

Доклады Российской академии наук. Науки о жизни, 2023, T. 509, № 1, стр. 115-118
Новые данные по гесперорнисам (Aves: Ornithurae) кампана Нижнего Поволжья (поздний мел, Россия)
Н. В. Зеленков 1, *, М. С. Архангельский 2, 3
1 Палеонтологический институт
им. А.А. Борисяка Российской академии наук
Москва, Россия
2 Саратовский государственный университет
Саратов, Россия
3 Саратовский государственный технический университет
Саратов, Россия
* E-mail: nzelen@paleo.ru
Поступила в редакцию 13.10.2022
После доработки 20.10.2022
Принята к публикации 20.10.2022
- EDN: NBFBNO
- DOI: 10.31857/S2686738922700123
Аннотация
Представления о таксономическом разнообразии позднемеловых гесперорнисов (Aves: Hesperornithidae) Европейской части России и Восточной Европы в целом остаются слабо разработанными, а морфология этих крупных нелетающих птиц – плохо изученной. Новые находки Hesperornithidae в местонахождении Карякино (Саратовская область, Россия) подтверждают сосуществование двух форм этих нелетающих морских птиц в кампане (середина позднего мела) Нижнего Поволжья. Для российского гесперорниса (Hesperornis rossicus Nessov et Yarkov, 1993) впервые описана бедренная кость – она указывает на выраженную морфологическую обособленность этого крупного вида от североамериканского H. regalis Marsh, 1872.
Гесперорнисы (семейство Hesperornithidae вымершего отряда Hesperornithiformes) – очень крупные утратившие способность к полету ныряющие птицы, населявшие моря Северного полушария во второй половине позднего мела. Гесперорнисы хорошо известны в палеонтологической летописи Северной Америки, но в Евразии их остатки очень редки, вследствие чего представления о таксономическом разнообразии и морфологии восточноевропейских и западноазиатских Hesperornithidae остаются очень плохо разработанными [1–3]. В связи с возможным голарктическим распространением отдельных видов Hesperornithiformes [4] остается неясным статус всех описанных с территории Евразии видов [3]. Так, очень крупные гесперорнисы из кампана (поздний мел) Восточной Европы чаще всего обозначаются как Hesperornis rossicus [5], однако валидность этого вида, известного по очень фрагментарным находкам, ставится под сомнение [1]. Иногда предполагается существование, по меньшей мере, еще одного вида гесперорнисов в кампане Восточной Европы [3, 6]. При этом из кампана Северной Америки описано до десяти видов Hesperornithiformes [2]. Рис. 1
Рис. 1.
Новые материалы по гесперорнисам (Aves: Hesperornithidae) из кампана местонахождения Карякино (Нижнее Поволжье, Саратовская область; верхний мел): а–в – Hesperornis rossicus Nessov et Yarkov, 1993, фрагментарная правая бедренная кость, экз. ПИН, № 5027/8, с каудальной (а), краниальной (б) и дистальной (в) сторон; г–е – Hesperornis sp., фрагментарный правый тибиотарсус, экз. ПИН, № 5027/9, с краниальной (г), каудальной (д) и дистальной (е) сторон. Обозначения: ai – apophysis internum ligament obliqui; cm – condylus medialis; cl – condylus lateralis; ctl – crista trochlearis lateralis; fp – fossa poplitea; ii – incisura intercondylaris; si – sulcus intercondylaris; sp – sulcus patellaris; tf – trochlea fibularis.
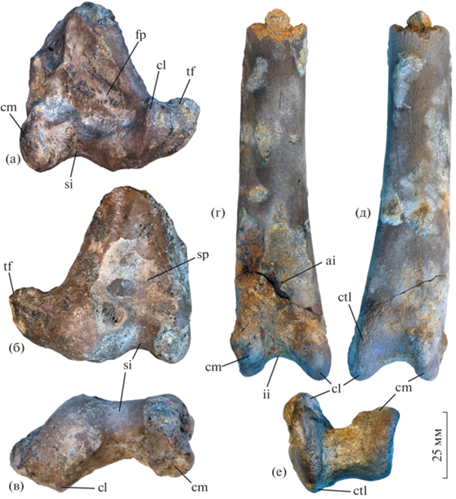
В этой работе описаны новые находки костей посткраниального скелета гесперорнисов из кампанского местонахождения Карякино (Саратовская область, Россия), откуда ранее указывались H. rossicus и, предположительно, еще один неназванный гесперорнис [3, 5]. Фрагментарные бедренная кость и тибиотарсус принадлежат птицам двух размерных классов и подтверждают существование двух форм Hesperornithidae в кампане Нижнего Поволжья. Бедренная кость, имеющая высокую диагностическую ценность у Hesperornithidae, здесь впервые описана для восточноевропейских гесперорнисов.
Дистальная половина бедренной кости (экз. ПИН, № 5027/8) принадлежит очень крупной птице, по размеру несколько превышающей североамериканских Hesperornis regalis. Размеры экз. ПИН, № 5027/8: максимальная ширина дистального эпифиза – 65.2 мм; ширина стержня на уровне проксимального края condylus lateralis ~41.2 мм; наименьшая краниокаудальная высота стержня – 17.8 мм; краниокаудальная высота condylus medialis – 32.5 мм; минимальная краниокаудальная высота sulcus intercondylaris – 18.4 мм. Этот образец характеризуется выраженно краниокаудально уплощенным и медиолатерально расширенным стержнем, очень мелкой и широкой sulcus patellaris, не доходящей дистально до sulcus intercondylaris, а также значительно выдвинутым латерально condylus lateralis вместе с trochlea fibularis. По сочетанию этих признаков описываемый образец сходен с представителями родов Hesperornis и Parahesperornis, но отличается от других Hesperornithiformes [7]. Бедренную кость из Карякино сближает с таковой Hesperornis, помимо крупных размеров, сильно выставленный латерально trochlea fibularis, крупная (простирающаяся практически до латеральной кромки стержня) fossa poplitea, а также в равной степени выдающиеся дистально condylae medialis et lateralis – у представителей рода Parahesperornis condylus lateralis выдвинут дистальнее, чем condylus medialis, а trochlea fibularis выступает латерально лишь умеренно [7].
Экз. ПИН, № 5027/8 в целом схож с бедренной костью Hesperornis regalis, но отличается несколько более крупными размерами, а также заметно сильнее выдающимся латерально и более узким проксимодистально trochlea fibularis, в результате чего дистальный эпифиз выглядит в целом более расширенным, чем у вышеупомянутого североамериканского вида. Проксимальная вершина condylus lateralis также сильнее отставлена латерально у экз. ПИН, № 5027/8, чем у H. regalis. Перегиб в дистальном крае кости между вершиной condylus lateralis и trochlea fibularis практически отсутствует. При виде с каудальной стороны костный валик, ограничивающий дистально fossa poplitea, у описываемого экземпляра уже и медиолатерально длиннее, чем у H. regalis. При виде с дистальной стороны дорсальная часть sulcus patellaris формирует слабовыраженную вырезку, которая у H. regalis явственно более вогнутая. Также при виде с дистальной стороны каудальный край condylus lateralis ориентирован заметно более полого (под небольшим углом к поперечной оси кости), чем у H. regalis, у которого этот мыщелок ориентирован более наклонно. Морфологическая дистинктивность данной бедренной кости, наряду с крупными размерами, позволяет относить ее к H. rossicus и, таким образом, подтверждает валидность этого вида, ранее установленного по тарсометатарсусу, также отличающемуся от такового H. regalis крупными размерами и морфологически. Подтвержденное присутствие H. rossicus в фауне местонахождения Карякино позволяет относить к этому виду также и описанный ранее [3] очень крупный фрагментарный тибиотарсус (экз. ЗИН, PO 6611), по размерам несколько превышающий таковые H. regalis.
Дистальная половина правого тибиотарсуса (экз. ПИН, № 5027/9) из Карякино отличается от ранее описанного экз. ЗИН, PO 6611 из этого же местонахождения [3] заметно меньшими размерами и более глубокой и проксимально расширенной incisura intercondylaris. При виде с дистальной стороны этот экземпляр, напротив, имеет дорсовентрально широкую костную перегородку между мыщелками. Плохая сохранность мыщелков у экз. ЗИН, PO 6611 не позволяет сравнить два экземпляра по строению суставных поверхностей. Размеры экз. ПИН № 5027/9: ширина стержня на уровне бугристости apophysis internum ligament obliqui [см.: 8] – 28.5 мм, краниокаудальная высота стержня в его проксимальной половине ~13.8 мм; поперечная ширина дистального эпифиза – 31.1 мм; краниокаудальная высота через condylus medialis – 21.2 мм; краниокаудальная высота через condylus lateralis – 30.6 мм. На основании размерных и морфологических отличий этот экземпляр не может быть отнесен к H. rossicus и, очевидно, представляет отдельный вид гесперорнисов. По размерам он соответствует североамериканскому H. regalis, но несколько отличается от него дистальным профилем кости. У экз. ПИН, № 5027/9 condylus lateralis существенно выступает краниально относительно condylus medialis, в то время как у H. regalis разница в краниальном выступе мыщелков выражена значительно слабее. Crista trochlearis lateralis мыщелка, напротив, заметно выдается относительно эпифиза у H. regalis, в то время как у экз. ПИН, № 5027/9 этот гребень менее выражен. Для других североамериканских видов Hesperornis дистальные тибиотарсусы не описаны.
Таким образом, новые находки в местонахождении Карякино подтверждают сосуществование в кампане на территории Нижнего Поволжья, по меньшей мере, двух видов гесперорнисов: очень крупного H. rossicus и более мелкой формы, по размеру сходной с H. regalis, но отличающейся от этого североамериканского вида морфологически. Помимо описанного выше тибиотарсуса экз. ПИН, № 5027/9 к более мелкой форме также могут быть отнесены описанные ранее фрагментарные тарсометатарсусы из Карякино [3, 5], которые отличаются от H. rossicus несколько меньшими размерами, но при этом также проявляют отличия и от крупных североамериканских видов H. regalis и H. chowi. Некоторые авторы [9, 10] не исключали возможность существования у гесперорнисов полового диморфизма, что могло бы объяснить наблюдаемую разницу в размерах у кампанских восточноевропейских гесперорнисов, однако присутствие двух морфологических типов тарсометатарсусов [3] поддерживает представления о сосуществовании двух таксонов. Несколько более мелкая форма кампанских гесперорнисов из Поволжья, по-видимому, требует выделения в отдельный вид [3], но мы воздерживаемся от его формального описания до ревизии североамериканских видов и получения более репрезентативных материалов по евроазиатским формам.
Список литературы
Bell A., Chiappe L.M. // J. Syst. Palaeontol. 2016. V. 14. P. 239–251.
Bell A., Chiappe L.M. // Diversity. 2022. V. 14. P. 267.
Zelenkov N.V., Panteleyev A.V., Yarkov A.A. // Paleontol. J. 2017. V. 51. P. 547–555.
Aotsuka K., Sato T. // Cret. Res. 2016. V. 63. P. 154–169.
Panteleyev A.V., Popov E.V., Averianov A.O. // Paleontol. Res. 2004. V. 8. P. 115–122.
Зеленков Н.В., Курочкин Е.Н. В кн.: Ископаемые рептилии и птицы. Часть 3. М.: Геос, 2015. С. 86–290.
Bell A., Chiappe L.M. // Life. 2020. V. 10. P. 62.
Зеленков Н.В., В кн.: Ископаемые рептилии и птицы. Часть 3. М.: Геос, 2015. С. 61–85.
Galton P.M., Martin L.D. In: Mesozoic birds: Above the heads of dinosaurs. Berkeley: Univ. California Pres, 2002. P. 489–538.
Bell A., Everhart M.J. // Trans. Kansas Acad. Sci. 2009. V. 112. P. 7–14.
Дополнительные материалы отсутствуют.
Инструменты
Доклады Российской академии наук. Науки о жизни