

Доклады Российской академии наук. Науки о жизни, 2023, T. 510, № 1, стр. 292-296
МОГУТ ЛИ ЗЕМЛЕРОЙКИ ИЗБЕГАТЬ КОНКУРЕНЦИИ ПРИ НАРУШЕНИИ ЭКОЛОГИЧЕСКОГО ПРАВИЛА ХАТЧИНСОНА?
А. Г. Васильев 1, *, академик РАН В. Н. Большаков 1, И. А. Васильева 1, Т. П. Коурова 1
1 Институт экологии растений и животных
Уральского Отделения Российской академии наук
Екатеринбург, Россия
* E-mail: vag@ipae.uran.ru
Поступила в редакцию 12.01.2023
После доработки 20.01.2023
Принята к публикации 20.01.2023
- EDN: QHKMSF
- DOI: 10.31857/S2686738923600012
Аннотация
Установлено, что правило Г.Х. Хатчинсона о соотношении массы и размеров тела и кормодобывающих органов близких видов не полностью соблюдается при сравнении видов землероек рода Sorex. Выявлено сходство центроидных размеров нижней челюсти крупных видов S. isodon и S. araneus. Методами геометрической морфометрии между видами установлены различия формы нижней челюсти, активно используемой при добывании корма. Обнаружены межвидовые различия по мандибулярным индексам, отражающим трофическую специализацию и функциональные особенности мандибул. Поэтому землеройки могут избегать конкуренции и при нарушении правила Хатчинсона, причем не столько благодаря размерам, сколько за счет трансформации нижней челюсти, ее специализации, способов захвата добычи и смены диеты. Следовательно, экологическое правило Хатчинсона – не обязательное, а лишь одно из условий уменьшения конкуренции между близкими видами.
Экологическое правило Г.Х. Хатчинсона [1] для гильдий таксономически близких видов (по принятой нами терминологии – таксоценов [2]) известно с середины прошлого века. Хатчинсон установил эмпирическую закономерность, что соотношение средних размеров тела и размеров кормодобывающих органов между ближайшими по размерам симпатрическими видами одного рода составляет приблизительно 1.28–1.30. Данные размерные соотношения как многократно подтверждались, так и опровергались [см. 3], и их изучение по-прежнему представляет интерес. Вероятными механизмами этого феномена предполагались как сортировка и отбор видов из регионального пула, так и то, что сдвиг размеров тела и кормодобывающих органов симпатрических видов должен снижать их конкуренцию. В XXI в. эти аспекты изучали методами морфометрии на галапагосских вьюрках [4], а также с помощью геометрической морфометрии на саламандрах рода Plethodon [5]. Несмотря на ключевой характер для экологии, проблема фиксированного соотношения размеров близких видов в сообществе, известная как отношение Хатчинсона, до сих пор не считается решенной.
Методы геометрической морфометрии [6, 7] позволяют разделить компоненты изменчивости размеров (по центроидным размерам – CS) и формы (по прокрустовым координатам объектов) и могут быть применены для проверки этого правила на модели кормодобывающего органа – нижней челюсти на примере симпатрических видов таксоцена землероек. Сравнение выборок на основе морфофункциональных мандибулярных индексов [8, 9] позволяет соотнести элементы функциональной экологии видов.
Цель исследования – проверка правила Хатчинсона на примере сравнения размеров и формы нижнечелюстных ветвей у симпатрических видов землероек рода Sorex Висимского заповедника (Средний Урал) и косвенная оценка степени их трофической специализации на основе использования методов геометрической морфометрии и морфофункциональных мандибулярных индексов.
Изучены коллекционные серии черепов сеголеток четырех симпатрических видов бурозубок рода Sorex, добытые в конце XX в. (июль–август) на территории Висимского природного заповедника (Средний Урал): Sorex isodon Turov, 1924 – равнозубая, (n = 28, 1986 г.), S. araneus Linnaeus, 1758 – обыкновенная (n = 50, 1986 г.), S. caecutiens Laxmann, 1788 – средняя (n = 45, 1986–1987 гг.), S. minutus Linnaeus, 1766 – малая (n = 16, 1986–1987, 1989–1992 гг.).
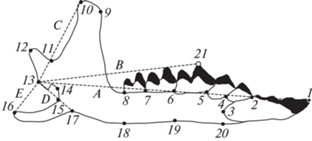
Фотографии нижних челюстей бурозубок получены с помощью планшетного сканера при увеличении 1200 dpi. Для анализа изменчивости размеров и формы нижней челюсти бурозубок использовали 20 меток-ландмарок на буккальной стороне правой ветви нижней челюсти (рис. 1).
Рис. 1.
Схема расстановки основных меток-ландмарок (1–20) и снятия промеров (A, B, C, D, E) для расчета мандибулярных индексов на буккальной стороне мандибулы обыкновенной бурозубки (Sorex araneus). Дополнительная метка 21 использована только для снятия промера B.
Суперимпозицию (superimposition) конфигураций ландмарок (landmarks) осуществили методом генерализованного Прокрустова анализа – GPA [6] с расчетом прокрустовых координат (Procrustes coordinates). Cравнение размеров мандибул выполнили по значениям центроидного размера (centroid size – CS) [6], выраженного в мм. По прокрустовым координатам провели канонический анализ выборок с расчетом матрицы обобщенных расстояний Махаланобиса (D). Парные различия оценили на основе апостериорного Q-теста Тьюки (Tukey’s pairwise post-hoc Q-test).
Расчеты мандибулярных морфофункциональных индексов бурозубок выполнили на основе промеров нижней челюсти (см. рис. 1: A, B, C, D, E) по формулам [8]: TI = C/A (темпорально-резцовый), TM = C/B (темпорально-молярный), MI = D/A (массетерно-резцовый), MM = D/B (массетерно-молярный), AM = E/D (ангулярно-массетерный).
Половые различия центроидных размеров (CS) у сеголеток каждого вида оказались малы и статистически недостоверны, поэтому дальнейшее сравнение проводили без учета пола животных. Результаты сравнения средних CS, косвенно характеризующих размер мандибул у четырех симпатрических видов бурозубок, представлены в табл. 1. Средние CS мандибул двух наиболее крупных видов – S. isodon и S. araneus оказались близки по величине и статистически не различались (Q = 1.197; p = 0.832). Однако они были значимо больше (p < 0.0001), чем у средней и малой бурозубок. Достоверны и различия по CS мандибул между средней и малой бурозубками (Q = 9.088; p < 0.0001). Тест Левене (Levene’s test) не выявил нарушения условий однородности дисперсий CS. При множественном сравнении CS однофакторным дисперсионным анализом (One-Way ANOVA) выявили значимые межвидовые различия (F = 243.8; d.f. = 3, 135; p < 0.0001).
Таблица 1.
Средние центроидные размеры (CS) нижней челюсти видов Sorex (с учетом стандартных ошибок ± SE); отношения Хатчинсона по величинам CS мандибул (треугольная матрица под диагональю); степень “разделения ниш” пар видов ρ (треугольная матрица над диагональю) – отношение разности CS (d) к величине объединенного среднеквадратического отклонения (w)
| Вид рода Sorex | Центроидный размер, CS ± SE (n) | isodon | araneus | caecutiens | minutus |
|---|---|---|---|---|---|
| isodon | 17.39 ± 0.15 (28) | – | 0.18* | 2.62 | 5.18 |
| araneus | 17.21 ± 0.09 (50) | 1.01* | – | 2.63 | 5.65 |
| caecutiens | 14.01 ± 0.15 (45) | 1.24 | 1.23 | – | 1.20* |
| minutus | 12.64 ± 0.12 (16) | 1.38 | 1.36 | 1.11* | – |
При оценке соотношений средних величин CS между равнозубой и обыкновенной бурозубками величина отношения Хатчинсона составила 1.01, а для средней/малой – 1.11, что отражает высокую вероятность конкуренции между этими парами видов (см. табл. 1). Однако в случае сравнения крупных и мелких по размерам бурозубок отношения Хатчинсона следовали его правилу (см. табл. 1). Следовательно, правило Хатчинсона подтвердилось не во всех случаях, а только между наиболее крупными и мелкими землеройками.
В известной модели Макартура и Левинса [10] степень разделения ниш двух видов (ρ) по величине отношения разности средних значений (d) общего ресурса к обобщенной величине среднеквадратического отклонения (w) должна быть ρ = d/w > 1 для поддержания возможности их сосуществования. Минимально допустимое отношение d/w между видами соответствует величине отношения лимитирующего сходства ρ = 1.56, при которой виды теоретически еще способны сосуществовать. Если d/w < 1, предполагается конкуренция, а при d/w > 3, исходя из “правила Хатчинсона”, конкуренция будет маловероятна.
Из табл. 1 следует, что не только правило Хатчинсона, но и отношение сходства ниш (ρ) не соблюдается по CS и их изменчивости между двумя парами наиболее крупных (S. isodon и S. araneus) и наиболее мелких видов (S. caecutiens и S. minutus), что косвенно указывает на вероятность конкуренции для каждой пары видов. Однако оба условия хорошо соблюдаются между крупными и мелкими видами (см. табл. 1).
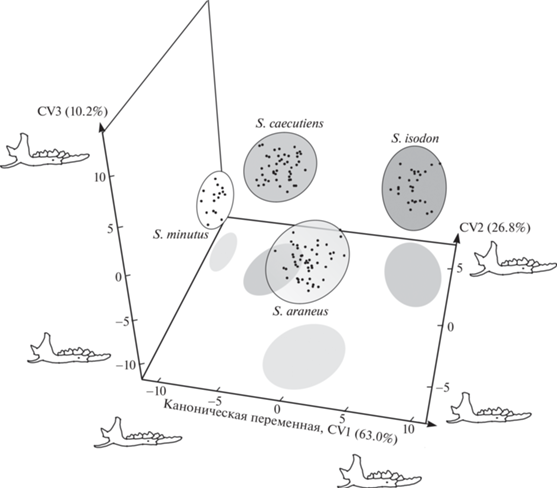
Результаты канонического анализа представлены на рис. 2. Из рисунка следует, что все межвидовые различия по форме нижней челюсти велики и упорядочены вдоль первой канонической переменной (CV1): от эллипсоида рассеивания ординат самой мелкой S. minutus, локализованного в области отрицательных значений оси, до эллипсоидов самых крупных видов – S. araneus и S. isodon. Выборка S. araneus наиболее смещена вдоль второй канонической оси (CV2), уклоняясь от S. isodon и других видов. В область положительных значений вдоль CV3 от всех остальных бурозубок уклонилась выборка S. caecutiens.
Рис. 2.
Результаты канонического анализа прокрустовых координат мандибул четырех симпатрических видов таксоцена землероек рода Sorex Висимского заповедника в морфопространстве канонических переменных (CV1–CV3).
Сравнение обобщенных дистанций Махаланобиса между выборками видов землероек (табл. 2) позволяет заключить, что сходные по размерам мандибулы S. isodon и S. araneus существенно удалены друг от друга по ее форме (D = 10.26; p < 0.0001). Несколько сближены по форме нижней челюсти S. araneus и S. caecutiens (D = 7.49; p < 0.0001), но и они при этом значимо различаются по размерам, не нарушая правила Хатчинсона. Таким образом, сходство видов по размеру мандибул компенсируется различиями в их конфигурации, причем, выборки всех видов существенно разобщены в морфопространстве, отражая видовую специфику формы нижней челюсти, связанную с трофической специализацией.
Таблица 2.
Обобщенные расстояния Махаланобиса (D) по прокрустовым координатам, характеризующим изменчивость формы нижней челюсти, между симпатрическими видами землероек рода Sorex Висимского заповедника (уровни значимости (p) расстояний представлены в верхней треугольной матрице)
| Вид | S. isodon | S. araneus | S. caecutiens | S. minutus |
|---|---|---|---|---|
| S. isodon | 0 | p < 0.0001 | p < 0.0001 | p < 0.0001 |
| S. araneus | 10.26 | 0 | p < 0.0001 | p < 0.0001 |
| S. caecutiens | 11.30 | 7.49 | 0 | p < 0.0001 |
| S. minutus | 17.29 | 13.06 | 9.24 | 0 |
Эллипсоиды изменчивости формы мандибул в выборках землероек характеризуют 95% дисперсии ординат особей. Вдоль каждой канонической переменной указаны доли межгрупповой изменчивости (%) и представлены контурные изображения (outlines) мандибул, соответствующие максимальным и минимальным значениям осей.
Поскольку различия формы мандибул должны сопровождаться и различиями в функции костно-мышечного аппарата нижней челюсти, оценили морфофункциональные мандибулярные индексы видов (табл. 3). Межвидовые различия по значениям мандибулярных индексов были велики и в большинстве случаев значимы. Между сходными по размерам мандибул S. isodon и S. araneus существенные различия выявлены по двум индексам (TI и TM), т.е. виды значимо различаются по продольным усилиям при разрезании добычи. Однако они не различаются по способности к грызению (индекс MM) и горизонтальному качанию левой и правой ветвей нижней челюсти (swinging, по терминологии Зажигина и Войта [11]) при перехватывании пойманной добычи – индекс АМ. Последнее связано с особым способом захвата добычи во время охоты и хорошо выражено у более мелких видов бурозубок – средней и малой (см. табл. 3). Интересно, что крупная S. isodon сходна с наименьшей по размерам S. minutus по величине массетерно-резцового индекса MI (t = 1.4; p > 0.05), т.е. эти виды слабо используют резцы при грызении, в отличие от S. caecutiens и S. araneus.
Таблица 3.
Сравнение средних значений мандибулярных индексов (с учетом стандартных ошибок ±SE) у четырех симпатрических видов землероек рода Sorex Висимского заповедника на Среднем Урале
| Мандибулярный индекс | Виды землероек рода Sorex | |||
|---|---|---|---|---|
| isodon (n = 28) |
araneus (n = 50) | caecutiens (n = 45) |
minutus (n = 16) |
|
| TI | 0.328 ± 0.002 | 0.312 ± 0.001 | 0.305 ± 0.001 | 0.296 ± 0.002 |
| TM | 0.565 ± 0.003 | 0.538 ± 0.002 | 0.524 ± 0.002 | 0.498 ± 0.004 |
| MI | 0.159 ± 0.001 | 0.162 ± 0.001 | 0.165 ± 0.001 | 0.157 ± 0.001 |
| MM | 0.274 ± 0.002 | 0.279 ± 0.001 | 0.284 ± 0.002 | 0.264 ± 0.002 |
| AM | 0.894 ± 0.011 | 0.882 ± 0.007 | 0.995 ± 0.008 | 1.013 ± 0.014 |
Таким образом, различия по форме нижней челюсти и мандибулярным индексам позволяют объяснить возможность избегать конкуренции сходным по размерам парам крупных и мелких бурозубок – соответственно, равнозубой и обыкновенной, с одной стороны, и средней и малой, с другой, за счет их трофический специализации на основе преобразования костно-мышечной структуры мандибул. Полученные данные косвенно указывают на специфику в предпочитаемой диете землероек, как по размерам добычи, так и по способности ее захвата во время охоты, что тоже позволяет избежать конкуренции. Снижению конкуренции S. isodon и S. araneus способствует избирательность биотопов [12].
Таким образом, землеройки способны избегать конкуренции и при нарушении правила Хатчинсона, причем не столько благодаря размерам, сколько за счет дифференциации по форме нижней челюсти, ее функциональной специализации, способов захвата добычи, размерного и механического состава диеты. Следует заключить, что экологическое правило Хатчинсона не является обязательным универсальным условием, а лишь одним из условий для уменьшения конкуренции между близкими видами, поскольку не только размеры, но главным образом структурно-функциональные особенности видов обеспечивают возможность их успешного синтопного сосуществования.
Список литературы
Hutchinson G.E. Homage to Santa Rosalia, or why are there so many kinds of animals? // Amer. Nat. 1959. V. 93. P. 145–159.
Hutchinson G.E. Treatise on Limnology. Vol. II. Introduction to Lake Biology and the Limnoplankton. N.Y.: John Wiley and Sons. 1967. P. 1115.
Lewin R. Santa Rosalia was a goat // Science. 1983. V. 221. P. 636–639.
Grant P.R., Grant B.R. Evolution of character displacement in Darwin 's finches // Science. 2006. V. 313. P. 224–226.
Adams D.C. Parallel evolution of character displacement driven by competitive selection in terrestrial salamanders // BMC Evol. Biol. 2010. V. 10 (72). P. 1–10.
Rohlf F.J., Slice D. Extension of the Procrustes method for the optimal superimposition of landmarks // Syst. Zool. 1990. V. 39. № 1. P. 40–59.
Zelditch M.L., Swiderski D.L., Sheets H.D., et al. Geometric Morphometrics for Biologists: A Primer. New York: Elsevier Acad. Press. 2004. P. 437.
Anderson Ph.S.L., Renaud S., Rayfield E.J. Adaptive plasticity in the mouse mandible // BMC Evolutionary Biology. 2014. V. 14. P. 85–93.
Васильев А.Г., Васильева И.А., Коурова Т.П., и др. Форпостная популяция белобрюхой белозубки на северной границе ареала в Оренбургской области // Фауна Урала и Сибири. 2022. № 2. С. 87–108.
MacArthur R., Levins R. The limiting similarity, convergence, and divergence of coexisting species // Amer. Nat. 1967. V. 101. № 921. P. 377–385.
Zazhigin V.S., Voyta L.L. Northern Asian Pliocene–Pleistocene beremendiin shrews (Mammalia, Lipotyphla, Soricidae): a description of material from Russia (Siberia), Kazakhstan, and Mongolia and the paleobiology of Beremendia // Journal of Paleontology. 2019. V. 93. № 6. P. 1234–1257.
Шварц Е.А., Демин Д.В., Замолодчиков Д.Г. Экология сообществ мелких млекопитающих лесов умеренного пояса (на примере Валдайской возвышенности). М.: Наука. 1992. С. 127.
Дополнительные материалы отсутствуют.
Инструменты
Доклады Российской академии наук. Науки о жизни