

Экология, 2020, № 3, стр. 224-229
Периодические изменения численности средней бурозубки (Sorex caecutiens, Eulipotyphla) в предгорной тайге Северного Урала и факторы ее популяционной динамики
А. В. Бобрецов a, b, *, Л. Е. Лукьянова c, А. Н. Петров b, Н. М. Быховец b
a Печоро-Илычский государственный природный биосферный заповедник
169436 Якша, ул. Ланиной, 8, Россия
b Институт биологии Коми научного центра УрО РАН
167982 Сыктывкар, ул. Коммунистическая, 28, Россия
c Институт экологии растений и животных УрО РАН
620144 Екатеринбург, ул. 8 Марта, 202, Россия
* E-mail: avbobr@mail.ru
Поступила в редакцию 27.09.2019
После доработки 21.10.2019
Принята к публикации 14.11.2019
Аннотация
Проанализированы многолетние данные колебаний численности средней бурозубки (Sorex caecutiens Laxmann) в предгорной тайге Северного Урала (Печоро-Илычский заповедник). Обнаружены периодические составляющие в популяционной динамике вида и выявлены факторы, влияющие на изменчивость показателей обилия. Для анализа временного ряда использовали спектральный анализ. Выявлена статистически значимая периодическая составляющая, равная 3 годам, которая не являлась строго регулярной. На динамику популяции средней бурозубки оказывают влияние как погодные условия, так и плотностно-зависимые механизмы регуляции численности вида.
Проблема динамики численности мелких млекопитающих занимает центральное место в популяционных исследованиях [1]. Вместе с тем в отношении землероек закономерности изменения их обилия, а также причины, обусловливающие эту вариабельность, менее известны. Кроме того, большинство работ по динамике популяций данной группы животных выполнено на обыкновенной бурозубке. У землероек отмечены как регулярные с определенным периодом (чаще всего 3–4 года) изменения численности [2–6], так и нерегулярные [7–9]. Первые часто называются циклическими колебаниями, хотя строгая цикличность и в этом случае отсутствует.
Многолетние изменения численности мелких млекопитающих, в том числе и землероек, определяются целым рядом внешних факторов и внутрипопуляционных механизмов [1, 10, 11]. Важную роль в динамике популяций землероек играют погодные условия [7, 12, 13], влияние которых особенно повышается в переходные сезоны года − в начале зимы и весной [3, 14, 15]. Однако в некоторых регионах такая связь отсутствует [16, 17]. По мнению других авторов [5, 6, 18], ведущими факторами периодических колебаний численности землероек являются плотностно-зависимые механизмы.
Следует отметить, что число публикаций по динамике популяций средней бурозубки невелико. Поэтому данная работа вносит определенный вклад в решение рассматриваемой проблемы. В ней анализируются результаты многолетних исследований по изменению численности данного вида в предгорном районе Печоро-Илычского заповедника. Целью работы было выявление периодичности в популяционной динамике средней бурозубки и анализ факторов, влияющих на изменчивость ее численности.
МАТЕРИАЛ И МЕТОДЫ
Средняя бурозубка (Sorex caecutiens Laxmann) − один из фоновых видов мелких млекопитающих предгорий Северного Урала, по численности уступает лишь обыкновенной бурозубке. Количественные учеты землероек проводили в августе 1988–2018 гг. в верховьях р. Печоры в урочище Гаревка (62°05′ с.ш., 58°27′ в.д.). Большая часть территории (76%) занята полидоминантной темнохвойной тайгой. Для лесов характерно мощное развитие мохового покрова, что является следствием высокой влажности и близостью к горным хребтам Северного Урала. Тесная связь данного вида с лесными сообществами таежного типа хорошо известна, и, следовательно, этот район вполне благоприятен для существования здесь средней бурозубки.
Учеты землероек проводили ловчими канавками и давилками в первой половине августа. Для анализа использовали данные по обилию землероек только по ельнику зеленомошному (наиболее благоприятному местообитанию для средней бурозубки). Усредненные данные по нескольким разным биотопам занижают оценки обилия и искажают картину динамики видов [19]. Канавки длиной 50 м с 5 конусами открывали на 12–15 дней. За показатель относительной численности принимали число животных, отловленных на 100 конусо-суток (ос. на 100 кон.-сут). Давилки выставляли в линию по 100 шт. через 5 м на четверо суток. В этом случае за показатель относительной численности принимали число животных, отловленных на 100 ловушко-суток (ос. на 100 лов.-сут). За период работы отработано 3250 кон.-сут и поймано 1857 особей, в давилки соответственно 13 400 ловушко-суток и 163 экз. Доля вида в суммарных уловах мелких млекопитающих в канавках составила 25.7%, тогда как в давилках − 8.2%.
Для выявления и оценки периодичности во временных рядах использовали метод спектрального анализа. Предварительно данные логарифмировали, так как распределение годовых показателей обилия уклонялось от нормального (соответствие проводили с помощью критерия Колмогорова-Смирнова). Целью метода является разложение временного ряда на отдельные составляющие (гармоники), которые имеют различные амплитуду и частоту. Наиболее мощные из них с ярко выраженными пиками на определенных частотах периодограммы и представляли периодические составляющие. Для выявления связей показателей численности животных с различными факторами использовали коэффициент ранговой корреляции Спирмена. Статистическая обработка материала проведена с использованием программного пакета Statistica 6.0 for Windows.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Изменение численности средней бурозубки по годам
Численность средней бурозубки за период наблюдений по отловам в канавки изменялась по годам от 2.6 (2018 г.) до 205.5 (2013 г.) ос. на 100 кон.-сут, по отловам в давилки − от 0 до 9.3 ос. на 100 лов.-сут (2013 г.). В годы депрессий землеройки часто в уловах давилками отсутствовали. Годы минимумов и максимумов обилия по данным обоих методов в общем совпадали (рис. 1), особенно это было характерно для максимумов численности. Максимальные показатели численности землероек (максимальные значения после депрессий) регистрировали на 2, 3, 4 и 5-й год, чаще всего на 3-й год.
Рис. 1.
Динамика численности средней бурозубки в ельнике зеленомошном предгорного района Печоро-Илычского заповедника по отловам в канавки (1) и давилки (2).
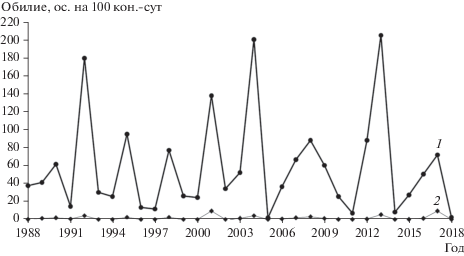
Спектральный анализ выявил статистически значимую периодическую составляющую, равную 3 годам, только по уловам в канавки, тогда как по отловам в давилки она была недостоверной (рис. 2). В последнем случае на результатах сказались невысокие показатели численности. Вместе с тем данные, полученные при помощи канавок и давилок, коррелировали между собой (r = +0.82, p < 0.001).
Рис. 2.
Периодограмма варьирования численности средней бурозубки в предгорном районе Печоро-Илычского заповедника (штриховые линии – уровни значимости) по отловам в канавки (1) и давилки (2).
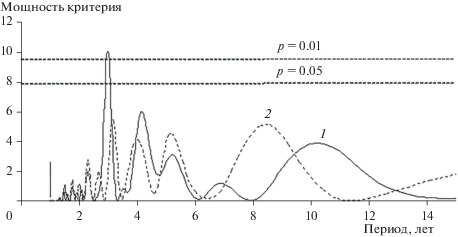
Периодические (циклические) колебания 3‑летней протяженности были обнаружены у средней бурозубки в бассейне Верхней Колымы [5]. В Центральной Сибири 4-летняя периодичность у многих видов мелких млекопитающих во второй половине 1990-х годов прошлого века сменилась сначала хаотическими изменениями, а затем в них стала улавливаться тенденция к 3-летним колебаниям обилия [4]. В Пинежском заповеднике (Архангельская область) у землероек отмечены подъемы численности один раз в три года [20]. По мнению Л.Н. Ердакова с соавт. [6], во многих популяциях обыкновенной бурозубки хорошо проявляется 3-летняя периодическая составляющая, но она невелика по мощности.
Вклад погодных и других внешних факторов в формирование динамики средней бурозубки
Физиологические особенности землероек (малые размеры тела, увеличение энергетических затрат) определяют высокую степень зависимости представителей этой группы животных от погодных условий [21]: они влияют на жизнедеятельность бурозубок как напрямую (температура воздуха и осадки), так и косвенно (через корма). Считается, что наиболее значимую роль играют “переходные” периоды − весна и осень [3, 14, 15].
Нами прежде всего было проанализировано влияние на численность землероек показателей средних месячных температур весны и осени, а также увлажнения. Коэффициенты корреляции для весеннего периода оказались статистически незначимыми (табл. 1). Невысокое достоверное влияние на численность землероек оказывают только температура воздуха октября предыдущего года, а также температурные показатели в целом за осень. При высокой температуре в эти периоды выживаемость животных выше, чем в другие годы. Отмечено также положительное воздействие количества осадков в августе текущего года. Возможно, это связано с тем, что при повышении влажности доступность кормов возрастает. Связь численности землероек с осадками отмечена во многих регионах [7, 18].
Таблица 1.
Оценка влияния разных климатических факторов на динамику численности средней бурозубки
| Фактор | Статистические параметры | ||
|---|---|---|---|
| r | t | p | |
| Температура воздуха в апреле текущего года | –0.04 | 0.21 | >0.05 |
| Температура воздуха в мае текущего года | –0.20 | 1.12 | >0.05 |
| Температура воздуха в октябре предыдущего года | +0.36 | 2.10 | <0.05 |
| Температура воздуха осенью предыдущего года | +0.43 | 2.60 | <0.01 |
| Сумма осадков в августе текущего года | +0.37 | 2.14 | <0.05 |
Число значимых климатических факторов, контролирующих обилие землероек, в разных регионах может существенно варьировать. На примере обыкновенной бурозубки было показано [7, 22, 23], что в разных частях ареала оно колебалось от 3 до 13.
Условия зимнего периода в Печоро-Илычском заповеднике большой роли для землероек не играют. Это связано с тем, что территория заповедника относится к одним из многоснежных в европейской части России (средняя глубина снега составляет 105 см), что нивелирует резкие перепады температуры воздуха у поверхности земли. Снег в этом районе ложится довольно рано, вследствие чего смертность землероек в зимний период здесь в целом незначительна. Отсюда связь между численностью землероек и глубиной снега, отмеченная в других регионах [15, 24], в Печоро-Илычском заповеднике отсутствует (r = +0.18, p = = 0.28).
Более того, даже для близких в географическом плане территорий влияние климатических факторов оценивается порою противоречиво. Так, Н.Е. Докучаев [12] считает, что в Северо-Восточной Азии условия зимнего периода наиболее важны в регулировании численности средней бурозубки. По другим данным [5] они не вносят существенного вклада в формирование динамики популяции этого вида. Слабое влияние они оказывают также и на короткохвостую землеройку Blarina brevicauda в условиях довольно сурового климата восточной части Северной Америки [25].
Однако погодные условия года могут дифференцированно воздействовать на популяцию животных на разных фазах цикла (табл. 2): число таких факторов в Печоро-Илычском заповеднике увеличивается, а коэффициенты корреляции имеют большие значения. На фазе подъема численности статистически значимыми являются четыре фактора. Чем меньше температура воздуха в августе предыдущего года, тем больше показатели численности в текущем году. Отрицательной является и связь обилия землероек с суммой осадков в июле текущего года – в этом случае большое количество осадков замедляет подъем численности животных. В то же время большое количество осадков в ноябре и декабре предыдущего года в виде снега положительно влияет на рост показателей обилия. Роль осадков довольно велика и на фазе снижения численности.
Таблица 2.
Влияние климатических факторов на численность средней бурозубки на разных фазах цикла
| Фактор | Статистические параметры | ||
|---|---|---|---|
| r | t | p | |
| Фаза депрессии | |||
| Температура воздуха в октябре предыдущего года | –0.68 | 2.52 | <0.05 |
| Фаза увеличения численности | |||
| Температура воздуха в августе предыдущего года | –0.70 | 2.59 | <0.05 |
| Сумма осадков в июле текущего года | –0.75 | 3.00 | <0.01 |
| Сумма осадков в ноябре предыдущего года | +0.92 | 6.07 | <0.01 |
| Сумма осадков в ноябре–декабре предыдущего года | +0.73 | 2.95 | <0.05 |
| Фаза снижения численности | |||
| Сумма осадков текущего года | –0.80 | 2.82 | <0.05 |
На разных фазах цикла хорошо прослеживается связь с началом наступления фенологических явлений в весенний и осенний периоды, хотя при анализе всего временнόго ряда она отсутствует. На фазе пика большое значение имеют сроки схода снега в лесу: чем раньше сходит снег, тем больше землероек в текущем году (r = –0.66; p < 0.05). Глубина депрессий определяется сроками начала установления постоянного снежного покрова в предыдущем году: чем раньше выпадает снег, тем выше значения численности на следующий год. Поздние сроки установления постоянного снежного покрова приводят к глубоким депрессиям (r = –0.60; p < 0.05).
Одним из факторов динамики численности вида могут быть межвидовые взаимодействия. Например, в Восточной Фенноскандии численность средней бурозубки как менее “конкурентоспособного” вида увеличивается только в периоды низкой численности обыкновенной бурозубки [26]. В Печоро-Илычском заповеднике динамика популяций всех фоновых видов мелких млекопитающих синхронна. Так, значение коэффициента корреляции между показателями обилия этих близких видов землероек составляет +0.88 (p < 0.001). Поэтому межвидовая конкуренция в динамике популяции средней бурозубки на исследуемой территории не играет заметной роли.
Роль внутрипопуляционных факторов в динамике численности средней бурозубки
Для средней бурозубки предгорий Северного Урала выявлена корреляция ряда демографических параметров с уровнем численности землероек. В периоды минимумов значительно возрастает число перезимовавших животных, в периоды подъемов и максимумов доля взрослых падает – 0.57 (p < 0.001). При этом существенно меняется число перезимовавших самок. Связь доли самок с уровнем обилия положительна (r = +0.51; p < 0.01): их относительное число увеличивается при высокой численности животных. В период депрессий доля перезимовавших самок варьирует от 0 до 50% и в среднем составляет 25.7 ± 6.2%, а на фазе максимумов она равна 67.7 ± 6.7% (от 42.8 до 92.8%).
Различия в средней плодовитости перезимовавших самок заключаются в том, что в периоды низкой численности животных размер выводка значительно выше (6.6 ± 0.3 эмбриона), тогда как при высокой он уменьшается (5.9 ± 0.3 эмбриона).
Аналогичная зависимость была отмечена нами для обыкновенной бурозубки предгорного района Печоро-Илычского заповедника [23]. В средней полосе России плотностно-зависимые механизмы являются основной причиной периодических колебаний численности землероек [18]. Они же играют определенную роль в динамике популяций обыкновенной и средней бурозубок в Тверской области [14]. Достоверные различия в среднем размере выводка в зависимости от уровня плотности животных обнаружены у ряда видов землероек Прибайкалья [27]. Ведущая роль авторегуляторных механизмов в изменении численности выявлена у средней бурозубки (бассейн Верхней Колымы) [5].
Одним из эффективных механизмов внутренней регуляции является скорость созревания молодых животных и их участие в размножении. Участие сеголеток в репродукции отмечено в некоторых регионах [28], но в условиях Севера европейской части России оно не играет роли [29]. В Республике Коми доля размножающихся молодых самок у средней бурозубки колебалась по районам от 0.4 до 2% [23].
ЗАКЛЮЧЕНИЕ
В колебаниях численности средней бурозубки предгорий Северного Урала выявлена статистически значимая периодическая составляющая, равная трем годам. Чаще всего именно через такой период происходят подъемы обилия данного вида.
Погодные условия играют определенную роль в популяционной динамике средней бурозубки в регионе. Однако они влияют наиболее заметно в периоды подъемов численности, снижения и депрессий. Наиболее высокие значения коэффициента корреляции связаны с количеством осадков в ноябре–декабре предыдущего года, что отражается на выживаемости животных. Межвидовые отношения не оказывают существенного влияния на динамику численности средней бурозубки: ее обилие изменяется синхронно с другими фоновыми видами землероек и полевок.
Отмечена роль плотностно-зависимых механизмов в динамике популяции вида. В зависимости от уровня обилия животных изменяется число перезимовавших особей, доля в них взрослых самок, а также средний размер выводков.
Таким образом, периодические колебания численности землероек в предгорьях Северного Урала представляют собой результат взаимодействия внутрипопуляционных процессов с изменчивыми условиями внешней среды.
Работа выполнена в рамках проекта № 18-4-4-30 “Динамика разнообразия животного мира западного макросклона Урала и сопредельных территорий (равнинной части Европейского Северо-Востока России) в условиях изменения среды” (№ регистрации АААА-А17-117112850234-5), а также при поддержке Комплексной программы УрО РАН (№ 18-4-4-28).
Авторы приносят благодарность к.б.н., н.с. лаборатории популяционной экологии и моделирования ИЭРиЖ УрО РАН К.В. Маклакову за помощь в обработке временных рядов и интерпретации полученных статистических результатов.
Список литературы
Krebs C.J. Population fluctuations in rodents. Chicago: University of Chicago Press, 2013. 306 p.
Korpimäki E., Norrdahl K., Huitu O., Klemola T. Predator induced synchrony in population oscillations of coexisting small mammal species // Proceedings of the Royal Society B: Biological Sciences. 2005. V. 272. № 1559. P. 193–202.
Zub K., Jędrzejewska B., Jędrzejewski W., Bartoń K.A. Cyclic voles and shrews and non-cyclic mice in a marginal grassland within European temperate forest // Acta Theriol. 2012. V. 57. № 3. P. 205–216.
Шефтель Б.И. Циклическая динамика популяций мелких млекопитающих и глобальные климатические изменения // Млекопитающие Северной Евразии: жизнь в северных широтах: Мат-лы междун. науч. конф. Сургут: Сургутский гос. ун-т, 2014. С. 19–20.
Киселев С.В., Ямборко А.В. Динамика численности средней (Sorex caecutiens) и равнозубой (Sorex isodon) бурозубок в бассейне Верхней Колымы // Зоол. журн. 2014. Т. 93. № 9. С. 1106–1116.
Ердаков Л.Н., Панов В.В., Литвинов Ю.Н. Цикличность хода численности в разных популяциях обыкновенной бурозубки // Экология. 2019. Т. 6. С. 446–455.
Балакирев А.Е., Окулова Н.М., Ивантер Э.В. К анализу факторных воздействий на многолетнюю динамику численности обыкновенной бурозубки на севере и юге ареала // Поволжский экологич. журн. 2004. № 2. С. 111–122.
Popov I.Y. Structure and dynamics of shrews on permanent plots in the European southern taiga // Advances in the Biology of Shrews II (Special publication of the international Society of shrew biologists). New York, 2005. V. 1. P. 291–301.
Tomášková L., Bejček V., Sedlaček F. et al. Population biology shrews (Sorex araneus and S. minutus) from a polluted area in central Europe // Advances in the Biology of Shrews II (Special publication of the international Society of shrew biologists). New York, 2005. V. 1. P. 189–197.
Bjørnstad O.N., Grenfell B.T. Noisy clockwork: time series analysis of population fluctuations in animals // Science. 2001. V. 293. № 5530. P. 638–643.
Жигальский О.А. Анализ популяционной динамики мелких млекопитающих // Зоол. журн. 2002. Т. 81. № 9. С. 1078–1106.
Докучаев Н.Е. Экология бурозубок Северо-Восточной Азии. М.: Наука, 1990. 160 с.
Виноградов В.В. Многолетняя динамика и структура сообществ землероек (Soricidae) горной тайги Восточного Саяна // Сибирский экологич. журн. 2012. Т. 19. № 1. С. 131–139.
Калинин А.А., Демидова Т.Б., Олейниченко В.Ю., Щипанов Н.А. Сезонная динамика численности землероек-бурозубок (Insectivora, Soricidae) // Зоол. журн. 2008. Т. 87. № 2. С. 218–225.
Ивантер Э.В. Очерки популяционной экологии мелких млекопитающих на северной периферии ареала. М.: Тов-во науч. изд. КМК, 2018. 770 с.
Henttonen H., Haukisalmi V., Kaikusalo A. et al. Long-term population dynamics of the common shrew Sorex araneus in Finland // Annales Zoologici Fennici. 1989. V. 26. № 4. P. 349–355.
Churchfield S., Hollier J., Brown V.K. Population dynamics and survivorship patterns in the common shrew Sorex araneus in southern England // Acta Theriol. 1995. V. 40. № 1. P. 53–68.
Ходашева К.С., Елисеева В.И. Землеройки в экосистемах Центральной лесостепи Русской равнины. М.: Наука, 1992. 112 с.
Ивантер Э.В., Коросов А.В., Якимова А.Е. Эколого-статистический анализ многолетних изменений численности мелких млекопитающих на северном пределе ареала (Северо-Восточное Приладожье) // Экология. 2015. № 1. С. 57–63.
Куприянова И.Ф., Сивков А.В. Сообщества мелких млекопитающих заповедника // Структура и динамика природных компонентов Пинежского заповедника (северная тайга, ЕТР Архангельская область). Архангельск, 2000. С. 168–178.
Ochocińska D., Taylor J.R.E. Living at the physiological limits: field and maximum metabolic rates of the common shrew (Sorex araneus) // Physiological Biochemical Zoology. 2005. V. 78. № 5. P. 808–818.
Окулова Н.М., Куприянова И.Ф., Сивков А.В. Динамика численности мелких млекопитающих Пинежского заповедника. Сообщ. 1. Обыкновенная бурозубка Sorex araneus // Териологические исследования. СПб., 2003. Вып. 4. С. 38–46.
Бобрецов А.В. Популяционная экология мелких млекопитающих равнинных и горных ландшафтов Северо-Востока европейской части России. М.: Тов-во науч. изд. КМК, 2016. 381 с.
Hlôška L., Chovancová B., Chovancová G., Fleischer P. Influence of climatic factors on the population dynamics of small mammals (Rodentia, Soricomorpha) on the sites affected by windthrow in the High Tatra Mts // Folia Oecologica. 2016. V. 43. № 1. P. 12–20.
Lima M., Merritt J.F., Bozinovic F. Numerical fluctuations in the northern short-tailed shrew: evidence of non-linear feedback signatures on population dynamics and demography // J. of Animal Ecology. 2002. V. 71. № 2. P. 159–172.
Ивантер Э.В., Курхинен Ю.П. Влияние антропогенной трансформации лесных экосистем Восточной Фенноскандии на популяции малой и средней бурозубок // Принципы экологии. 2014. № 2. С. 21–25.
Малышев Ю.С. Средняя бурозубка – Sorex caecutiens Laxmann, 1758 Верхнеангарской котловины: численность, ландшафтное распределение, особенности структуры и репродукции популяции // Байкальский зоологич. журн. 2014. № 2(15). С. 92–102.
Dokuchaev N.E. Reproduction of shrews (Sorex) in Siberia // Advances in the biology of shrews II (Special publication of the international Society of shrew biologists). New York, 2005. V. 1. P. 419–431.
Shchipanov N.A., Zima J., Churchfield S. Introducing the common shrew // Shrews, Chromosomes and Speciation (Cambridge Studies in Morphology and Molecules: New Paradigms in Evolutionary Bio). Searle J., Polly P., Zima J. Eds. Cambridge: Cambridge University Press, 2019. P. 19–67.
Дополнительные материалы отсутствуют.