

Доклады Российской академии наук. Науки о жизни, 2023, T. 509, № 1, стр. 109-114
Rhinolophus mehelyi scythotauricus subsp. nov. (Rhinolophidae, Chiroptera) из нижнего плейстоцена пещеры Таврида в Крыму
Академик РАН А. В. Лопатин 1, *
1 Палеонтологический институт им. А.А. Борисяка Российской академии наук
Москва, Россия
* E-mail: alopat@paleo.ru
Поступила в редакцию 22.11.2022
После доработки 25.11.2022
Принята к публикации 26.11.2022
- EDN: NAQJQC
- DOI: 10.31857/S2686738922600947
Аннотация
По неполному черепу из нижнеплейстоценовых отложений пещеры Таврида в центральном Крыму описан новый вымерший подвид подковоносов Мегели, Rhinolophus mehelyi scythotauricus subsp. nov., самый крупный представитель группы R. euryale. По эволюционному уровню он занимает промежуточное положение между плио-плейстоценовым R. mehelyi birzebbugensis Storch, 1974 и современными представителями вида, но его крупные размеры и относительно узкие верхние моляры могут свидетельствовать о принадлежности к особой филогенетической линии в пределах R. mehelyi Matschie, 1901. R. mehelyi scythotauricus subsp. nov. является первой ископаемой находкой вида в Крыму; также это одна из наиболее северных находок R. mehelyi.
Коллекция раннеплейстоценовых млекопитающих из пещеры Таврида в центральном Крыму (Белогорский район, пос. Зуя) включает многочисленные остатки рукокрылых семейств Rhino-lophidae и Vespertilionidae [1, 2]. Помимо остатков крупных подковоносов Rhinolophus macrorhinus cimmerius Lopatin, 2022 из группы R. ferrumequinum [1] в сборах 2022 г. имеется неполный череп еще одного представителя Rhinolophidae – описываемого ниже Rhinolophus mehelyi scythotauricus subsp. nov. из группы R. euryale.
Фаунистический комплекс пещеры Таврида датируется поздним виллафранком, около 1.8–1.5 млн л.н. [3, 4].
Материал хранится в Палеонтологическом институте им. А.А. Борисяка РАН (ПИН) в г. Москве. Методика измерений черепа по [5]. Терминология строения зубов по [6]. Иллюстрации выполнены с помощью цифрового фотоаппарата Nikon D800 с объективом AF-S Micro NIKKOR 60mm f/2.8G ED, рентгеновского микротомографа Neoscan N80 и сканирующего электронного микроскопа Tescan Vega 3 XMU в ПИН.
Отряд Chiroptera Blumenbach, 1779
Семейство Rhinolophidae Gray, 1825
Род Rhinolophus Lacépède, 1799
Rhinolophus mehelyi Matschie, 1901
Rhinolophus mehelyi scythotauricus Lopatin, subsp. nov.
Название подвида по античной Крымской Скифии и племени скифотавров (тавроскифов).
Голотип – ПИН, № 5644/770, неполный череп с правыми C1–M3 и левыми C1, P4–M3; Россия, Республика Крым, Белогорский район, местонахождение Таврида (точка “Логово гиен”); нижний плейстоцен.
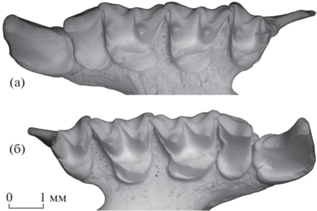
Описание (рис. 1–3). Размеры относительно крупные, несколько меньше, чем у современного R. ferrumequinum (Schreber, 1774). Ростральный отдел черепа сравнительно узкий, клыки слабо разнесены (рис. 1а, 1б, 2а). Носовое вздутие хорошо развито, спереди немного выступает за край носовой вырезки и нависает над носовым отверстием (рис. 1в, 1г). Парные срединные вздутия плохо обособлены друг от друга, парные боковые вздутия слабо выражены. Надглазничные гребни резкие, ограниченная ими сзади ростральная (лобная) впадина отчетливо углубленная. Сагиттальный гребень хорошо развит. Межглазничный промежуток узкий.
Рис. 1.
Rhinolophus mehelyi scythotauricus Lopatin, subsp. nov., голотип ПИН, № 5644/770, неполный череп с правыми C1–M3 и левыми C1, P4–M3: а – с дорсальной стороны, б – с вентральной стороны, в – с правой латеральной стороны, г – с левой латеральной стороны, д – с ростральной стороны; Россия, Крым, пещера Таврида; нижний плейстоцен.
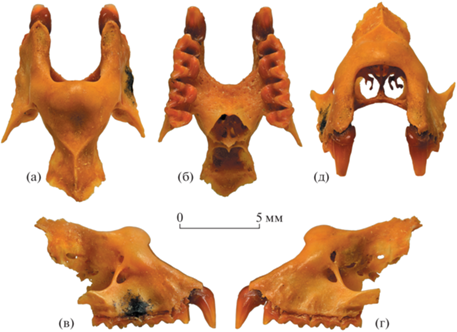
Рис. 2.
Rhinolophus mehelyi scythotauricus Lopatin, subsp. nov., голотип ПИН, № 5644/770, неполный череп с правыми C1–M3 и левыми C1, P4–M3: а – с вентральной стороны; б – носовая полость с ростральной стороны (латеральный тяж правого комплекса турбиналий отломан); в – томограмма носовой полости, дорсальная проекция; г – томограмма левой половины носовой полости, сагиттальная проекция (в антеровентральной части комплекса турбиналий видны крупные костные тяжи, более длинный латеральный тяж сильнее выступает дистально; угловатый инородный объект над ними – мелкий фрагмент породы); Россия, Крым, пещера Таврида; нижний плейстоцен.
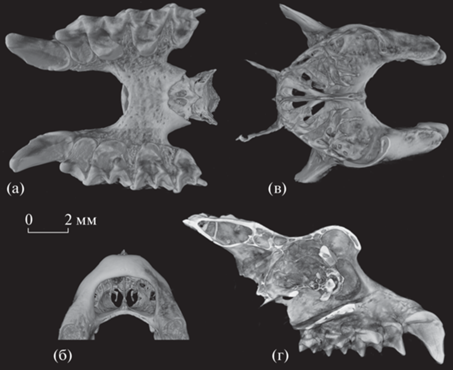
Рис. 3.
Rhinolophus mehelyi scythotauricus Lopatin, subsp. nov., голотип ПИН, № 5644/770, верхние зубы с окклюзиальной стороны: а – левые C1, P4–M3; б – правые С1–M3; Россия, Крым, пещера Таврида; нижний плейстоцен.
Подглазничное отверстие крупное, округло-треугольной формы. Костный мостик, отделяющий подглазничное отверстие от глазницы, тонкий в вентральной части, но значительно расширяется дорсально, непосредственно под слезным отверстием. Слезное отверстие крупное, овальное, расположено на уровне середины M2 (рис. 1в, 1г). Скуловая дуга в передней части низкая и тонкая.
Небный мостик очень короткий (длина без переднего и заднего выступов составляет около 27% от длины верхнечелюстного зубного ряда C1–M3), его передний край расположен на уровне контакта метастиля M1 и парастиля M2, а задний край находится на уровне парастиля M3.
Носовое отверстие субпрямоугольной формы (рис. 1д, 2б). В антеровентральной части комплекса турбиналий обособлены два резко изогнутых тонких костных тяжа – медиальный и латеральный (рис. 2в, 2г). Они относительно короткие и демонстрируют строение, близкое таковому современного R. mehelyi ([7], рис. 6B; [8], рис. 2). Сравнительно небольшой медиальный тяж имеет узкий лопастевидный вентральный отросток. Конец удлиненной дистальной порции латерального тяжа достигает уровня границы M2/M3 и находится существенно мезиальнее заднего края небного мостика (рис. 2г).
Ряд C1–P4 по длине в 1.6–1.7 раза уступает ряду M1–M3.
Верхний клык С1 относительно тонкий и невысокий. Вершина слегка загнута дистально. Задний режущий гребень мощный, с широкой фасеткой стирания (рис. 3а); антеролингвальное и лабиальное ребра тонкие. Основание антеролингвального ребра немного вентральнее цингулюма формирует валикообразное вертикальное утолщение; апикальнее него по ребру тянется плоская фасетка стирания, из-за которой выпуклое окончание указанного утолщения выглядит как небольшой скошенный зубец в антеролингвальном углу коронки C1. Задний дополнительный бугорок хорошо развит. Основание коронки овальное, с небольшим постеролингвальным выступом. Постеролабиальный и постеролингвальный края основания коронки почти прямые, с едва намеченными впадинами (рис. 3б). Цингулюм широкий, непрерывный, волнистый, на антеролабиальной стороне резко изогнут апикально.
Рудиментарный однокорневой верхний малый премоляр P2 с округлой уплощенной коронкой (рис. 3б). Он смещен к лабиальному краю зубного ряда и не препятствует контакту C1 и P4 (однако между этими зубами имеется узкая щель). Главный бугорок P2 очень маленький и низкий, с его едва возвышенной вершиной связаны два коротких и тонких субпродольных гребня – передний и задний. Имеется слабый непрерывный цингулюм.
Верхний большой премоляр P4 короткий и широкий, с крупным талоном и легкой задней вырезкой (рис. 3б). Парастиль смещен базально и крайне редуцирован (выражен лишь закруглением цингулюма), парастилярный выступ практически отсутствует. Цингулюм непрерывный, узкий на всем протяжении, но заметно расширен в постеролингвальной части.
Верхние моляры M1 и M2 относительно узкие, с очень слабой полкой гипокона, более заметной на M1. Прецингулюм и постцингулюм очень тонкие. Задняя вырезка слабая.
M3 мелкий, субтреугольный, с большим пара-стилярным выступом. Метастиль отсутствует, метакон хорошо развит. Прецингулюм короткий и узкий, постцингулюм очень слабо выражен в основании постеролингвальной стороны коронки.
Размеры голотипа в мм. Длина небного мостика – 2.1; высота носового вздутия (от переднего края небного мостика до вершины) – 3.7; ширина носового отверстия – 3.1; межглазничная ширина – 2.6; ширина на уровне C1–C1 – 5.75; ширина на уровне M2–M2 – 7.85; ширина на уровне M3–M3 – 8.1; длина C1–M3 (dex – правый, sin – левый) – 7.7 (dex), 7.65 (sin); длина C1–P4 – 3.1 (dex), 3.0 (sin); длина P4–M3 – 5.9 (dex), 5.95 (sin); длина M1–M3 – 5.0 (dex, sin); наименьшее расстояние между C1 и P4 – 0.05 (dex, sin). Размеры зубов (L – длина, W – ширина, H – высота): C1 (dex): L – 1.85, W – 1.35, H – 2.4; C1 (sin): L – 1.85, W – 1.3, H – 2.4; P2 (dex): L – 0.35, W – 0.4; P4 (dex, sin): L – 1.1, W – 1.9; M1 (dex, sin): L – 2.0, W – 2.0; M2 (dex, sin): L – 1.9, W – 2.05; M3 (dex, sin): L – 1.25, W – 1.85.
Сравнение и замечания. Строение лицевой части черепа, костного неба, турбиналий и верхних зубных рядов (длинные и узкие верхние моляры, слабо развитые полки гипокона M1–M2) свидетельствуют о принадлежности описываемой формы к виду Rhinolophus mehelyi. При этом величиной она существенно превосходит современные подвиды R. mehelyi mehelyi Matschie, 1901 (Южная Европа, Западная Азия) и R. mehelyi tuneti Deleuil et Labbé, 1955 (Северная Африка) и сближается с R. mehelyi birzebbugensis Storch, 1974 из плиоцена Болгарии (MN15b) и раннего плейстоцена Мальты [9, 10].
Rhinolophus mehelyi scythotauricus subsp. nov. немного крупнее R. mehelyi birzebbugensis (табл. 1). У R. mehelyi birzebbugensis с Мальты длина M1–M3 составляет 4.6 мм ([9], табл. 3) против 5.0 мм у нового подвида из пещеры Таврида, при этом по длине верхних моляров экземпляры R. mehelyi birzebbugensis из Болгарии в среднем уступают крымской форме, а по максимальным значениям почти тождественны ей ([10], табл. 2). Соответственно, новая форма существенно превосходит по величине всех других представителей группы R. euryale, включая позднемиоценового R. antonioi Crespo et al., 2017 [11] и плиоценового R. neglectus Heller, 1936 [12, 13], а также вымерших и современных R. euryale Blasius, 1853. Таким образом, Rhinolophus mehelyi scythotauricus subsp. nov. – самый крупный представитель группы R. euryale, приближающийся по размерам к R. ferrumequinum. Несколько более крупный R. maghrebensis Gunnell, Eiting et Geraads, 2011 из терминального плиоцена Марокко ([6], табл. 1), иногда предположительно рассматривающийся в составе группы R. euryale [11, 14], явно принадлежит к группе R. ferrumequinum (см. [6]).
Таблица 1.
Размеры (в мм) верхних зубов Rhinolophus mehelyi Matschie, 1901 (n – число экземпляров)
| Параметры | R. mehelyi scythotauricus subsp. nov. | R. mehelyi birzebbugensis [10] | R. mehelyi mehelyi [10] | ||||
|---|---|---|---|---|---|---|---|
| голотип | n | пределы | среднее | n | пределы | среднее | |
| Длина C1 | 1.85; 1.85 | 213 | 1.6–2.0 | 1.72 | 14 | 1.45–1.75 | 1.57 |
| Ширина C1 | 1.3; 1.35 | 213 | 1.1–1.57 | 1.31 | 14 | 1.1–1.32 | 1.24 |
| Длина P4 | 1.1; 1.1 | 3 | 0.97–1.27 | 1.14 | 14 | 1.02–1.2 | 1.09 |
| Ширина P4 | 1.9; 1.9 | 3 | 1.7–1.97 | 1.85 | 14 | 1.62–1.97 | 1.8 |
| Длина M1 | 2.0; 2.0 | 33 | 1.67–1.95 | 1.81 | 14 | 1.62–1.7 | 1.66 |
| Ширина M1 | 2.0; 2.0 | 33 | 1.82–2.2 | 2.06 | 14 | 1.72–2.0 | 1.9 |
| Длина M2 | 1.9; 1.9 | 45 | 1.6–1.85 | 1.72 | 14 | 1.5–1.62 | 1.56 |
| Ширина M2 | 2.05; 2.05 | 45 | 1.87–2.3 | 2.03 | 14 | 1.7–2.07 | 1.85 |
| Длина M3 | 1.25; 1.25 | 20 | 1.15–1.27 | 1.19 | 14 | 1.05–1.25 | 1.13 |
| Ширина M3 | 1.85; 1.85 | 19 | 1.75–1.95 | 1.84 | 14 | 1.57–1.7 | 1.63 |
Морфологически R. mehelyi scythotauricus subsp. nov. отличается от R. mehelyi birzebbugensis более коротким небным мостиком, сильнее смещенным лабиально рудиментарным P2, редуцированным парастилем P4 и менее расширенными M1–M2 (см. [9, 10]). В целом можно заключить, что по эволюционному уровню новый подвид занимает промежуточное положение между R. mehelyi birzebbugensis и современными представителями вида, хотя его крупные размеры и относительно узкие верхние моляры могут свидетельствовать о принадлежности к отдельной филогенетической линии в пределах R. mehelyi.
Различия Rhinolophus mehelyi scythotauricus subsp. nov. с совместно встречающимся в нижнеплейстоценовых отложениях пещеры Таврида R. macrorhinus cimmerius Lopatin, 2022 из группы R. ferrumequinum включают заметно более мелкие общие размеры (см. [1], табл. 1), существенно иное строение верхних клыков и моляров (у R. macrorhinus cimmerius клыки более массивные и крупные, с четкой постеролингвальной впадиной в основании, M1–M2 сильно расширенные, в том числе за счет очень мощно развитой полки гипокона, а M3 более длинный и широкий), крайнюю редукцию парастиля P4, менее массивную ростральную часть черепа, более короткий небный мостик, широкое овальное слезное отверстие, субпрямоугольное носовое отверстие и нависающее над краем носовой вырезки носовое вздутие (у R. macrorhinus cimmerius слезное отверстие щелевидное, носовое отверстие полукруглое, а носовое вздутие не нависает над краем носовой вырезки). Кроме того, R. mehelyi scythotauricus subsp. nov. сильно отличается от R. macrorhinus cimmerius строением костных тяжей антеровентральной части комплекса турбиналий (см. [1], рис. 3в), а именно узкой вентральной лопастью медиального тяжа и существенно меньшей длиной латерального тяжа.
Наиболее древние ископаемые остатки современного подвида Rhinolophus mehelyi mehelyi известны из среднего плейстоцена Южной Европы и Передней Азии [14, 15]; также имеются сведения о находках R. mehelyi совместно с R. macrorhinus в верхах нижнего плейстоцена южной Венгрии [16]. Ныне живущие подковоносы Мегели добывают пищу (преимущественно, чешуекрылых) как в лесных, так и в открытых обстановках, селятся в карстовых пещерах вблизи водоемов [15, 17, 18]. Нередко они образуют большие колонии, иногда совместно с другими видами летучих мышей близкого размера [19]. Вероятно, редкость находок R. mehelyi scythotauricus subsp. nov. (по сравнению с R. macrorhinus cimmerius) в отложениях пещеры Таврида связана с локальными особенностями распределения колоний обоих видов на разных участках этой большой пещеры в раннем плейстоцене.
Rhinolophus mehelyi scythotauricus subsp. nov. представляет собой первую ископаемую находку вида в Крыму (о сомнительных регистрациях современных представителей см. [20]); также это одна из наиболее северных находок R. mehelyi (см. [19]).
Материал. Голотип.
Список литературы
Лопатин А.В. Раннеплейстоценовый подковонос Rhinolophus macrorhinus cimmerius subsp. nov. (Rhinolophidae, Chiroptera) из пещеры Таврида в Крыму // Доклады Российской академии наук. Науки о жизни. 2022. Т. 506. С. 396–405.
Лопатин А.В. Раннеплейстоценовый кожан Eptesicus praeglacialis (Vespertilionidae, Chiroptera) из пещеры Таврида в Крыму // Доклады Российской академии наук. Науки о жизни. 2023. Т. 508. С. 95–104.
Лопатин А.В., Вислобокова И.А., Лавров А.В. и др. Пещера Таврида – новое местонахождение раннеплейстоценовых позвоночных в Крыму // Доклады Академии наук. 2019. Т. 485. № 3. С. 381–385.
Лопатин А.В., Тесаков А.С. Раннеплейстоценовая белозубка Crocidura kornfeldi (Lipotyphla, Soricidae) из Крыма // Доклады Российской академии наук. Науки о жизни. 2021. Т. 501. № 1. С. 499–504.
Topál G., Csorba G. The subspecific division of Rhino-lophus luctus Temminck, 1835, and the taxonomic status of R. beddomei Andersen, 1905 (Mammalia, Chiroptera) // Miscellanea Zoologica Hungarica. 1992. V. 7. P. 101–116.
Gunnell G.F., Eiting T.P., Geraads D. New late Pliocene bats (Chiroptera) from Ahl al Oughlam, Morocco // Neues Jahrbuch für Geologie und Paläontologie. Abhandlungen. 2011. V. 260. № 1. P. 55–71.
Curtis A.A., Simmons N.B. Unique turbinal morphology in horseshoe bats (Chiroptera: Rhinolophidae) // The Anatomical Record. 2017. V. 300. № 2. P. 309–325.
Curtis A.A., Smith T.D., Bhatnagar K.P., et al. Maxilloturbinal aids in nasophonation in horseshoe bats (Chiroptera: Rhinolophidae) // The Anatomical Record. 2020. V. 303. № 1. P. 110–128.
Storch G. Quartäre Fledermaus-Faunen von der Insel Malta // Senckenbergiana Lethaea. 1974. V. 55. P. 407–434.
Popov V. Pliocene small mammals (Mammalia, Lipotyphla, Chiroptera, Lagomorpha, Rodentia) from Muselievo (North Bulgaria) // Geodiversitas. 2004. V. 26. № 3. P. 403–491.
Crespo V.D., Sevilla P., Mansino S., et al. Bats from the classical site of Venta del Moro (Late Miocene, Eastern Spain) // Historical Biology. 2018 (2017). V. 30. № 3. P. 317–326.
Heller F. Eine oberpliocäne Wirbeltierfauna aus Rheinhessen // Neues Jahrbuch für Mineralogie, Geologie und Paläontologie. Abteilung B. 1936. V. 76. P. 99–160.
Wołoszyn B.W. Pliocene and Pleistocene bats of Poland // Acta Palaeontologica Polonica. 1987. V. 32. № 3–4. P. 207–325.
Horáček I., Maul L.C., Smith K.T., et al. Bat remains (Mammalia, Chiroptera) from the Middle Pleistocene site of Qesem Cave, Israel, with the first Pleistocene record of fruit bats in the Mediterranean region // Palaeontologia Electronica. 2013. V. 16. № 3 (23A). P. 1–19; palaeo-electronica.org/content/2013/513-qesem-bats.
Salari L., Agnelli P., Calcagnile L. The fossil bat assemblages from the Grotta dei Pipistrelli in Pantalica (southeastern Sicily, Italy): chronological and palaeoecological implications // Comptes Rendus Palevol. 2019. V. 18. № 4. P. 417–441.
Pazonyi P., Trembeczki M., Mészáros L., Szentesi Z. Preliminary report on the Early Pleistocene vertebrate sites of Beremend Crystal Cave (Beremend 16, South Hungary) and on their palaeoecological importance // Fragmenta Palaeontologica Hungarica. 2019. V. 36. P. 115–140.
Dietz C., von Helversen O., Nill D. Bats of Britain, Europe and Northwest Africa. London: A&C Black, 2009. 399 p.
Dondini G., Tomassini A., Inguscio S., Rossi E. Rediscovery of Mehely’s horseshoe bat (Rhinolophus mehelyi) in peninsular Italy // Hystrix. Italian Journal of Mammalogy. 2014. V. 25. № 1. P. 59–60.
Csősz I., Jére C., Bücs S., et al. The presence of Mehely’s horseshoe bat Rhinolophus mehelyi in South-Western Romania // North-Western Journal of Zoology. 2015. V. 11. № 2. P. 351–356.
Иваницкий А.Н. Подковонос Мегели Rhinolophus mehelyi Matschie, 1901 // Красная книга Российской Федерации. 2 изд. Москва: ФГБУ ВНИИ Экология, 2021. С. 942–943.
Дополнительные материалы отсутствуют.
Инструменты
Доклады Российской академии наук. Науки о жизни