

Экология, 2019, № 3, стр. 163-171
Влияние внутригодового распределения характеристик климата на разнообразие сосудистых растений в Среднем Поволжье
П. А. Шарый a, *, А. В. Иванова b, Л. С. Шарая b, Н. В. Костина b
a Институт физико-химических и биологических проблем почвоведения РАН
142290 Московская обл., Пущино, ул. Институтская, 2, Россия
b Институт экологии Волжского бассейна РАН
445003 Самарская обл, Тольятти, ул. Комзина, 10, Россия
* E-mail: p_shary@mail.ru
Поступила в редакцию 03.05.2018
После доработки 13.06.2018
Принята к публикации 20.06.2018
Аннотация
Изучена связь видового богатства сосудистых растений на площадках размером 10 × 10 км с климатическими показателями в Среднем Поволжье. Выявлены наиболее значимые показатели – сумма осадков и температура марта, температура октября и осадки зимы, которые при совместном действии обусловливают 74% варьирования видового богатства. В условиях относительно низких климатических градиентов для анализа использована специальная функция суммы осадков марта, описывающая выраженную в виде пика нелинейную зависимость богатства сосудистых растений от нее. Дана экологическая интерпретация связей, построена карта богатства видов. Обсуждается гипотеза о том, что на территории с небольшими градиентами климата малые перепады внутригодовых показателей температуры и осадков могут являться критическими факторами для изменения числа видов сосудистых растений в пространстве.
Изучение зависимости разнообразия растительности от факторов окружающей среды составляет одну из труднейших, но и важнейших задач экологии [1]. При глобальном рассмотрении разнообразие, за некоторыми исключениями, увеличивается от полюсов к экватору, что является одной из наиболее известных экологических закономерностей [2]. Однако при рассмотрении подобных закономерностей модели стратегии отбора видов [3, 4] заметно упрощены, так как данному набору стратегий могут отвечать различные условия окружающей среды [5]. Для формализации предположения о том, что число видов в данном типе местообитания связано с общим числом видов региона, введено понятие регионального пула видов [5]. Пул видов зависит от истории развития региона и эволюции растительности. Эти процессы трудно оценить, однако пространственное распределение разнообразия растений в регионе зависит от факторов окружающей среды, следовательно, его описание и моделирование намного проще. Поскольку видовой пул увеличивается с площадью региона, размеры площадок наблюдения часто фиксируют. Число видов на площадках фиксированного размера есть плотность видов [1, 5, 6], нередко называемая также богатством или числом видов.
В противоположность тропическим лесам, где основная часть видов находится в древесном ярусе, большинство видов растений в умеренных лесах Европы и США встречается в травяном ярусе [7, 8]. В умеренной полосе Европы богатство видов травянистой растительности лучше отражает вариацию ресурсов в пространстве, в то время как деревья – за бóльшее время и для бóльших площадей [8].
Основой для сравнения богатства видов с факторами окружающей среды являются модели множественной регрессии [4, 7–10]. Применяют также модели эффекта средней области MDE (mid-domain effect), в которых рассматривается случайное помещение видов в область, границы которой эти виды не могут пересекать [11]. Однако на примере богатства видов папоротников показано, что роль MDE невелика, как и подходов со многими моделями, различие между которыми было незначительно [12]. Для исследований также используют генерализованные аддитивные модели множественной регрессии или непараметрические модели “случайного леса” [13].
Причины зависимости богатства видов от условий среды изучали многие авторы. В работе [14] приведено 120 гипотез об этом, причем автор замечает, что приведенный им список гипотез далеко не полон. Большое число гипотез, а также разнообразие факторов среды, влияющих на богатство видов, свидетельствуют о том, что причины пространственной изменчивости богатства видов сложны и пока недостаточно хорошо поняты [15, 16].
Предположение о важной роли энергии солнечной радиации для разнообразия, высказанное в работе [6], было поддержано в ряде исследований [17–19]. В локальном масштабе показано [10], что распределение солнечной энергии по склонам с учетом сомкнутости крон деревьев было самым влиятельным фактором среды для разнообразия травянистых растений. Среди ведущих факторов пространственного распределения богатства видов называют площадь, климат и энергию, историю региона, региональный пул видов, свойства почв и фрагментацию лесов.
Особый интерес вызывают пики на кривых зависимости таксономического разнообразия от географической широты или высоты над уровнем моря. Одна из причин возникновения пиков такова: для богатства видов трав в умеренных лесах Европы пики не наблюдались из-за того, что сухой вес надземной фитомассы трав в лесах редко превышает 300 г/м2, что недостаточно для конкурентного исключения видов, в то время как для лугов широтный пик достигается при фитомассе 450–600 г/м2 [8]. При глобальном изучении связи разнообразия папоротников с высотой в горах обнаружены резкие пики, обусловленные особым характером изменения увлажнения с высотой [12].
Результаты регионального изучения связи богатства видов сосудистой растительности с факторами среды зависят от масштаба, т.е. от размера изучаемых площадок [20–22]. При региональных исследованиях изменения климата в пространстве, как правило, сравнительно малы, и связи между богатством видов и климатическими факторами могут оказываться не столь тесными, как при глобальных исследованиях [2]. При использовании климатической базы данных WorldClim высокого разрешения (900 м) [23] наиболее популярны в исследованиях среднегодовые температура и осадки, средняя температура наиболее теплого времени года, средние осадки наиболее влажного времени года и стандартные отклонения этих переменных [13]. Используют также годовую реальную и потенциальную эвапотранспирации [6, 13, 24]. Однако в этих условиях может стать ведущей роль климата отдельных месяцев, что изучено пока недостаточно. О.В. Морозовой [2] высказано предположение, что вариации факторов среды внутри площадок размером 10 × 10 км могут играть важную роль в этой ситуации; таких исследований на количественном уровне, однако, пока очень мало.
Следует отметить, что во многих региональных исследованиях чаще всего учитывали среднегодовые климатические факторы [6, 12, 24; и др.], реже – влияние климата отдельных месяцев [9] или времен года [13], при этом нелинейный вид связей специально не рассматривался. Наша гипотеза состоит в том, что видовое разнообразие сосудистых растений в условиях относительно мягкого климата и слабых климатических градиентов в первую очередь определяется влиянием внутригодовых вариаций температур и осадков, при этом особо важным может быть вид нелинейной функции в связи. Цель данной работы – проверка высказанной гипотезы на примере регионального изучения богатства видов сосудистых растений в Среднем Поволжье.
ОБЪЕКТЫ И МЕТОДЫ
Число видов сосудистых растений подсчитывали на 25 площадках размером 10 × 10 км в Среднем Поволжье (Самарская и Ульяновская области) на территории размером 282 × 285 км. На юго-востоке этого региона располагаются степи, севернее – лесостепная зона и местами леса (от сосновых и сосново-широколиственных до дубрав). Покрытие лесами в регионе низкое. Почвы меняются от черноземов на юге до дерново-подзолистых на севере. Абсолютная высота на площадках наблюдения колеблется от 38 до 239 м при средней 117 ± 58 м (здесь и ниже разброс между точками наблюдения описан одним стандартным отклонением).
Для изучаемого региона характерны три типа почв: черноземы, серые лесные и дерново-подзолистые; их учет проводился по почвенной карте масштаба 1 : 2.5 млн. Если площадке отвечало два типа почв, то брали тип почвы по преобладанию, т.е. площадь которого больше.
Флористические описания проводили в 2004–2017 гг. [25–29; и др.]. За этот период каждую из площадок посещали несколько раз, включая сезонные повторы (с мая по сентябрь). Кроме того, были использованы и литературные данные.
Для выявления наиболее полного видового состава высших сосудистых растений в пределах указанной площадки (10 × 10 км) в различные сезоны вегетации закладывали от 3 до 6 временных лагерей, от которых прокладывали серию маршрутов длиной до 5–7 км с охватом максимального количества экотопов. Многие маршруты посещали несколько раз в разные годы и разные сезоны года, что обеспечивало выявление общей картины флористического разнообразия, которая, мы полагаем, на сегодняшний день близка к описанию полного разнообразия, являющегося неким идеалом, недостижимым на практике. Положение некоторых площадок и лагерей в них для части Самарской области показано на карте в работе [30]. Для целей настоящей работы отобрано 25 таких площадок с наиболее полным описанием флоры в них. Включали также и адвентивные виды, которые на староосвоенной территории успели распространиться и не сосредоточены в местах начала их заселения.
Богатство видов на площадках составило от 395 до 690 при среднем 511 ± 80. В табл. 1 приведены некоторые характеристики точек наблюдения, показанных на рис. 3. Средние многолетние данные месячных осадков и температур, температур и сумм осадков по сезонам, а также значения 19 биоклиматических переменных, например среднегодовая сумма осадков и ее коэффициент вариации, взяты из базы данных WorldClim [23]. В WorldClim месячные климатические переменные усреднены за 50 лет (1950–2000 гг.) и представлены с пространственным разрешением 900 м.
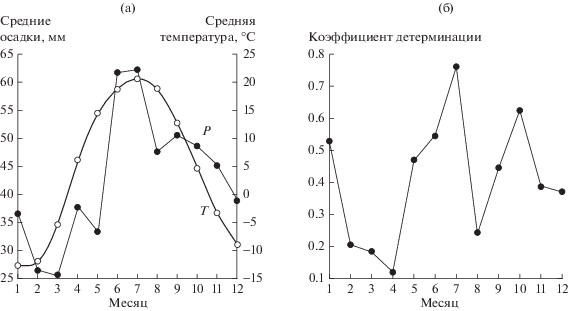
Изменение по месяцам усредненных по 25 площадкам значений температур и осадков по данным WorldClim показано на рис. 1a. Среднегодовая температура воздуха в регионе составляет 4.5 ± 0.5°С, а среднегодовая сумма осадков – 514 ± 30 мм.
Рис. 1.
Внутригодовое распределение среднемесячных осадков P и температур T (а) и распределение тесноты связи между месячными осадками и температурами воздуха (б).
Для установления связей между количеством видов на обследованных площадках и характеристиками климата использовалась множественная регрессия. Помимо этого, рассматривали значения абсолютной высоты и типы почв.
Статистические методы. Базовые предположения множественной регрессии есть (1) нормальность распределения остатков (ошибок), (2) постоянство дисперсии, (3) отсутствие значимой линейной связи между предикторами и (4) отсутствие значимой автокорреляции остатков. Предположение (1) проверяли по графику нормальной вероятности, (2) – с помощью графика зависимости остатков от предсказанной зависимой переменной [31] и (3) – по критерию того, что максимум факторов увеличения дисперсии должен быть меньше 5.15 [32]. Любые комбинации предикторов, для которых критерий (3) нарушался, исключали из рассмотрения. Автокорреляция остатков быстро падает с расстоянием между точками наблюдения, становясь для растительности незначимой при расстоянии 250 м [33]. В нашем случае это расстояние превышало 10 км, и автокорреляция не учитывалась.
Выбор наиболее значимых предикторов осуществляли путем перебора всех комбинаций из четырех предикторов, для которых выполнялся критерий (3); отбирали ту комбинацию предикторов, для которой найден наибольший коэффициент детерминации [31]. Пятый предиктор оказывается обычно уже незначимым в модели. Таким образом, выбор предикторов в полученной модели не зависел от предпочтений авторов.
Нами были использованы центрированные квадраты климатических факторов, т.е. для фактора X его квадрат X 2 заменяли на (X – XAV)2, где XAV – среднее по выборке. Это уменьшает линейную связь между предикторами [31] и особенно важно, когда отклонение X от среднего XAV имеет существенное значение в понимании происходящих явлений.
Для верификации моделей применяли эмпирический критерий Degr < 50% [32], где показатель деградации модели в новых точках наблюдения есть Degr = 100(R2/$R_{{\Pr }}^{2}$ – 1), а $R_{{\Pr }}^{2}$ – “предсказательный” коэффициент детерминации, рассчитанный по методике кросс-валидации Аллена [31]. Анализ модели и построение карты считались обоснованными только при выполнении этого критерия.
Оценку значимости предиктора в модели и установление относительной значимости предикторов проводили с помощью t-статистик Стьюдента [31], подписанных в уравнениях регрессии нижним индексом. Если модуль t-статистики был меньше рассчитанного по статистически таблицам значения 2.086, то предиктор в модели рассматривался как статистически незначимый.
Для проведения расчетов и построения карт использовалась разработанная П.А. Шарым программа “Аналитическая ГИС Эко” версии 1.08r.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Распределение тесноты связи среднемесячных температур и осадков по месяцам показано на рис. 1б. Для каждого месяца использовали 25 точек (площадок) наблюдения, по которым рассчитывали коэффициент детерминации для этого месяца по данным WorldClim.
Наиболее тесная связь осадков с температурами наблюдалась для июля (R2 = 0.759, P < 10–6), а наименее – для апреля (R2 = 0.119, связь незначима). Для февраля, марта, апреля и августа связи между осадками и температурами были практически незначимыми (P > 0.01). При этом связь была отрицательна для любого месяца. Отметим, что в холодное время года тесная отрицательная связь между температурой и осадками может позитивно влиять на видовое разнообразие растений, так как низким температурам чаще соответствуют большие осадки, препятствующие потере тепла почвами. В марте, однако, температура воздуха и осадки практически не связаны, что может обусловливать особую неустойчивость климатических условий, например условия от более суровых, с малыми осадками и низкими температурами, до благоприятных с относительно большим слоем свежевыпавшего снега и низкими температурами.
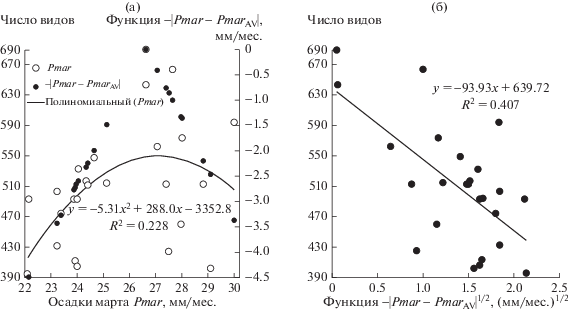
Отметим также очень тесную положительную связь осадков марта с суммой осадков зимы в регионе (R2 = 0.922, P < 10–6). Чаще всего защитную роль для почв играет не только свежевыпавший мартовский снег, но и снег, выпавший в течение всей зимы. Старый снег более плотный и имеет более высокую теплопроводность; он слабее защищает почву от охлаждения, чем свежевыпавший. Но их совместное действие из-за этой положительной связи усиливает защиту почв от охлаждения при больших осадках в марте, а малым осадкам марта часто отвечают и малые зимние осадки. Поэтому осадки марта играют на изучаемой территории Среднего Поволжья особую роль. Связь числа видов растений с осадками марта, представленная на рис. 2, демонстрирует резкий пик при PmarAV = 26.65 мм/месяц (среднее осадков марта по точкам наблюдения), который хорошо описывает функция |Pmar–PmarAV|1/2.
Рис. 2.
Зависимость числа видов от осадков марта (белые кружки) с резким пиком: квадратичный тренд показан кривой, значения функции –|Рmar–PmarAV| – черными кружками (а), и парная связь числа видов с |Рmar–PmarAV|1/2 (б).
При использовании в качестве предикторов характеристик климата, абсолютной высоты и типов почв полученное уравнение множественной регрессии для числа видов Nsp имеет следующий вид:
(1)
$\begin{gathered} Nsp = - 135.0\left| {Pmar{\kern 1pt} --{\kern 1pt} Pma{{r}_{{{\text{AV}}}}}} \right|_{{--7.05}}^{{1/2}} + \\ + \,\,98.46Toc{{t}_{{ + 4.54}}} + 0.4115(Pwin{\kern 1pt} --{\kern 1pt} Pwi{{n}_{{{\text{AV}}}}})_{{ + 3.34}}^{2} + \\ + \,\,95.11(Tmar{\kern 1pt} --{\kern 1pt} Tma{{r}_{{{\text{AV}}}}})_{{ + 2.60}}^{2} + {\text{ }}193.5; \\ {{R}^{2}} = {\text{ }}0.742(Degr = {\text{ }}26\% ),\,\,\,\,P < {\text{ }}{{10}^{{--4}}}, \\ \end{gathered} $Для парной связи числа видов с |Pmar–PmarAV|1/2 имеем R2 = 0.407, в то время как R2 для парной связи числа видов с остальными предикторами значительно меньше (0.033, 0.020 и 0.007 соответственно). Поэтому мы используем именно |Pmar–PmarAV|1/2. При учете совместного действия предикторов ситуация становится менее контрастной, как описано значениями модулей t-статистик в уравнении (1), где t-статистики подписаны нижними индексами. Взяв сумму модулей t-статистик (17.53) за 100%, получаем следующую оценку относительного вклада предикторов в модель (1) по значениям t-статистик: 40, 26, 19 и 15%.
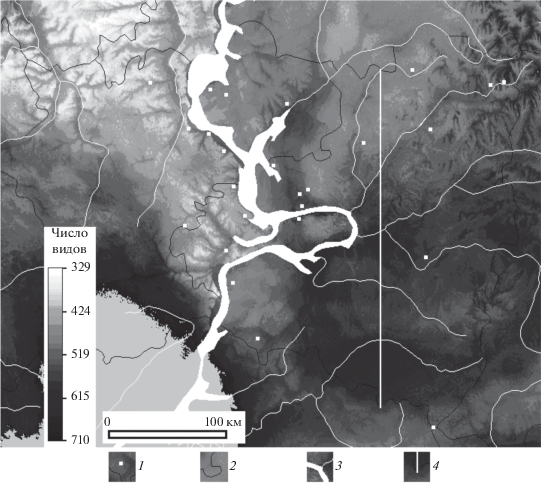
Рис. 3.
Карта пространственного распределения богатства видов, построенная по модели (1). На карте также показаны: 1 – центры площадок наблюдения, 2 – границы административных областей, 3 – водные объекты и 4 – трансекта.
Рассмотрим подробнее экологическую интерпретацию вошедших в модель (1) предикторов богатства видов сосудистых растений.
При малых Pmar по сравнению с PmarAV предиктор |Pmar–PmarAV|1/2 принимает большие значения, и поскольку связь с ним отрицательная, то богатство видов при малых Pmar уменьшается (см. рис. 2а). Этому способствует как избыточное охлаждение почв из-за уменьшения снежного покрова, так и недостаток влаги весной. Описанный эффект поддерживается выраженной согласованностью между осадками марта и зимы. Охлаждение почв и недостаток влаги отрицательно сказываются на видовом разнообразии при Pmar < < PmarAV. Увеличение осадков в марте (Pmar > > PmarAV) также снижает богатство видов из-за избыточного увлажнения почвы. Резкий пик (см. рис. 2а) вызван усилением первого и второго эффектов тесной связью между осадками марта и зимы. Среднее значение предиктора |Pmar–PmarAV|1/2 по точкам наблюдения составляет (2.17 ± 1.25 мм)1/2; стандартному отклонению 1.25 отвечает отклонение Pmar от PmarAV на 1.6 мм/месяц. Снижение богатства видов при этом отклонении относительно Pmar = PmarAV, согласно модели (1), при прочих равных предикторах составляет 211.
Второй предиктор – температура октября (Toct) – имеет положительную и линейную связь с видовым богатством сосудистых растений. В октябре температура еще положительна, но невелика (4.5 ± 0.5°С), поэтому на площадках с увеличенными значениями температуры можно ожидать и большее число видов, а на более холодных площадках – уменьшенное. Видимо, это связано с граничным положением октября, так как в ноябре в Среднем Поволжье отмечаются морозные температуры. Возрастание температур октября на 0.5°С (стандартное отклонение) приводит к тому, что в более теплых в этом месяце местах число видов на 49 больше при прочих равных предикторах. Однако это в 4.3 раза меньше оценки падения на 211 при изменении осадков в марте.
Третий предиктор – (Pwin–PwinAV)2, где Pwin – количество осадков зимой. Положительная связь с этим предиктором означает, что число видов возрастает как на площадках с увеличенными, так и уменьшенными относительно среднего значениями Pwin (PwinAV = 102 ± 10 мм). Взяв для оценок стандартное отклонение (10 мм), из уравнения (1) получаем рост числа видов на 41 при прочих равных предикторах. Заметим, что это в пять раз меньше, чем при оценке из-за отклонения осадков марта от среднего. Таким образом, отклонения как осадков марта, так и суммы осадков зимы от их средних приводят к изменению видового богатства сосудистых растений. Знаки этих изменений противоположны, что корректирует первый предиктор при больших изменениях Pwin.
Связь с четвертым предиктором (Tmar–TmarAV)2 положительна, что означает падение числа видов как при возрастании, так и при уменьшении температуры в марте от среднего значения по площадкам TmarAV. Заметим, что Tmar в тенденции снижается с ростом Pmar. Более теплые условия ранней весной (повышение Tmar) приводят к увеличению числа видов, что вполне ожидаемо в тех местах региона, где такое повышение происходит. Уменьшенным Tmar отвечают площадки, рост числа видов на которых предположительно связан с тем, что на них встречаются другие виды, более приспособленные к холодной весне. Но эффект повышения мартовских температур небольшой: среднее значение TmarAV = –5.5 ± 0.4°С, и для оценки положим отклонение Tmar от TmarAV равным стандартному отклонению 0.4°С, что дает рост числа видов на 15; это на порядок меньше, чем оценка 211 для Pmar. В действительности предикторы меняются не по одному, а совместно.
По модели (1) для данных разрешения 10.8 км была построена карта пространственного распределения богатства видов. Для улучшения визуального восприятия карты мы применили уравнение (1) к матрицам большего разрешения (рис. 3). Это мало влияло на значения предсказываемого моделью числа видов.
Богатство видов заметно увеличено на юго-востоке региона, в степной зоне, и уменьшено на северо-западе и северо-востоке региона. Предсказанные значения богатства видов менялись для региона от 357 до 709, что близко к диапазону в точках наблюдения (395–690), но несколько шире. Этого следовало ожидать, так как климатические факторы в регионе меняются в более широких пределах, чем в точках наблюдения. Изменение богатства видов сосудистых растений для степей, лугов и дубрав вдоль трансекты с юга на север Самарской области (см. рис. 3) показано на рис. 4. Для построения этого графика использовали более точные значения, полученные по модели (1) при шаге решетки матрицы 10.8 км. За ноль здесь взято расстояние от точки пересечения южной границы Самарской области с трансектой. Видно, как меняется богатство видов от степи к лугам и дубравам. Линейный характер графика не означает, что число видов во всем регионе линейно зависит от широты, т.е. если брать среднее число видов (среднее вдоль параллели) как функцию широты для всей местности, то такая зависимость уже не будет линейной, поскольку на юго-западе региона, в Ульяновской области, нет степи.
Рис. 4.
Рассчитанное по модели (1) число видов как функция расстояния по трансекте от южной границы Самарской области на север (штриховая линия показывает линейный тренд).
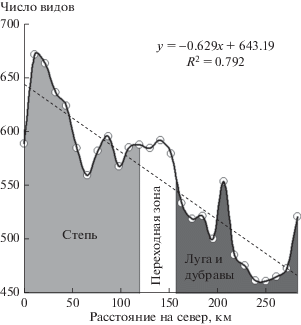
На юге этой трансекты лежит степь, затем следует сравнительно узкая переходная зона, включающая пойменные луга, а север занимает лесостепь с преобладанием лугов и редких дубрав. На трансекте наблюдается тесная связь богатства видов с расстоянием на север (n = 27, R2 = 0.792, P < 10–6).
В юго-западном участке карты, где не было точек наблюдения, количество зимних осадков возрастало до 150 мм, что выходит за пределы диапазона значений Pwin исследуемых площадок (от 82 до 122 мм). Поскольку выход далеко за пределы диапазона калибровки может быть источником существенных ошибок в модели [31], этот участок, где было предсказано высокое богатство видов (1323), был исключен из рассмотрения (см. рис. 3).
Для сравнения рассмотрим модель, в которой использовались только годовые характеристики климата, высота над уровнем моря и типы почв. Типы почв учитывали с помощью индикаторной переменной Chern, равной единице для черноземов и нулю для других типов почв. В результате было получено следующее уравнение:
(2)
$\begin{gathered} Nsp = --27.61\,Chern\,\,Tyea{{r}_{{--2.39}}}-- \\ - \,\,0.7386{{Z}_{{--2.24}}} + \,\,14.86\,dTyea{{r}_{{ + 2.14}}} + \\ + \,\,62.60\,Chern\,\,\left( {Tyear--Tyea{{r}_{{{\text{AV}}}}}} \right)_{{ + 0.66}}^{2} + 654.5; \\ {{R}^{2}} = \,\,0.583\,\,(Degr = {\text{ }}214\% ),\,\,\,\,P < 0.01. \\ \end{gathered} $Здесь Tyear – среднегодовая температура, Z – высота над уровнем моря, dTyear – амплитуда среднегодовой температуры, т.е. разность между средним максимумом температуры наиболее теплого месяца и средним минимумом температуры наиболее холодного месяца.
Эта модель объясняет значительно меньший процент дисперсии числа видов (58%) по сравнению с моделью (1), которая объясняет 74% дисперсии. При использовании только годовых показателей климата подобная связь как здесь, так и в региональных исследованиях других авторов была значительно менее тесной, объясняя, например, 50% дисперсии богатства видов высших сосудистых растений Австрийских Альп в работе [24], где размеры площадок (5.9 км) близки к нашим (10 км).
Таким образом, без учета климатических месячных и сезонных характеристик обнаруживается значительно менее тесная связь между богатством видов и факторами среды. Это подтверждает нашу гипотезу о существенном влиянии распределения характеристик климата по месяцам и сезонам на видовое разнообразие сосудистых растений в исследованном регионе. Отметим, что в модель (1), кроме зимних осадков, вошли климатические характеристики “пограничных” месяцев (март и октябрь), когда температура воздуха в соседних месяцах меняет знак (см. рис. 1а).
Существенным для выявления тесной связи богатства видов с факторами среды является и то, что в изучаемом регионе зависимость богатства видов от осадков марта имеет резко нелинейный характер (см. рис. 2а). Подобный пик описан в работе [12] для трансект в тропиках, который был согласован с резким изменением характера увлажнения в горах. Главный предиктор в нашей модели (1) – модуль разности осадков марта – также отражает критические изменения условий увлажнения, важные для богатства видов на исследуемой равнинной территории.
Таблица 1.
Характеристики исследовательских площадок
| № площадки | Координаты центров площадок | Число семейств | Число родов | Число видов | Ошибка* | |
|---|---|---|---|---|---|---|
| широта | долгота | |||||
| 1 | 48°51′12′′ | 54°22′18′′ | 74 | 391 | 493 | 21.8 |
| 2 | 49°36′52′′ | 53°29′21′′ | 78 | 357 | 643 | –61.6 |
| 3 | 49°36′42′′ | 53°35′01′′ | 77 | 332 | 562 | –27.8 |
| 4 | 49°35′33′′ | 54°14′27′′ | 97 | 338 | 690 | 75.0 |
| 5 | 49°18′00′′ | 53°47′21′′ | 81 | 333 | 515 | –3.7 |
| 6 | 48°41′03′′ | 54°25′39′′ | 74 | 349 | 494 | 8.7 |
| 7 | 49°41′37′′ | 53°36′57′′ | 76 | 313 | 513 | –10.5 |
| 8 | 48°52′16′′ | 52°36′18′′ | 81 | 326 | 512 | –33.2 |
| 9 | 49°32′26′′ | 53°25′09′′ | 81 | 342 | 664 | 59.3 |
| 10 | 48°21′28′′ | 54°09′06′′ | 73 | 358 | 474 | 32.9 |
| 11 | 48°45′35′′ | 53°57′16′′ | 76 | 380 | 533 | 17.6 |
| 12 | 48°35′26′′ | 54°06′22′′ | 73 | 280 | 503 | 8.2 |
| 13 | 48°38′03′′ | 53°15′52′′ | 66 | 269 | 493 | 17.7 |
| 14 | 50°59′53′′ | 53°00′35′′ | 91 | 356 | 594 | 11.7 |
| 15 | 47°57′03′′ | 54°31′07′′ | 71 | 321 | 405 | –17.6 |
| 16 | 51°17′40′′ | 53°54′29′′ | 73 | 269 | 512 | 21.2 |
| 17 | 51°11′47′′ | 54°20′13′′ | 71 | 270 | 460 | –15.2 |
| 18 | 52°06′57′′ | 54°08′37′′ | 70 | 255 | 424 | –70.7 |
| 19 | 52°17′04′′ | 54°09′24′′ | 83 | 321 | 574 | 7.8 |
| 20 | 50°27′49′′ | 53°53′55′′ | 61 | 217 | 402 | –56.4 |
| 21 | 50°46′39′′ | 51°47′07′′ | 61 | 300 | 548 | 30.1 |
| 22 | 48°54′59′′ | 53°28′56′′ | 64 | 234 | 412 | –12.5 |
| 23 | 48°09′59′′ | 53°27′23′′ | 75 | 290 | 517 | 77.5 |
| 24 | 48°49′24′′ | 53°42′36′′ | 71 | 261 | 432 | –4.0 |
| 25 | 48°39′14′′ | 53°01′34′′ | 70 | 239 | 395 | –76.4 |
Список литературы
Уиттекер Р. Сообщества и экосистемы. М.: “Прогресс”, 1980. 327 с.
Морозова О.В. Пространственные тренды таксономического богатства флоры сосудистых растений // Биосфера. 2011. Т. 3. № 2. С. 190–207.
MacArthur R.H., Wilson E.O. The Theory of Island Biogeography. Princeton, N.J.: Princeton University Press, 1967. 203 p.
Grime J.P. Plant strategies and vegetation processes. Chichester: J. Wiley, 1979. 222 p.
Taylor D.R., Aarssen L.W., Loehle C. On the relationship between r/K selection and environmental carrying capacity: a new habitat templet for plant life history strategies // Oikos. 1990. V. 58. P. 239–250.
Currie D.J. Energy and large-scale patterns of animal- and plant-species richness // The American Naturalist. 1991. V. 137. P. 27–49.
Richerson P.J., Lum K.-L. Patterns of plant species diversity in California: relation to weather and topography // The American Naturalist. 1980. V. 116. P. 504–536.
Axmanová I., Chytrý M., Zelený D. et al. The species richness–productivity relationship in the herb layer of European deciduous forests // Global Ecology and Biogeography. 2012. V. 21. P. 657–667.
Морозова О.В. Таксономическое богатство флоры Восточной Европы: факторы пространственной дифференциации. М.: Наука, 2008. 328 с.
Шарый П.А., Шарая Л.С., Сидякина Л.В., Саксонов С.В. Влияние солнечной энергии и сомкнутости крон деревьев на богатство видов травянистой растительности юга лесостепи // Сибирский экологич. журн. 2017. № 5. С. 539–552. https://doi.org/10.15372/SEJ20170502
Colwell R.K., Rahbek C., Gotelli N.J. The mid-domain effect and species richness patterns: what have we learned so far? // The American Naturalist. 2004. V. 163. P. E1–E23.
Kessler M., Kluge J., Hemp A., Ohlemüller R. A global comparative analysis of elevational species richness patterns of ferns // Global Ecology and Biogeography. 2011. V. 20. P. 868–880.
Csergö A.M., Salguero-Gómez R., Broennimann O. et al. Less favourable climates constrain demographic strategies in plants // Ecology Letters. 2017. V. 12. https://doi.org/10.1111/ele.12794
Palmer M.W. Variation in species richness: towards a unification of hypotheses // Folia Geobot. Phytotax. Praha. 1994. V. 29. P. 511–530.
Бигон М., Харпер Дж., Таунсенд К. Экология. Особи, популяции и сообщества. Т. 2. М.: Мир, 1989. 477 с.
Миркин Б.М., Наумова Л.Г. Современное состояние основных концепций науки о растительности. Уфа: “Гилем”, 2012. 488 с.
Hawkins B.A., Field R., Cornell H.V. et al. Energy, water, and broad-scale geographic patterns of species richness // Ecology. 2003. V. 84. P. 3105–3117.
Davies T.J., Savolainen V., Chase M.W. et al. Environmental energy and evolutionary rates in flowering plants // Proceedings of the Royal Society of London. Ser. B. 2004. V. 271. P. 2195–2200.
Storch D., Evans K.L., Gaston K.J. The species-area-energy relationship // Ecology Letters. 2005. V. 8. P. 487–492.
Weiher E., Howe A. Scale-dependence of environmental effects on species richness in oak savannas // J. of Vegetation Science. 2003. V. 14. P. 917–920.
Kreft H., Sommer J.H., Barthlott W. The significance of geographic range size for spatial diversity patterns in Neotropical palms // Ecography. 2006. V. 29. P. 21–30.
Field R., Hawkins B.A., Cornell H.V. et al. Spatial species-richness gradients across scales: a meta-analysis // J. of Biogeography. 2009. V. 36. P. 132–147.
Hijmans R.J., Cameron S.E., Parra J.L. et al. Very high resolution interpolated climate surfaces for global land areas // International J. of Climatology. 2005. V. 25. P. 1965–1978.
Moser D., Dullinger S., Englisch T. et al. Environmental determinants of vascular plant species richness in the Austrian Alps // J. of Biogeography. 2005. V. 32. P. 1117–1127.
Саксонов С.В., Савенко О.В., Иванова А.В., Конева Н.В. Флора Сусканского заказника в Самарской области (Низменное Заволжье, Мелекесско-Ставропольский флористический район) // Фиторазнообразие Восточной Европы. 2007. № 2. С. 125–156.
Иванова А.В., Сенатор С.А., Саксонов С.В., Раков Н.С. Материалы к флоре урочища Байтуган Камышлинского района Самарской области // Фиторазнообразие Восточной Европы. 2011. № 9. С. 187–217.
Савенко О.В., Саксонов С.В., Сенатор С.А. Материалы для флоры Узюковского лесного массива // Исследования в области естественных наук и образования: Межвуз. сб. науч.-исслед. работ. Самара, 2011. Вып. 2. С. 48–53.
Раков Н.С. Флора села Большое Нагаткино (Ульяновское Предволжье) // Самарская Лука: проблемы региональной и глобальной экологии. 2015. Т. 24. № 2. С. 125–154.
Сенатор С.А., Саксонов С.В., Раков Н.С. и др. Сосудистые растения Тольятти и окрестностей (Самарская область) // Фиторазнообразие Восточной Европы. 2015. Т. 9. № 1. С. 32–101.
Иванова А.В., Костина Н.В., Кузнецова Р.С. Взаимосвязь флористического и ландшафтного разнообразия территории на примере физико-географического района лесостепной зоны // Изв. Саратовского ун-та. Нов. серия. Химия, биология, экология. 2017. Вып. 4. С. 481–485.
Montgomery D.C., Peck E.A. Introduction to linear regression analysis. New York: John Wiley & Sons, 1982. 504 p.
Шарый П.А., Пинский Д.Л. Статистическая оценка связи пространственной изменчивости содержания органического углерода в серой лесной почве с плотностью, концентрацией металлов и рельефом // Почвоведение. 2013. № 11. С. 1344–1356. https://doi.org/10.7868/S0032180X13090104
Lischke H., Guisan A., Fischlin A., Bugmann H. Vegetation responses to climate change in the Alps – Modeling studies / Cebon P., Dahinden U., Davies H., Imboden D., Jaeger C. Eds. A view from the Alps: Regional perspectives on climate change. Boston: MIT Press, 1998. Chapter 6. P. 309–350.
Дополнительные материалы отсутствуют.