

Доклады Российской академии наук. Науки о жизни, 2023, T. 513, № 1, стр. 533-538
Среднеголоценовый бурый медведь (Ursus arctos) с острова Большой Ляховский (Новосибирские Острова)
М. Ю. Чепрасов 1, *, Г. Г. Боескоров 1, 2, Г. П. Новгородов 1, А. Н. Тихонов 1, 3, Л. В. Григорьева 1, Е. С. Булыгина 4, Н. В. Слободова 4, 5, Ф. С. Шарко 4, 6, А. В. Протопопов 7, А. В. Недолужко 8
1 Федеральное государственное автономное образовательное учреждение высшего образования “Северо-Восточный федеральный университет
имени М.К. Аммосова”
Якутск, Россия
2 Федеральное государственное бюджетное учреждение науки “Институт геологии алмаза и благородных металлов Сибирского отделения Российской академии наук”
Якутск, Россия
3 Федеральное государственное бюджетное учреждение науки “Зоологический институт Российской академии наук”
Санкт-Петербург, Россия
4 Национальный исследовательский центр “Курчатовский институт”
Москва, Россия
5 Национальный исследовательский университет “Высшая школа экономики”
Москва, Россия
6 Федерального государственного учреждения “Федеральный исследовательский центр Фундаментальные основы биотехнологии
Российской академии наук”
Москва, Россия
7 Академия наук Республики Саха
Якутск, Россия
8 Автономная некоммерческая образовательная организация высшего образования
“Европейский университет в Санкт-Петербурге”
Санкт-Петербург, Россия
* E-mail: nohsho@mail.ru
Поступила в редакцию 10.07.2023
После доработки 26.07.2023
Принята к публикации 28.07.2023
- EDN: XQKHRW
- DOI: 10.31857/S2686738923600498
Аннотация
Приводится морфологическое описание уникальной находки субфоссильного бурого медведя (Ursus arctos L., 1758) – его замороженной мумии, найденной впервые в истории. Находка представляет из себя хорошо сохранившуюся тушу зверя возрастом около 3.5 тысяч лет. Обсуждаются результаты исследования медведя методами компьютерной томографии и анализа ДНК.
Плейстоценовые остатки U. arctos, представленные костями скелета, достаточно редки и встречаются на территории Якутии с начала среднего неоплейстоцена [1, 2].
Первая находка замороженной мумии древнего бурого медведя была обнаружена в августе 2020 г. на острове Большой Ляховский (Новосибирский архипелаг) в 8 км северней устья р. Большой Этерикан, от которой медведь получил название “Этериканский”.
Вытаивающую из мерзлоты тушу медведя обнаружили оленеводы общины “Муостаах” на склоне обрыва моря Лаптевых. В марте 2021 г. туша медведя была привезена в г. Якутск в Музей мамонта, где хранится в морозильной камере при t = –20°C под инвентарным номером MM-F54.
Ветеринарные исследования не выявили возбудителей сибирской язвы, трихинеллеза и других особо опасных заболеваний.
Радиоуглеродное датирование, проведенное по лучевой кости правой конечности в Carbon Analysis Lab Co., Ltd. (Южная Корея), показало, что возраст находки относится к среднему голоцену (3460 ± 40 л.н. (OTg220171), не калиброванная дата) [3].
Компьютерная томография ископаемого U. arctos была проведена 20.04.2022 г. в диагностическом центре “RadX-Med” (г. Якутск) с помощью томографа “Toshiba Aquilion 16”. Измерения черепа и нижней челюсти выполнены по методике Г.А. Новикова [4] с помощью программного обеспечения “Vidar Dicom Viewerс 3.0”.
Туша хорошо сохранилась (рис. 1) и незначительно мумифицирована. Спина деформирована (уплощена), по-видимому, из-за травмы, приведшей к гибели особи (см. ниже), а также вследствие длительной транспортировки мумии с острова Б. Ляховский при положительных температурах воздуха.
Рис. 1.
Замороженная туша Этериканского медведя. Вверху – вид сбоку, в середине – вид сверху, внизу – вид снизу.

Особенности телосложения и пропорции тела характерны для бурого медведя: голова большая и тяжелая с широким и относительно высоким лбом, умеренно вытянутая; конец носа крупный, черного цвета; губы большие, способные к вытягиванию. Холка высокая, передние конечности относительно длинные, мощные. Задняя часть тела (круп и бедра) массивнее передней. Подошвы кистей и стоп лишены волос (рис. 1, 2). По наружным половым органам определено, что медведь был самкой.
Рис. 2.
Части тела Этериканского медведя. Слева – голова, вид сбоку. В середине и справа – подошвы передней и задней лап.
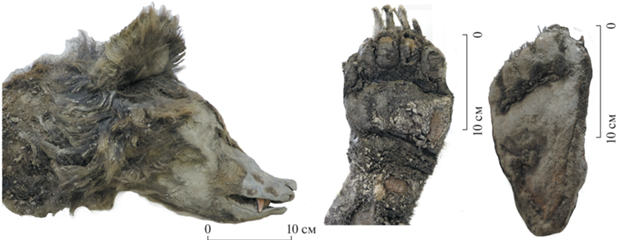
Шерсть имеет желтовато-светлокоричневый цвет (верх головы, плечи), бурый, темно-бурый и почти черный цвет (шея, бедра, пах). На большей части спины она выпала. На спине и плечах сохранились участки двуцветных волос: нижняя их часть светло-коричневая, верхняя – бурая. Нижние части лап темнее спины, покрыты короткими темно-бурыми, почти черными волосами. Длина волос: 8–8.2 см (брюхо), 10–12 см (шея), 10–11 см (плечи, бедра), 11–13.5 см (пах), 11–11.5 см (бока).
Мех плотный с длинными остевыми волосами до 12–15 см и с густым подшерстком характерен для бурого медведя в осенне-зимний период [1, 5]. Высокая упитанность медведя и большое количество абдоминального жира (толщиной до 4–5 см), указывают на то, что он был готов залечь в спячку или только что пробудился [6].
По совокупности признаков (степень зарастания венечного и носового швов [7, 8], степень стертости зубов [8], ширина пальмарной мозоли (табл. 1)) можно констатировать, что Этериканская медведица была половозрелой особью возрастом около 5 лет.
Таблица 1.
Размеры (в см) и масса тела (в кг) взрослых самок бурых медведей из Восточной России
| Размеры и масса | Современные | Субфо-ссильный | ||||||
|---|---|---|---|---|---|---|---|---|
| Якутия, Верхоянье [9], n = 3 | Центральная Якутия, n = 4* | Северо-Восток Азии ** | Якутия, о. Б. Ляховский | |||||
| Limit (пределы вариаций) | M (средняя) | Limit | M ± m*** | Limit | M ± m | n | ||
| Высота в холке | 73–84 | 77 | 73–88 | 80.5 ± 3.23 | 82–111 | 96.2 ± 2.51 | 11 | 75 |
| Длина тела | 135–145 | 140 | 160–172 | 160 ± 2.55 | 160–197 | 172.4 ± 3.96 | 11 | 155–160 |
| Обхват груди за лопатками | 63–71 | 68 | 89–110 | 98.8 ± 4.4 | 65–144 | 113.0 ± 9.08 | 7 | 114 |
| Длина уха | 13–13 | 13 | 9–11 | 9.8 ± 0.75 | 10–14 | 11.8 ± 0.40 | 10 | 10.5 |
| Ширина передней мозоли | 11–11.5 | 11.2 | 12–13 | 12.4 ± 0.24 | 11.5–15.5 | 13.9 ± 0.36 | 11 | 11.8 |
| Длина ступни | 22–23 | 22 | – | – | – | – | – | 22.4 |
| Масса тела | 56–75 | 65 | – | – | 105–170 | 134.9 ± 8.33 | 8 | 77.6 |
* неопубл. данные Боескорова Г.Г.; **вычисления Боескорова Г.Г. по: [6]; ***m – ошибка средней.
По ряду промеров тела медведь с Б. Ляховского острова превышает параметры самок современного бурого медведя из Верхоянья и соответствует таковым более крупной формы медведя из центральной Якутии (табл. 1). Самки U. arctos из Северо-Востока Азии (Чукотка, бассейн р. Омолон, северное Приохотье) [6] более крупные, чем медведи из Якутии (табл. 1).
По большинству промеров черепа исследуемый U. arctos соответствует средним размерам черепов самок современного вида с территории Якутии, измеренных нами, и по ряду промеров несколько превосходят черепа медведиц из Верхоянья и южной Якутии [1] (табл. 2). Бурый медведь из Северо-Востока Азии в среднем крупнее современных и субфоссильного медведя из Якутии (табл. 2).
Таблица 2.
Размеры черепа (мм) взрослых самок бурых медведей из Восточной России
| Промеры | Современные | Субфос-сильный | ||||||
|---|---|---|---|---|---|---|---|---|
| Якутия | Северо-Восток Азии | Якутия | ||||||
| [9]; n = 5 | наши данные | [6]; n = 7 | о. Б. Ляховский | |||||
| Limit | M±m | Limit | M±m | n | Limit | M±m | ||
| Кондилобазальная длина | 256–278 | 268.0 ± 3.69 | 263–302 | 286.0 ± 4.42 | 7 | 271.5–332 | 307.7 ± 8.7 | 294.9 |
| Общая длина | 272–296 | 285.0 ± 1.48 | 274–311 | 298.3 ± 3.79 | 9 | – | – | 297.4 |
| Длина твердого неба | 139–149 | 145.0 ± 1.94 | 142–154.6 | 151.5 ± 2.0 | 6 | 142–176.1 | 160.4 ± 4.5 | 147.8 |
| Переднезадний диаметр клыка у основания | – | – | 15.6–19 | 17.0 ± 0.38 | 9 | – | – | 17.3 |
| Длина верхнего ряда щечных зубов Р4 – М2 | 66–68 | 67.3 ± 0.16 | 62–79 | 70.06 ± 1.57 | 9 | – | – | 69.4 |
| Скуловая ширина | 167–183 | 176.0 ± 2.86 | 168–198 | 177.0 ± 4.19 | 7 | 117.3–220.0 | 190.0 ± 14.2 | 182 |
| Высота затылка | 72–87 | 82.6 ± 1.89 | 72.5–83 | 79.74 ± 4.62 | 5 | – | – | 78.4 |
| Ширина в надглазничных отростках | 92–94 | 93.8 ± 0.37 | 89.4–106 | 96.3 ± 2.57 | 7 | – | – | 100.6 |
Некрупные размеры Этериканского медведя, схожие с таковыми современного U. arctos Якутии, могут свидетельствовать о том, что своеобразная экоморфа якутского медведя, обитающая в суровых условиях со скудным низкокалорийным рационом, сформировалась к середине голоцена.
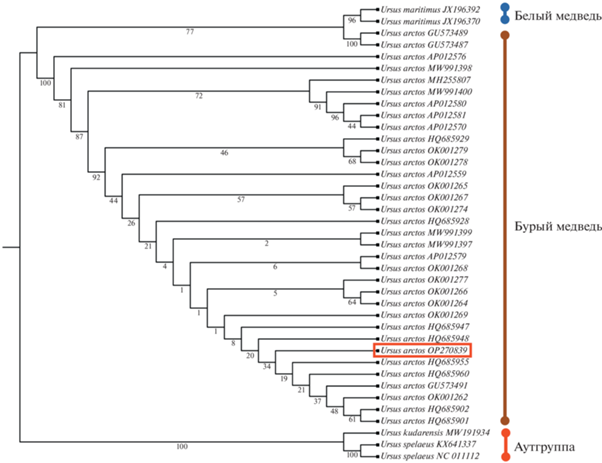
Молекулярно-генетическое исследование мягких тканей Этериканского медведя проводили в НИЦ “Курчатовский институт” с соблюдением всех необходимых для подобных экспериментов условий. Из мышечной ткани была выделена ДНК и приготовлены ДНК-библиотеки, как было описано ранее [10]. ДНК-библиотеки были отсеквенированы на приборе Illumina Novaseq6000. Полученные данные секвенирования прошли стандартные процедуры очистки, используя программный конвейер PALEOMIX 1.2.14 [11]. Последовательность митогенома была получена с применением программного обеспечения SPAdes v3.15.3 [12]. Для видовой идентификации были отобраны последовательности гена цитохрома б (CYTB) и контрольной области митохондриальной ДНК (D-петля). Филогенетическая реконструкция с использованием RaxML v8.2.12. [13], показала, что Этериканский бурый медведь (OP270839) группируется вместе с современными бурыми медведями (рис. 3).
Рис. 3.
Реконструкция филогенетического дерева видов рода Ursus построенная на основании последовательностей гена цитохрома б (CYTB) и контрольной области митохондриальной ДНК (D-петля) методом максимального правдоподобия (ML). Бутстреп значения указаны под ветвями филогенетического древа. Номера NCBI представителей рода Ursus указаны на окончаниях ветвей древа. Образец Этериканского медведя отмечен красным прямоугольником. В качестве аутгруппы выбраны образцы пещерного медведя (U. spelaeus) и кударского пещерного медведя (U. kudarensis).
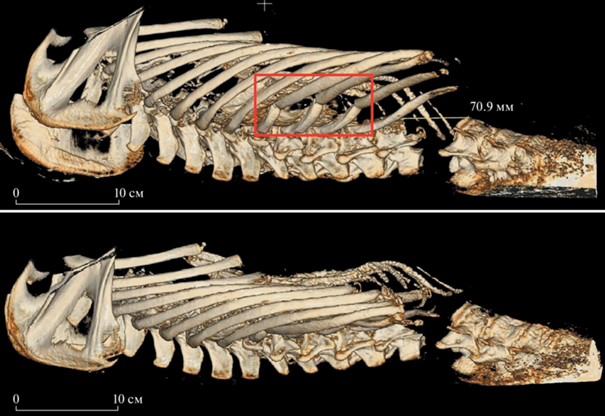
Причиной гибели медведя стало механическое повреждение позвоночного столба, а также закрытая торакальная травма. В результате повреждения позвоночника произошло его отделение на уровне первого и второго поясничных позвонков (травматическая ампутация), что, по-видимому, привело к разрыву спинного мозга и параличу нижнего отдела тела. Закрытая травма грудной клетки сопровождалась компрессионной механической асфиксией (сдавливание груди и живота твердым предметом) с ее смещением вправо и переломом 10-го-12-го ребер (рис. 4). По-видимому, медведь был придавлен свисавшей глыбой монолита многолетней мерзлоты.
Рис. 4.
Механические повреждения костей скелета Этериканского медведя. На верхнем рисунке (латерально-дорсальный вид) красным квадратом отмечены места переломов ребер. На нижнем рисунке (латеральный вид) показана сдавленная грудная клетка.
Место обнаружения мумии медведя расположено в 250–300 км от современной северной границы распространения U. arctos, которая в Якутии ограничивается лесотундрой, в зону тундры он заходит очень редко и только в летнее время [9].
Пока неясна причина обнаружения бурого медведя на острове Б. Ляховский: был ли это необычно дальний заход от основного места обитания, каким образом он попал на остров – вплавь через пролив Дм. Лаптева или по льду в весеннее время? Мог ли этот медведь найти в голоцене приемлемые условия для обитания на острове? Эти вопросы нам предстоит выяснить в процессе дальнейшего изучения Этериканского медведя.
Список литературы
Боескоров Г.Г., Барышников Г.Ф. Позднечетвертичные хищные млекопитающие Якутии. СПб.: Наука, 2013. 199 с.
Boeskorov G.G., Baryshnikov G.F., Tikhonov A.N. et al. New Data on Large Brown Bear (Ursus arctos L., 1758, Ursidae, Carnivora, Mammalia) from the Pleistocene in Yakutia. // Dokl. Earth Sc. 486, 617–622 (2019).
Boulygina E., Sharko F., Cheprasov M. et al. Ancient DNA Reveals Maternal Philopatry of the Northeast Eurasian Brown Bear (Ursus arctos) Population during the Holocene. // Genes. 2022; 13(11):1961.
Новиков Г.А. Хищные млекопитающие фауны СССР. М.; Л.: АН СССР, 1956. 294 с.
Гептнер В.Г., Наумов Н.П., Юргенсон П.Б. и др. Морские коровы и хищные. Т. 2. Млекопитающие Советского Союза. М.: Высш. школа, 1967. 1004 с.
Чернявский Ф.Б., Кречмар М.А. Бурый медведь (Ursus arctos L.) на Северо-Востоке Сибири. Магадан: ИБПС СВНЦ ДВО РАН, 2001. 93 с.
Клевезаль Г.А. Принципы и методы определения возраста млекопитающих. М.: Тов. научн. изд. КМК, 2007. 283 с.
Гуськов В.Ю. Методика определения возраста бурого медведя (Ursus arctos Linnaeus, 1758) по черепу // Успехи наук о жизни, 2014. № 9. С. 100–106.
Тавровский В.А., Егоров О.В., Кривошеев В.Г. и др. Млекопитающие Якутии. М.: Наука, 1971. 659 с.
Boulygina E., Tsygankova S., Sharko F., et al. // Journal of Archaeological Science: Reports. 2020. V. 31. P. 102357.
Schubert M., Ermini L., Der Sarkissian C. et al. // Nature Protocols. 2014. V. 9. P. 1056–1082.
Bankevich A., Nurk S., Antipov D. et al. // Journal of Computational Biology. 2012. V. 19. P. 455–477.
Stamatakis A., Hoover P., Rougemont J. // Systematic Biology. 2008. V. 57. P. 758–771.
Дополнительные материалы отсутствуют.
Инструменты
Доклады Российской академии наук. Науки о жизни