соответствует представлениям о появлении на рубеже плиоцена и плейстоцена у некоторых лесных форм европейских летучих мышей (включая Eptesicus) климатически обусловленной экологической адаптации – гибернации в пещерах.
Изучение остатков раннеплейстоценовых рукокрылых из пещеры Таврида в центральном Крыму (Белогорский район, пос. Зуя) позволило описать новый подвид подковоносов Rhinolophus macrorhinus cimmerius Lopatin, 2022 (Rhinolophidae) и установить присутствие гладконосых летучих мышей родов Eptesicus, Plecotus и Myotis (Vespertilionidae) [1].
В настоящей статье описываются черепные и нижнечелюстные остатки крупных кожанов Eptesicus praeglacialis Kormos, 1930 из нижнеплейстоценовых отложений пещеры Таврида (сборы 2018, 2020 и 2021 гг.). Возраст фаунистического комплекса определен в пределах позднего виллафранка, около 1.8–1.5 млн л.н. [2, 3].
Изученный материал хранится в Палеонтологическом институте им. А.А. Борисяка РАН (ПИН) в г. Москве. Терминология строения зубов по [4]. Размеры приведены в мм. Иллюстрации подготовлены при помощи цифрового фотоаппарата Nikon D800 с объективом AF-S Micro NIKKOR 60mm f/2.8G ED, сканирующего электронного микроскопа Tescan Vega 3 XMU и рентгеновского микротомографа Neoscan N80 в ПИН.
Отряд Chiroptera Blumenbach, 1779
Семейство Vespertilionidae Gray, 1821
Подсемейство Vespertilioninae Gray, 1821
Триба Eptesicini Volleth et Heller, 1994
Род Eptesicus Rafinesque, 1820
Eptesicus praeglacialis Kormos, 1930
Eptesicus n. sp.: [5: с. 500].
Eptesicus praeglacialis n. sp.: [6: с. 237].
Голотип – Венгерский геологический институт, Будапешт, № 06/4728 [7: с. 56]; левая нижнечелюстная кость с P4–M3 и восходящей ветвью; Румыния, Бихор, местонахождение Бетфия 2, или Сомлёберг (Somlyóberg), близ Пюспёкфюрдо (Püspökfürdő); нижний плейстоцен.
Описание (рис. 1–4). Размеры крупные, заметно больше, чем у современного E. serotinus (Schreber, 1774). Череп массивный, низкий, мозговая коробка удлиненная, широкая (существенно шире лицевого отдела), куполовидно выпуклая в задней части, с легкими вздутиями позади межглазничного промежутка. Заглазничное сужение хорошо выражено. Ширина черепа в скуловых дугах намного превосходит ширину в мастоидных отростках.
Рис. 1.
Eptesicus praeglacialis Kormos, 1930: а–е – экз. ПИН, № 5644/741, череп с правыми и левыми P4 и M2–M3: а – с дорсальной стороны, б – с вентральной стороны, в – с правой латеральной стороны, г – с левой латеральной стороны, д – с ростральной стороны, е – с затылочной стороны; ж–к – экз. ПИН, № 5644/742, череп с правыми C1–M2 и левыми P4–M3: ж – с дорсальной стороны, з – с вентральной стороны, и – с правой латеральной стороны, к – с затылочной стороны; л–п – экз. ПИН, № 5644/743, череп с левыми P4–M3 и правыми M2–M3: л – с дорсальной стороны, м – с вентральной стороны, н – с правой латеральной стороны, о – с ростральной стороны, п – с затылочной стороны; р–у – экз. ПИН, № 5644/744, неполный череп с правыми и левыми C1–M3: р – с дорсальной стороны, с – с вентральной стороны, т – с правой латеральной стороны, у – с ростральной стороны; Россия, Крым, пещера Таврида; нижний плейстоцен.
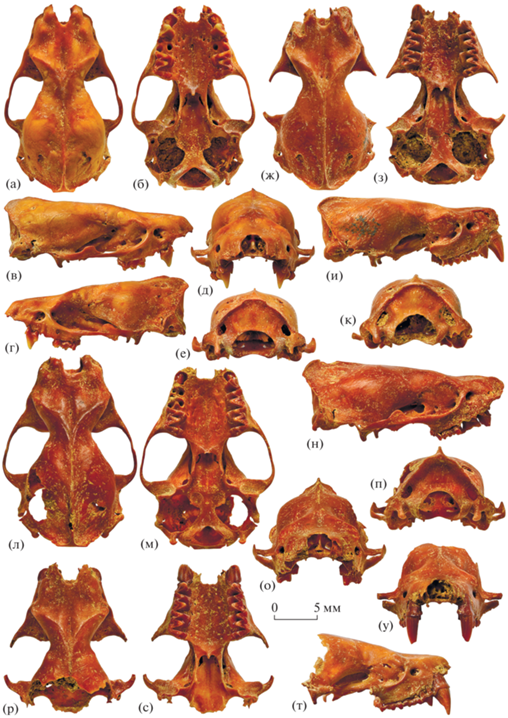
Рис. 2.
Eptesicus praeglacialis Kormos, 1930: а–г – экз. ПИН, № 5644/745, неполный череп с правыми и левыми C1–M3: а – с дорсальной стороны, б – с вентральной стороны, в – с левой латеральной стороны, г – с ростральной стороны; д–л – экз. ПИН, № 5644/746, неполный череп с правыми C1–M1 и левыми C1–P4: д – с дорсальной стороны, е – с вентральной стороны, ж – с правой латеральной стороны, з – с ростральной стороны; и–л: компьютерная томография: и, к – левая половина носовой полости: и – сагиттальная проекция, вблизи носовой перегородки (видны этмотурбиналии I, II и III); к – сагиттальная проекция, более латерально (видны сечения этмотурбиналий и максиллотурбиналии); л – носовая полость, аксиальная проекция на уровне M1; Россия, Крым, пещера Таврида; нижний плейстоцен.

Рис. 3.
Eptesicus praeglacialis Kormos, 1930: а – экз. ПИН, № 5644/747, фрагмент левой верхнечелюстной кости с P4–M3 с окклюзиальной стороны; б–г – экз. ПИН, № 5644/751, левая нижнечелюстная кость с I1–I2 и C1–M3: б – с лабиальной стороны, в – с окклюзиальной стороны, г – с лингвальной стороны; д–з – экз. ПИН, № 5644/750, правая нижнечелюстная кость с I2–M3: д – с лабиальной стороны, е – с окклюзиальной стороны, ж – с окклюзиальной стороны с лингвальным наклоном, з – с лингвальной стороны; и–л – экз. ПИН, № 5644/752, правая нижнечелюстная кость с P4–M3: и – с лабиальной стороны, к – с окклюзиальной стороны, л – с лингвальной стороны; м–о – экз. ПИН, № 5644/753, левая нижнечелюстная кость с M3: м – с лабиальной стороны, н – с окклюзиальной стороны, о – с лингвальной стороны; Россия, Крым, пещера Таврида; нижний плейстоцен.
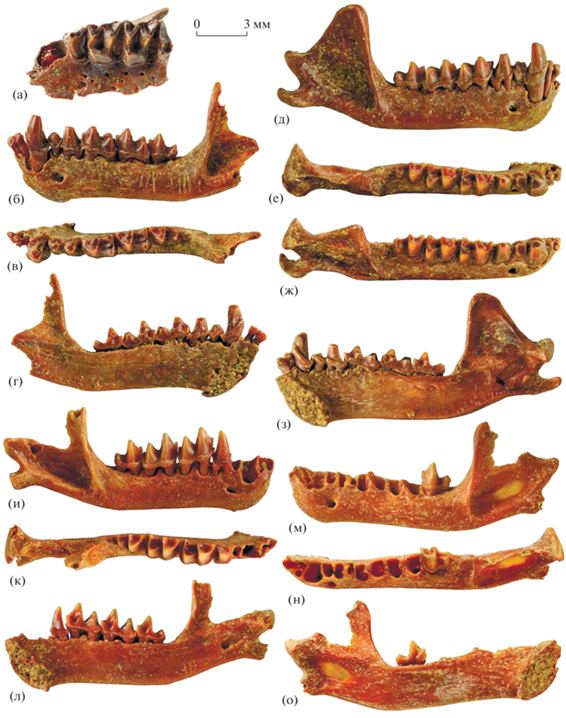
Рис. 4.
Eptesicus praeglacialis Kormos, 1930, зубные ряды с окклюзиальной стороны: а – экз. ПИН, № 5644/747, слабо стертые левые P4–M3 и альвеола C1; б – экз. ПИН, № 5644/752, нестертые правые P4–M3 и альвеолы I1–P2; в – экз. ПИН, № 5644/750, умеренно стертые правые I2–M3 и альвеола I1; г – экз. ПИН, № 5644/751, умеренно стертые левые I1–I2 и C1–M3 и альвеола I3; Россия, Крым, пещера Таврида; нижний плейстоцен.

Лицевой отдел черепа крупный, ростральная часть широкая, с дорсальной стороны слегка вогнутая, с четкой продольно вытянутой узкой срединной впадиной. Клыки широко расставленные. Вырезка носового отверстия сравнительно неглубокая, V-образной или U-образной формы. Лобные кости расширены над глазницами и в месте наибольшего латерального расширения формируют умеренно развитые предглазничные отростки. Надглазничные гребни хорошо выраженные. Сагиттальный гребень низкий почти на всем протяжении, но высокий в наиболее задней части, которая заметно выступает дистально, нависая над затылком (рис. 1в, 1г, 1и, 1н). Затылочные гребни высокие, резкие, их соединенные с окончанием сагиттального гребня поперечные порции короткие, прямые (рис. 1а, 1ж, 1л). Теменные отверстия крупные, раздвоенные.
Подглазничное отверстие крупное, округлое, расположено на уровне задней части P4 и переднего края M1. Костный мостик, отделяющий подглазничное отверстие от глазницы, короткий, но массивный. Округлое слезное отверстие находится внутри глазницы, на уровне середины или задней части M1. Скуловая дуга относительно прямая, низкая и тонкая, в ее средней части четко выделяется высокий и крупный заглазничный отросток трапециевидной, округло-треугольной или полукруглой формы (рис. 1в, 1г, 1н).
Задний край U-образной передней небной вырезки находится на уровне середины C1. Костное небо длинное, оканчивается немного дистальнее уровня заглазничного сужения. Срединный шип костного неба хорошо выраженный, но сравнительно короткий. Мезоптеригоидная ямка широкая, крыловидные отростки хорошо развитые, с немного отклоненными медиально задними концами (рис. 1б, 1с). Суставные впадины сравнительно небольшие. Боковые затылочные отростки короткие, не выступают дистальнее или вентральнее затылочных мыщелков.
Носовые раковины (турбиналии) характерного для Vespertilionidae строения [8: рис. 7D, 7H ]; на компьютерных томографических срезах (рис. 2и–2л) хорошо различимы крупные этмотурбиналии I, II и III, вместе занимающие значительную часть объема задней половины носовой полости, а также продольно вытянутая максиллотурбиналия вблизи дна носовой полости.
Верхний зубной ряд включает два резца, клык, один премоляр и три моляра (рис. 1з, 1с, 2б, 3а, 4а); верхний малый премоляр отсутствует. Ряд C1–P4 по длине в 1.5 раза уступает ряду M1–M3.
Судя по альвеолам, верхние резцы I1 и I2 тесно сближены и располагаются примерно на одной поперечной линии. Альвеола I1 немного крупнее альвеолы I2. Альвеола C1 отделена от альвеолы I2 небольшим промежутком; поверхность кости на месте этого промежутка вогнутая, с множеством мелких ямок.
Верхний клык С1 массивный, высокий, слегка загнут постеролингвально. Лабиальная сторона полукруглая, лингвальная сторона отчетливо вогнута в задней части. От вершины отходят четыре резких субвертикальных гребня – передний, антеролингвальный, толстый лабиальный и особенно мощный задний гребень, который соединяется с хорошо развитым задним выступом коронки. Цингулюм мощный, волнообразно изогнутый.
Верхний большой премоляр P4 вдвое ниже C1, короткий и относительно узкий, с субтреугольной окклюзиальной поверхностью, с крупным полукруглым талоном и узким непрерывным цингулюмом. Парастиль мелкий, гребень постпаракристы длинный, высокий. Имеется отчетливая задняя вырезка.
Верхние моляры M1 и M2 массивные, с трапециевидной окклюзиальной поверхностью. Стили крупные, выступ гипокона хорошо развит. Цингулюм узкий. M2 расширен поперечно сильнее, чем M1, парастиль на M2 вытянут существенно больше (поэтому W-образная фигура из гребней паракона и метакона выражена лучше). На M1 выступ гипокона выдается лингвально сильнее, чем протокон, тогда как на M2, напротив, сильнее выступает протокон. Задняя вырезка очень слабая.
M3 сильно редуцированный, узко-треугольный, вытянутый поперечно, с крупным парастилярным выступом, конец которого охватывает метастиль M2. На M3 метастиль отсутствует, метакон очень мелкий. Цингулюм на передней и лингвальной сторонах тонкий, но отчетливый; на постеролабиальной стороне (на участке от пара-стиля до метакона) цингулюм очень слабый, на уровне метакона отсутствует или едва намечен.
Горизонтальная ветвь нижнечелюстной кости относительно короткая и высокая, со слегка вогнутым в средней части вентральным краем и мощно развитым высоким подбородочным выступом (рис. 3б, 3д, 3и, 3м). Подбородочное отверстие относительно мелкое, округлое, расположено под P2 или промежутком P2/P4. Симфиз крупный, достигает уровня P2 или передней части P4. Нижнечелюстное отверстие мелкое, щелевидное, располагается немного вентральнее уровня альвеолярного края кости.
Восходящая ветвь нижнечелюстной кости высокая и длинная. Венечный отросток треугольный, с субвертикальным передним краем, относительно узкой вершиной и пологим задним склоном. Массетерная впадина глубокая, обрамляющие ее гребни высокие, мощные. Впадина височной мышцы глубокая и обширная, вентрально ограничена хорошо развитым медиальным гребнем, тянущимся от основания венечного отростка до суставного мыщелка. Сочленовный отросток короткий, суставный мыщелок широкий. Длина постдентальной части нижнечелюстной кости (расстояние от M3 до заднего края суставного мыщелка) на 10–15% превышает длину M1–M3. Угловой отросток широкий и массивный в основании, с узким концом, длинный, крючковидно загнутый дорсомедиально, заметно выступает дистально за задний край суставного мыщелка. На медиальной стороне основания углового отростка имеется отчетливая бороздка, которая протягивается от уровня нижнечелюстного отверстия до уровня вырезки между сочленовным и угловым отростками. На латеральной стороне углового отростка располагаются два мощных продольных гребня, дорсальный и вентральный. Они начинаются на уровне шейки мыщелка и сливаются под его головкой, а далее единый гребень тянется до окончания углового отростка, ограничивая (вместе с дорсальным гребнем) довольно обширную вогнутую площадку на его дорсолатеральной поверхности.
Нижний зубной ряд состоит из трех резцов, клыка, двух премоляров и трех моляров (рис. 3б–3з, 4б–4г). Длина ряда C1–P4 приблизительно в 1.7–1.8 раза меньше длины ряда M1–M3.
Нижние резцы мелкие, заметно наклонены вперед. Альвеолы I1 и I2 тесно сближены, коронка I2 частично перекрывает коронку I1. Альвеола I3 сильно смещена лабиально, но при этом корень I3 наклонен лингвально, в результате чего I3 налегает на I2 и коронки всех трех резцов формируют единую дугу.
Коронки нижних резцов трехлопастные, лабиально выпуклые, приблизительно одинаковых размеров. Лабиальная доля крупнее двух других. На задней стороне каждого резца имеется мелкий дополнительный бугорок, расположенный на уровне границы между лабиальной и срединной долями. На антеролабиальной стороне находится отчетливый цингулид, на I2 и I3 в его пределах развит дополнительный цингулярный бугорок. I3 тесно прилегает к C1.
Нижний клык C1 массивный, высокий, субвертикальный. Основание коронки округло-треугольное, передний и боковые края выпуклые, задний край почти прямой. Имеется слабо выраженный антеролингвальный дополнительный бугорок. Задняя стенка главного бугорка немного вогнутая, с мощным постеролингвальным ребром, на котором при стирании формируется широкий гребень. В постеролингвальном углу короткого заднего выступа коронки располагается мелкий дополнительный бугорок. Цингулид мощно развит на всех сторонах, но отсутствует в месте смыкания C1 с коронкой I3.
Нижний малый премоляр P2 однокорневой, однобугорковый, приблизительно в 2.2 раза ниже C1. Коронка широкая, с округло-треугольной окклюзиальной поверхностью (лингвальная сторона существенно длиннее лабиальной). Цингулид широкий, непрерывный, на его антеролингвальном и заднем выступах имеются маленькие цингулярные бугорки.
Нижний большой премоляр P4 приблизительно в 1.4 раза уступает по высоте C1. Основание коронки P4 округленно-квадратное, с непрерывным мощным цингулидом и антеролингвальным дополнительным бугорком. Вершина главного бугорка остроконечная (плоская при стирании), антеролабиальная сторона выпуклая, лингвальная сторона прямая, ограничена резкими передним и постеролингвальным ребрами, задняя сторона немного вогнутая. Задний выступ коронки короткий, с выступающим постеролингвально дополнительным бугорком.
Нижние моляры массивные, миотодонтного типа, с мощным эктоцингулидом и в разной степени развитым энтоцингулидом под выходом из широко открытого тригонидного бассейна. Талонидный бассейн закрыт высоким энтокристидом. M1 и M2 приблизительно равной длины, с выступающим постеролингвально крупным гипоконулидом; паракристид на M1 короче, чем на M2, косой кристид присоединен к задней стенке тригонида более лабиально. M3 относительно короткий, с узким двухбугорковым талонидом (иногда имеется крохотный гипоконулид, см. рис. 3ж, 4в); косой кристид присоединен к средней части задней стенки тригонида.
Размеры в мм. Экз. ПИН, № 5644/741: наибольшая длина черепа – 21.85; кондилобазальная длина – 21.6; кондилоканинная длина – 20.3; основная длина (от передней небной вырезки до большого затылочного отверстия) – 17.0; длина рострума (до заднего края небного отростка верхнечелюстной кости) – 9.5; длина костного неба (от передней небной вырезки до основания срединного шипа) – 8.5; длина суставной впадины (dex – правая, sin – левая) – 1.7 (dex, sin); ширина суставной впадины – 2.8 (dex, sin); межглазничная ширина (в заглазничном сужении) – 4.8; ширина в скуловых дугах – 14.2; ширина мозговой коробки – 10.5; ширина в мастоидных отростках – 11.4; ширина на уровне M2–M2 – 9.0; ширина мезоптеригоидной ямки (между основаниями крыловидных отростков) – 2.9; ширина большого затылочного отверстия – 4.5; высота мозговой коробки (от вентрального края затылочного мыщелка до вершины сагиттального гребня) – 7.5; длина P4–M3 – 6.5 (dex), 6.4 (sin). Размеры зубов (L – длина, W – ширина, H – высота): P4 (dex): L – 1.8; W – 1.9; P4 (sin): L – 1.7; W – 1.9; M2 (dex): L – 2.3; W – 2.7; M2 (sin): L – 2.2; W – 2.7; M3 (dex): L – 1.0; W – 2.6; M3 (sin): L – 1.0; W – 2.5.
Экз. ПИН, № 5644/742: наибольшая длина черепа – более 21.0 (рострум обломан); кондилоканинная длина – 19.8; основная длина – 17.0; длина костного неба – 8.3; длина суставной впадины – 1.6 (sin); ширина суставной впадины – 3.0 (sin); межглазничная ширина – 4.95; ширина мозговой коробки – 11.6; ширина в мастоидных отростках – 12.7; ширина на уровне M2–M2 – 9.2; ширина мезоптеригоидной ямки – 2.9; ширина большого затылочного отверстия – 4.5; высота мозговой коробки – 7.8; длина C1–P4 – 3.55 (dex); длина P4–M3 – 6.7 (sin); длина M1–M3 – 5.5 (sin). Отношение длины M1–M3 к длине C1–P4 – 1.55. Размеры зубов: C1 (dex): L – 2.2; W – 1.6; H – 3.4; P4 (dex): L – 1.5; W – 2.0; P4 (sin): L – 1.5; W – 2.1; M1 (dex): L – 2.3; W – 2.6; M1 (sin): L – 2.4; W – 2.5; M2 (dex): L – 2.2; W – 2.8; M2 (sin): L – 2.4; W – 2.8; M3 (sin): L – 1.2; W – 2.5.
Экз. ПИН, № 5644/743: наибольшая длина черепа – 22.5; кондилоканинная длина – 20.8; основная длина – 17.5; длина рострума – 9.5; длина костного неба – 9.0; длина суставной впадины – 1.7 (dex, sin); ширина суставной впадины – 3.2 (dex, sin); межглазничная ширина – 4.85; ширина в скуловых дугах – 15.6; ширина мозговой коробки – 11.4; ширина в мастоидных отростках – 12.3; ширина на уровне M2–M2 – 9.6; ширина мезоптеригоидной ямки – 2.9; ширина большого затылочного отверстия – 4.5; высота мозговой коробки – 8.1; длина P4–M3 – 6.8 (sin); длина M1–M3 – 5.3 (sin). Размеры зубов: P4 (sin): L – 1.65; W – 2.2; M1 (sin): L – 2.35; W – 2.6; M2 (dex): L – 2.3; W – 2.75; M2 (sin): L – 2.25; W – 2.85; M3 (dex): L – 1.05; W – 2.5; M3 (sin): L – 1.1; W – 2.6.
Экз. ПИН, № 5644/744: длина рострума – 9.0; длина костного неба – 8.7; межглазничная ширина – 4.7; ширина на уровне C1–C1 – 7.6; ширина на уровне M2–M2 – 9.4; длина C1–M3 – 8.75 (dex), 8.65 (sin); длина C1–P4 – 3.6 (dex, sin); длина P4–M3 – 6.7 (dex), 6.65 (sin); длина M1–M3 – 5.4 (dex, sin). Отношение длины M1–M3 к длине C1–P4 – 1.5 (dex, sin). Размеры зубов: C1 (dex): L – 2.25; W – 1.9; H – 3.5; C1 (sin): L – 2.25; W – 2.0; H – 3.3; P4 (dex): L – 1.6; W – 2.2; P4 (sin): L – 1.6; W – 2.15; M1 (dex): L – 2.25; W – 2.55; M1 (sin): L – 2.25; W – 2.5; M2 (dex, sin): L – 2.2; W – 2.8; M3 (dex): L – 1.1; W – 2.55; M3 (sin): L – 1.05; W – 2.55.
Экз. ПИН, № 5644/745: длина рострума – 9.0; длина костного неба – 8.3; межглазничная ширина – 4.9; ширина на уровне C1–C1 – 7.0; ширина на уровне M2–M2 – 8.6; длина C1–M3 – 8.2 (dex), 8.3 (sin); длина C1–P4 – 3.3 (dex, sin); длина P4–M3 – 6.4 (dex), 6.5 (sin); длина M1–M3 – 5.0 (dex), 5.05 (sin). Отношение длины M1–M3 к длине C1–P4 – 1.52 (dex), 1.53 (sin). Размеры зубов: C1 (dex): L – 2.25; W – 1.7; H – 3.4; C1 (sin): L – 2.15; W – 1.7; H – 3.4; P4 (dex): L – 1.5; W – 2.0; P4 (sin): L – 1.6; W – 2.0; M1 (dex, sin): L – 2.25; W – 2.45; M2 (dex): L – 2.25; W – 2.75; M2 (sin): L – 2.25; W – 2.7; M3 (dex, sin): L – 0.9; W – 2.5.
Экз. ПИН, № 5644/746: длина костного неба – 8.4; ширина на уровне C1–C1 – 7.65; длина C1–P4 – 3.85 (dex), 3.8 (sin). Размеры зубов: C1 (dex): L – 2.35; W – 1.85; H – 3.15; C1 (sin): L – 2.25; W – 1.85; H – 3.2; P4 (dex): L – 1.85; W – 2.2; P4 (sin): L – 1.85; W – 2.15; M1 (dex): L – 2.45; W – 2.7.
Экз. ПИН, № 5644/757, C1 (dex): L – 2.4; W – 1.9; H – 3.55.
Экз. ПИН, № 5644/758, C1 (dex): L – 2.2; W – 1.9; H – 3.7.
Экз. ПИН, № 5644/747 (sin): длина P4–M3 – 6.6; длина M1–M3 – 5.2. Размеры зубов: P4: L – 1.5; W – 2.1; M1: L – 2.25; W – 2.6; M2: L – 2.2; W – 2.85; M3: L – 1.05; W – 2.45.
Экз. ПИН, № 5644/748 (sin): P4: L – 1.75; W – 2.25; M1: L – 2.35; W – 2.7.
Экз. ПИН, № 5644/749 (sin): P4: L – 1.65; W – 2.2.
Экз. ПИН, № 5644/750 (dex): длина нижнечелюстной кости – 16.7; длина I2–I3 – 0.95; C1–M3 – 9.4; C1–P4 – 3.4; P2–M3 – 8.1; P2–P4 – 2.1; P4–M3 – 7.3; M1–M3 – 6.2; постдентальная длина – 6.6; расстояние от переднего края вершины венечного отростка до заднего края суставного мыщелка (плечо силы височной мышцы) – 5.7; расстояние от переднего края головки суставного мыщелка до вентрального края углового отростка (плечо силы массетера) – 3.2; высота горизонтальной ветви под P2 – 2.9, под M3 – 2.6; высота в венечном отростке – 6.4; ширина суставного мыщелка – 2.65. Отношение длины M1–M3 к длине C1–P4 – 1.82. Размеры зубов: I2: лабиолингвальная длина (LL) – 0.5; W – 0.8; I3: LL – 0.5; W – 0.9; C1: L – 1.5; W – 1.75; P2: L – 1.0; W – 1.15; P4: L – 1.3; W – 1.35; M1: L – 2.2; W – 1.6; M2: W – 1.6; M3: L – 1.9; ширина тригонида (WTR) – 1.3; ширина талонида (WTAL) – 0.9.
Экз. ПИН, № 5644/751 (sin): длина I1–M3 – 10.4; I1–C1 – 2.3; I1–I2 – 1.0; C1–M3 – 9.25; C1–P4 – 3.45; P2–M3 – 7.9; P2–P4 – 2.1; P4–M3 – 7.0; M1–M3 – 5.9; высота горизонтальной ветви под P2 – 2.8, под M3 – 2.5. Отношение длины M1–M3 к длине C1–P4 – 1.71. Размеры зубов: I1: LL – 0.5; W – 0.8; I2: LL – 0.55; W – 0.75; C1: L – 1.5; W – 1.7; P2: L – 0.95; W – 1.1; P4: L – 1.25; W – 1.25; M1: L – 2.1; W – 1.55; M2: L – 2.05; W – 1.6; M3: L – 1.7; WTR – 1.35; WTAL – 0.9.
Экз. ПИН, № 5644/752 (dex): длина нижнечелюстной кости – 16.2; длина P4–M3 – 7.05; M1–M3 – 6.1; постдентальная длина – 6.7; высота горизонтальной ветви под P2 – 2.6, под M3 – 2.4; ширина суставного мыщелка – 2.4. Размеры зубов: P4: L – 1.15; W – 1.3; M1: L – 2.15; W – 1.7; M2: L – 2.15; W – 1.65; M3: L – 1.7; WTR – 1.35; WTAL – 0.9.
Экз. ПИН, № 5644/753 (sin): длина нижнечелюстной кости – 16.35; альвеолярная длина: I1–M3 – 9.8; C1–M3 – 8.7; P4–M3 – 6.85; M1–M3 – 5.6; постдентальная длина – 6.5; расстояние от переднего края вершины венечного отростка до заднего края суставного мыщелка – 5.6; высота горизонтальной ветви под P2 – 2.5, под M3 – 2.3; высота в венечном отростке – 6.1; ширина суставного мыщелка – 2.4. Размеры M3: L – 1.8; WTR – 1.3; WTAL – 0.75.
Экз. ПИН, № 5644/754 (dex): альвеолярная длина: I1–M3 – 9.6; C1–M3 – 8.7; высота горизонтальной ветви под P2 – 2.8, под M3 – 2.6.
Экз. ПИН, № 5644/755 (dex): высота горизонтальной ветви под M3 – 2.6. Размеры зубов: M2: L – 2.15; W – 1.7; M3: L – 1.75; WTR – 1.35; WTAL – 0.85.
Экз. ПИН, № 5644/759, P4 (dex): L – 1.3; W – 1.35.
Экз. ПИН, № 5644/760, M1 (dex): L – 2.15; W – 1.55.
Статистические данные представлены в табл. 1.
Таблица 1.
Размеры (в мм) Eptesicus praeglacialis Kormos, 1930 из нижнего плейстоцена пещеры Таврида, Крым (n – число экземпляров)
| Параметры | n | пределы | среднее |
|---|---|---|---|
| Длина костного неба | 6 | 8.3–9.0 | 8.53 |
| Межглазничная ширина | 5 | 4.7–4.95 | 4.84 |
| Ширина на уровне C1–C1 | 3 | 7.0–7.65 | 7.42 |
| Ширина на уровне M2–M2 | 5 | 8.6–9.6 | 9.16 |
| Длина C1–M3 | 4 | 8.2–8.75 | 8.48 |
| Длина C1–P4 | 7 | 3.3–3.85 | 3.57 |
| Длина P4–M3 | 9 | 6.4–6.8 | 6.58 |
| Длина M1–M3 | 7 | 5.0–5.5 | 5.26 |
| Длина C1 | 9 | 2.15–2.4 | 2.26 |
| Ширина C1 | 9 | 1.6–2.0 | 1.82 |
| Длина P4 | 14 | 1.5–1.85 | 1.65 |
| Ширина P4 | 14 | 1.9–2.25 | 2.1 |
| Длина M1 | 10 | 2.25–2.45 | 2.31 |
| Ширина M1 | 10 | 2.45–2.7 | 2.57 |
| Длина M2 | 11 | 2.2–2.4 | 2.25 |
| Ширина M2 | 11 | 2.7–2.85 | 2.77 |
| Длина M3 | 10 | 0.9–1.2 | 1.04 |
| Ширина M3 | 10 | 2.45–2.6 | 2.53 |
| Длина P4–M3 | 3 | 7.0–7.3 | 7.12 |
| Длина M1–M3 | 3 | 5.9–6.2 | 6.07 |
| Длина P4 | 4 | 1.15–1.3 | 1.25 |
| Ширина P4 | 4 | 1.25–1.35 | 1.31 |
| Длина M1 | 4 | 2.1–2.2 | 2.15 |
| Ширина талонида M1 | 4 | 1.55–1.7 | 1.6 |
| Длина M2 | 3 | 2.05–2.15 | 2.12 |
| Ширина талонида M2 | 4 | 1.6–1.7 | 1.64 |
| Длина M3 | 5 | 1.7–1.9 | 1.77 |
| Ширина тригонида M3 | 5 | 1.3–1.35 | 1.33 |
| Ширина талонида M3 | 5 | 0.75–0.9 | 0.86 |
| Постдентальная длина | 3 | 6.5–6.7 | 6.6 |
Изменчивость. Возрастная изменчивость зубов, обусловленная их стиранием, выражается в увеличении площади фасеток стирания и уменьшении высоты бугорков. По степени стирания верхних клыков для представителей рода Eptesicus выделяются шесть категорий, или классов, стирания [9, 10]: (1) клыки с острой вершиной; (2) клыки с округлой вершиной; (3–6) клыки с плоской вершиной, стертые: (3) на 1/8–1/7 высоты коронки, (4) на 1/6–1/5 высоты коронки, (5) на 1/4–1/3 высоты коронки, (6) на 1/2 высоты коронки и более. По этой классификации экз. ПИН, №№ 5644/742, 744 и 745 относятся к категории 3, а экз. ПИН, № 5644/746 – к категории 4 (рис. 1у, 2г, 2з). Изолированные верхние клыки могут быть отнесены к категориям 2 (экз. ПИН, № 5644/758) и 3 (экз. ПИН, № 5644/757). Так как стирание клыков зависит от локальных и сезонных особенностей питания, прямое сопоставление классов стирания с возрастом в годах затруднительно [10], однако очевидно, что первые две категории относятся к молодым особям первых лет жизни, а последующие – к взрослым и старым особям [9].
Фрагмент нижнечелюстной кости экз. ПИН, № 5644/752 с нестертыми P4–M3 (рис. 3и–3л, 4б) явно принадлежит молодой особи первого года жизни.
Сравнение и замечания. Вид E. praeglacialis был кратко описан [6] из нижнего плейстоцена Румынии (Бетфия 2) по фрагменту нижнечелюстной кости с P4–M3 (без изображения). Сведения о другом типовом местонахождении [11: с. 155] и ином месте хранения голотипа [12: с. 89] ошибочны [7, 13].
Позже E. praeglacialis был идентифицирован (по большей части, без иллюстраций и морфологической характеристики) в нижнем плейстоцене Венгрии [14], Австрии [15], Мальты [16–18]. Eptesicus cf. praeglacialis был описан по изолированным верхним и нижним зубам из нижнего и среднего плейстоцена Венгрии [7].
Отнесение крымского раннеплейстоценового кожана к E. praeglacialis основывается на соответствующей величине (размеры голотипа в мм: длина нижнечелюстной кости – 16.7, альвеолярная длина C1–M3 – 8.85, длина P4–M3 – 7.05, длина M1–M3 – 6.1) [6, 19] и идентичных признаках нижнечелюстной кости (подбородочное отверстие под нижним малым премоляром; глубокая массетерная впадина; очень глубокая впадина височной мышцы с мощным медиальным гребнем; относительно узкая вершина венечного отростка; широкий угловой отросток с бороздкой на медиальной стороне и двумя резкими гребнями на латеральной стороне) и нижних коренных зубов (массивные, с мощным цингулидом), диагностических для данного вида [6]. Указанные признаки и крупные размеры (особенно крупные зубные ряды) отличают E. praeglacialis от наиболее близкого по величине и морфологии рецентного вида E. serotinus (Schreber, 1774), в ископаемом состоянии известного со среднего плейстоцена [20], а также от вымерших видов из плиоцена Европы (E. kowalskii Wołoszyn, 1987 и E. mossoczyi Wołoszyn, 1987) [11, 19].
Современный крупный кожан E. serotinus, сближаемый с E. praeglacialis, широко распространен как в открытых ландшафтах, так и в лесных местообитаниях и эпизодически использует пещеры для гибернации. Вместе с тем остатки E. serotinus обычны в пещерных тафоценозах погадочного происхождения, так как летучие мыши этого вида регулярно становятся добычей сов [19]. По числу остатков E. praeglacialis на порядок уступает наиболее многочисленным видам раннеплейстоценовых рукокрылых из пещеры Тав-рида – Rhinolophus macrorhinus Topál, 1963 и Myotis spp., но при этом представлен значительным числом образцов хорошей сохранности (почти целых черепов и нижнечелюстных костей), без каких-либо признаков дигестии, свойственной погадочным материалам. Некоторые экземпляры E. praeglacialis из пещеры Таврида несут следы зубов мелких млекопитающих. Довольно длинные, субпараллельные, плотно расположенные (частично перекрывающиеся) узкие поверхностные бороздки отмечаются на теменных костях неполного черепа экз. ПИН, № 5644/744 (рис. 1р). Короткие широкие бороздки располагаются на левой стороне дорсальной поверхности ростральной части черепа экз. ПИН, № 5644/745 (рис. 2а). На некоторых нижнечелюстных костях тонкие бороздки разной длины, субперпендикулярные вентральному краю кости, фиксируются на лабиальной и лингвальной сторонах горизонтальной ветви вблизи восходящей ветви (экз. ПИН, №№ 5644/750–752; рис. 3б, 3д, 3з, 3л), а также за симфизной областью (экз. ПИН, № 5644/752; рис. 3л); сходные бороздки отмечены на венечном отростке нижнечелюстной кости экз. ПИН, № 5644/750 (рис. 3д). Судя по глубине и локализации, все эти следы зубов представляют собой свидетельства избирательного поедания сохранившихся на костях остатков мягких тканей и не связаны с остеофагией. Так как в результате дигестии костный материал из погадок хищных птиц обычно полностью очищен от мягких тканей, наличие таких следов питания может указывать, что в тафоценоз попали остатки погибших в пещере особей E. praeglacialis, которые использовали ее в качестве укрытия. Это соответствует основанным на анализе тафономии европейских местонахождений представлениям о появлении на рубеже плиоцена и плейстоцена у некоторых лесных форм летучих мышей (Pipistrellus, Vesperti-lio, Nyctalus, Eptesicus) климатически обусловленной экологической адаптации – гибернации в пещерах [11].
Распространение. Нижний плейстоцен Европы.
Материал. Из пещеры Таврида 20 экземпляров, в том числе три почти целых черепа (без слуховых барабанов): с правыми и левыми P4 и M2–M3 (экз. ПИН, № 5644/741), с правыми C1–M2 и левыми P4–M3 (экз. ПИН, № 5644/742), с левыми P4–M3 и правыми M2–M3 (экз. ПИН, № 5644/743); три неполных черепа: с правыми и левыми C1–M3 (экз. ПИН, №№ 5644/744, 745), с правыми C1–M1 и левыми C1–P4 (экз. ПИН, № 5644/746); три фрагмента верхнечелюстных костей: с P4–M3 (экз. ПИН, № 5644/747), с P4–M1 (экз. ПИН, № 5644/748), с P4 (экз. ПИН, № 5644/749); пять полных или почти полных нижнечелюстных костей: с I2–M3 и целой восходящей ветвью (экз. ПИН, № 5644/750), с I1–I2 и C1–M3 (экз. ПИН, № 5644/751), с P4–M3 (экз. ПИН, № 5644/752), с M3 (экз. ПИН, № 5644/753), без зубов (экз. ПИН, № 5644/754); два фрагмента нижнечелюстных костей: с M2–M3 (экз. ПИН, № 5644/755), без зубов (экз. ПИН, № 5644/756); четыре изолированных зуба: два C1 (экз. ПИН, №№ 5644/757, 758), P4 (экз. ПИН, № 5644/759) и M1 (экз. ПИН, № 5644/760).
Список литературы
Лопатин А.В. Раннеплейстоценовый подковонос Rhinolophus macrorhinus cimmerius subsp. nov. (Rhinolophidae, Chiroptera) из пещеры Таврида в Крыму // Доклады Российской академии наук. Науки о жизни. 2022. Т. 506. С. 396–405.
Лопатин А.В., Вислобокова И.А., Лавров А.В., и др. Пещера Таврида – новое местонахождение раннеплейстоценовых позвоночных в Крыму // Доклады Академии наук. 2019. Т. 485. № 3. С. 381–385.
Лопатин А.В., Тесаков А.С. Раннеплейстоценовая белозубка Crocidura kornfeldi (Lipotyphla, Soricidae) из Крыма // Доклады Российской академии наук. Науки о жизни. 2021. Т. 501. № 1. С. 499–504.
Gunnell G.F., Eiting T.P., Geraads D. New late Pliocene bats (Chiroptera) from Ahl al Oughlam, Morocco // Neues Jahrbuch für Geologie und Paläontologie. Abhandlungen. 2011. V. 260. № 1. P. 55–71.
Kormos T. Az 1913. évben végzett ásatásaim eredményei // A Magyar Királyi Földtani Intézet 1913. évi jelentése. Budapest, 1914. P. 498–505.
Kormos T. Diagnosen neuer Säugetiere aus der Oberpliozänen Fauna des Somlyóberges bei Püspökfürdő // Annales Musei Nationalis Hungarici. 1930. V. 27. P. 237–246.
Topál G. Bats from the Lowermost Pleistocene Locality 15 at Beremend, Hungary (Mammalia, Chiroptera) // Fragmenta Mineralogica et Palaeontologica. 1985. V. 12. P. 51–57.
Ito K., Tu V.T., Eiting T.P., et al. On the embryonic development of the nasal turbinals and their homology in bats // Frontiers in Cell and Developmental Biology. 2021. V. 9. Art. 613545. P. 1–19.
Christian J.J. The natural history of a summer aggregation of the big brown bat, Eptesicus fuscus fuscus // American Midland Naturalist. 1956. V. 55. № 1. P. 66–95.
Gol’din P., Godlevska L., Ghazali M. Age-related changes in the teeth of two bat species: dental wear, pulp cavity // Acta Chiropterologica. 2018. V. 20. № 2. P. 519–530.
Rosina V.V., Sinitsa M.V. Bats (Chiroptera, Mammalia) from the Turolian of the Ukraine: phylogenetic and biostratigraphic considerations // Neues Jahrbuch für Geologie und Paläontologie. Abhandlungen. 2014. V. 272. № 2. P. 147–166.
Barti L. Istoricul cercetărilor chiropterologice de pe teritoriul României contemporane şi baza datelor faunistice de la începuturi până în 1944 // Nymphaea. Folia naturae Bihariae. 2005. V. 32. P. 53–114.
Pálfy J., Dulai A., Gasparik M., et al. Catalogue of invertebrate and vertebrate paleontological type specimens of the Hungarian Natural History Museum. Budapest: Hungarian Natural History Museum, 2008.
Kretzoi M. Die Nager und Lagomorphen von Voigtstedt in Thüringen und ihre chronologische Aussage // Paläontologische Abhandlungen. Abteilung A. 1965. V. 2. № 2–3. P. 585–661.
Rabeder G. Weitere Grabungsergebnisse von der altpleistozänen Wirbeltierfundstelle Deutsch-Altenburg 2 // Die Hohle. 1973. V. 24. № 1–4. P. 8–15.
Storch G. Quartäre Fledermaus-Faunen von der Insel Malta // Senckenbergiana Lethaea. 1974. V. 55. P. 407–434.
Tata C., Kotsakis T. Italian fossil chiropteran assemblages: a preliminary report // Geo.Alp. 2005. V. 2. P. 53–60.
Sevilla García P. Estudio paleontológico de los Quirópteros del Cuaternario español // Paleontologia i Evolució. 1988. V. 22. P. 113–233.
Wołoszyn B.W. Pliocene and Pleistocene bats of Poland // Acta Palaeontologica Polonica. 1987. V. 32. № 3–4. P. 207–325.
Pereswiet-Soltan A. First record of serotine bat Eptesicus serotinus (Chiroptera: Vespertilionidae) from Early-Middle Holocene in southern Europe (Boeotia – Greece) // Hystrix. Italian Journal of Mammalogy. 2016. V. 27. № 2. P. 209–211.
Дополнительные материалы отсутствуют.
Инструменты
Доклады Российской академии наук. Науки о жизни