

Экология, 2020, № 3, стр. 181-189
Поиск и выявление системы плейстоценовых рефугиумов вида Pinus sylvestris L.
С. Н. Санников a, И. В. Петрова a, Е. В. Егоров a, Н. С. Санникова a, *
a Ботанический сад УрО РАН
620144 Екатеринбург, ул. 8 Марта, 202а, Россия
* E-mail: sannikovanelly@mail.ru
Поступила в редакцию 07.08.2019
После доработки 18.09.2019
Принята к публикации 08.10.2019
Аннотация
На основе сравнительного аллозимного анализа минимальных генетических дистанций Неи между 8 филогеографическими группами популяций (ФГП) Pinus sylvestris L. “ледниковой” зоны и 26 популяциями южной “внеледниковой” зоны в пределах всего ареала выявлена локализация 9 основных и 8 второстепенных гипотетичных плейстоценовых рефугиумов (ГПР) этого вида. С помощью эмпирических “инвестиционных индексов рефугиумов” сопоставлен относительный вклад их генофонда в генофонд филогеографических групп ледниковой зоны. Показано полирефугиальное происхождение популяций этой зоны. Предложена классификация плейстоценовых рефугиумов P. sylvestris по их локализации в ареале и роли в формировании современного генофонда вида.
Одна из ключевых проблем современной эволюционной биологии – выявление места, времени происхождения и путей миграции популяций в различных частях ареала видов. Несмотря на бурное развитие молекулярно-генетических методов в последние 50 лет, ответ на эти вопросы для большинства видов растений и животных остается “книгой за семью печатями”. Общим методологическим недостатком геногеографических исследований популяций растений, в частности Pinus sylvestris L., на наш взгляд, являются их региональная фрагментарность, различия и несопоставимость спектров изучаемых изозимов или гаплотипов и, как следствие, отсутствие информации о структуре и генетических связях популяций в пределах всего ареала вида.
В последнее время широко обсуждается вопрос о местонахождении ледниковых рефугиумов Pinus sylvestris L. – одного из главных лесообразующих видов Северной Евразии. В этом плане в Западной и Центральной Европе проведены многие палеоботанические и популяционно-генетические исследования, однако достаточно определенные выводы о локализации и тем более относительной роли различных популяций-рефугиумов в формировании генофонда современных популяций этого вида в ледниковой зоне ареала пока не сделаны [1–8]. Одни авторы предполагают существование ледниковых убежищ сосны обыкновенной на Балканах [3, 7, 9], другие – на западе Альп [4], третьи – на Южном Урале [10, 11]. При этом конкретные геногеографические параметры пока приведены лишь по Балканскому и Южно-Уральскому рефугиумам [11].
К давним дискуссионным вопросам проблемы поиска и выявления плейстоценовых рефугиумов относится сохранность рефугиумов P. sylvestris в некоторых крайне северных (перигляциальных) регионах внеледниковой зоны. Существование таких ледниковых убежищ, из которых происходила приоритетная реколонизация популяций в голоцене, предполагается на юго-западе Ирландии [1, 12], в восточной Прибалтике [5, 6, 13] и даже на Лофотенских островах, отепляемых палео-Гольфстримом [14]. Гипотетичные рефугиумы популяций Скандинавии выявлены также в Чехии и Словакии [9]. Однако пока все эти гипотезы еще недостаточно подтверждены количественным геногеографическим анализом.
Решению проблемы, на наш взгляд, препятствует прежде всего отсутствие достаточно репрезентативной информации о геногеографической структуре вида и ее взаимосвязях в пределах всего ареала. Кроме того, в России недостаточны палеоботаническая изученность ареала P. sylvestris и число полиморфных маркеров ДНК для выявления географической изменчивости вида, особенно в Сибири [5, 15–17].
На данном этапе исследований пока более информативным в плане выявления рефугиумов популяций P. sylvestris оказался их тотально-видовой (свыше 250 выборок) аллозимный анализ на достаточно густой и равномерной сети трансект, пересекающих весь ареал вида, т.е. на основе системного подхода “вид в целом ареале” [18, 19]. Ранее нами был проведен [11] сравнительный анализ “минимальных генетических дистанций” Неи, отражающих генетическое сходство между популяциями северной (ледниковой) зоны ареала и 12 маргинальными популяциями в его южной части. Выявлены два наиболее вероятных гипотетичных плейстоценовых рефугиума (ГПР) – Балканский и Южно-Уральский и четыре второстепенных – Тургайский, Алтайский, Южно-Прибайкальский и Тувинский [11]. Однако в трансконтинентальной полирефугиальной системе вида Pinus sylvestris L., кроме названных убежищ выживания, вероятно, существовали и некоторые другие. Например, нам [20] удалось выявить ГПР для популяций в крайней северо-восточной части ареала в Якутии.
Цель настоящей работы – анализ и обобщение результатов широкомасштабного поиска, выявления и сравнительной оценки вклада в формирование генофонда популяций P. sylvestris ледниковой зоны системы наиболее вероятных ГПР, расположенных не только в маргинальной южной, но и в северной (перигляциальной) части внеледниковой зоны последней фазы оледенения.
ОБЪЕКТЫ И МЕТОДЫ
Для выявления гипотетических плейстоценовых рефугиумов вида Pinus sylvestris L. использованы следующие основные методические подходы:
1. Тотальный геногеографический анализ аллозимной структуры популяций вида на относительно равномерной сети трансект, пересекающих весь ареал вида (“вид в целом ареале”).
2. Выявление и сопоставимая оценка генетических дистанций популяций P. sylvestris ледниковой зоны и их групп с общевидовой системой ГПР южной маргинальной части ареала.
3. Подразделение системы ГПР по их географической локализации и потенциальной роли в формировании генофонда ледниковой зоны на две группы – маргинальные южные (“анцестральные”) и перигляциальные (“авангардные”).
Исследования проведены на двух сериях объектов, охватывающих весь ареал P. sylvestris (рис. 1, табл. 1). Первая серия включает 26 популяций, расположенных во внеледниковой части ареала последней ледниковой фазы. Из них 13 популяционных выборок расположены вдоль южной границы ареала и отнесены к анцестральным – Мадрид, Бергамо, Рила, Тауз, Крыктытау, Наурзум, Актогай, Усть-Каменогорск, Кызыл, Кяхта, Нижний Цасучей, Архара, Комсомольск-на-Амуре, а остальные 13 – в северной перигляциальной части внеледниковой зоны. По данным палинологии [4, 12, 13, 15, 16, 21–23] они сохранялись и существовали не позднее 12–13 тыс. лет ВР, т.е. до начала их реколонизации на север в голоцене. К этим авангардным ГПР отнесены Пиренеи, Бавария, Шумава (Чехия), Низкие Татры (Словакия), Карпаты, Крым, Архыз, Иремель, Тахтаброд, Телецкое озеро, Иркутск, Романовка, Тында.
Рис. 1.
Картосхема географического распределения гипотетических анцестральных и авангардных плейстоценовых рефугиумов (ГПР) Pinus sylvestris L. и выявление среди них основных (DN78 = 0–0.008) рефугиумов в пределах всего ареала вида: 1 – граница ареала P. sylvestris L.; 2 – границы максимального покровного оледенения; 3 – южные границы распространения островной реликтовой почвенной мерзлоты на территории Западной Сибири и Европы; 4 – ориентировочные южные границы перигляциальной зоны на территории Восточной Сибири; 5 – анцестральные ГПР; 6 – авангардные ГПР; 7 – основные ГПР для восьми филогеографических регионов; 8 – границы ФГП: Шт, ЦЕ, Ск, Рр, Ур, ЗС, СС, Як (см. табл. 1).
Шифры популяций: Аз – Архыз, Ак – Актогай, Ах – Архара, Бв – Бавария, Бг – Бергамо, Ик – Иркутск, Ир – Иремель, Кз – Кызыл, Км – Комсомольск-на-Амуре, Кт – Крыктытау, Кх – Кяхта, Мд – Мадрид, Мк – Микуличин, Нз – Наурзум, НЦ – Нижний Цасучей, Пр – Пиренеи, РК – Роман-Кош, Рл – Рила, Рм – Романовка, Сл – Словакия, Тз – Тауз, Тн – Тында, ТО – Телецкое озеро, Тх – Тахтоброд, УК – Усть-Каменогорск, Чх – Чехия.
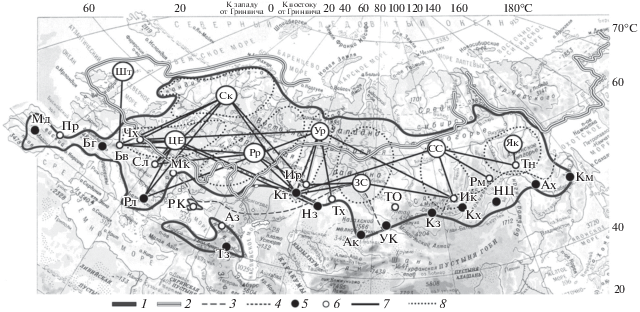
Таблица 1.
Названия и шифры ФГП ледниковой зоны ареала и популяций внеледниковой зоны ареала
| ФГП | Шифр | Ледниковая зона ареала | ||
|---|---|---|---|---|
| Географическое распределение популяций ледниковой зоны по ФГП |
||||
| Шотландия | Шт | Шотландия | ||
| Центральная Европа | ЦЕ | Брест, Калининград, Гомель, Минск, Западная Двина, Псков | ||
| Скандинавия | Ск | Выборг, Осло, Кемь, Петрозаводск, Мурманск, Упсала, Лунд, Колари | ||
| Русская равнина | Рр | Архангельск, Мезень, Вологда, Москва, Воронеж, Сыктывкар, Киев | ||
| Урал | Ур | Нижний Тагил, Новая Ляля, Ивдель, Печора, Карпинск, Печеро-Илыч, Красновишерск, Соликамск | ||
| Западная Сибирь | ЗС | Геологическое, Надым, Заводоуспенское, Сургут, Ишим, Тарко-Сале, Междуреченск, Ханты-Мансийск | ||
| Средняя Сибирь | СС | Братск, Кодинск, Ванавара, Северобайкальск, Зима, Тура, Качуг, Туруханск | ||
| Якутия | Як | Мирный, Якутск, Витим, Алдан, Вилюйск, Хандыга, Олекминск | ||
| Внеледниковая зона ареала | ||||
| Популяция | Шифр | Популяция | Шифр | |
| Архыз | Аз* | Наурзум | Нз | |
| Актогай | Ак | Нижний Цасучей | НЦ | |
| Архара | Ах | Пиренеи | Пр* | |
| Бавария | Бв* | Роман-Кош | РК* | |
| Бергамо | Бг | Рила | Рл | |
| Иркутск | Ик* | Романовка | Рм* | |
| Иремель | Ир* | Словакия | Сл* | |
| Кызыл | Кз | Тауз | Тз | |
| Комсомольск-на-Амуре | Км | Тында | Тн* | |
| Крыктытау | Кт | Телецкое озеро | ТО* | |
| Кяхта | Кх | Тахтоброд | Тх* | |
| Мадрид | Мд | Усть-Каменогорск | УК | |
| Микуличин | Мк* | Чехия | Чх* | |
Во вторую серию включена 51 популяционная выборка из 8 филогеографических групп популяций (ФГП), по 7–8 выборок в каждой, за исключением Шотландии (одна выборка), расположенных в различных ландшафтных странах ледниковой части ареала (Британские острова, Центральная Европа, Скандинавия, Русская равнина, Урал, Западная Сибирь, Средняя Сибирь, Якутия).
Основной методический подход системного геногеографического анализа в пределах всего ареала вида заключается в сравнительном иерархическом анализе средних генетических дистанций Неи [24] между 26 названными выше популяциями ГПР внеледниковой зоны и 8 филогеографическими группами популяций (ФГП) ледниковой зоны.
Аллозимный анализ в общей сложности 77 популяционных выборок (по 30–48 деревьев в каждой, всего около 3500 особей) выполнен с помощью общепринятых методов [25] с некоторыми модификациями [26] по 16 белок-кодирующим локусам, в том числе 14 полиморфным, десяти ферментных систем (GOT, 6-PGDH, SKDH, GDH, ADH, FDH, PGM, DIA, EST-f, SOD). Генетические дистанции Неи [24] вычислены по программам BYOSYS [27], а их кластерный анализ проведен по методу невзвешенных попарногрупповых средних (УПГМА) по программам NTSYS [28].
Критерием генетического сходства предполагаемого рефугиума с генофондом той или иной северной группы популяций служила минимальная величина средней генетической дистанции Неи [24] менее 0.015 между ними. Если по нашей геносистематической шкале [18] DN78 не превышает субпопуляционный уровень (0.001–0.008), то минимальные генетические дистанции и соответствующие рефугиумы отнесены к I субпопуляционному рангу сходства, а при DN78 от 0.008 до 0.015 – ко II популяционному рангу.
Для оценки вклада популяции того или иного рефугиума в генофонд северных групп популяций применен эмпирический “инвестиционный индекс рефугиума” Iirf = 1/(DN78 × 100) [11]. Он позволяет намного контрастнее выявить различия относительного вклада различных рефугиумов в формирование генетического пула той или иной группы или отдельной популяции. При этом общий вклад всех изучавшихся ГПР в генофонд данной ФГП может быть определен как сумма их Iirf, а вклад каждого ГПР в генофонд этой ФГП – как доля (%) их интегрального вклада.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Анализ минимальных генетических дистанций. Географический анализ минимальных DN78 I (≤0.008) и II (от 0.008 до 0.015) ранга между популяциями ФГП в направлении от Шотландии к Якутии позволяет на двух уровнях их сходства с популяциями ГПР выявить локализацию основных и второстепенных гипотетичных рефугиумов в пределах всего ареала P. sylvestris.
Для региона Шотландии выявляется лишь один ГПР I ранга (Бавария, DN78 = 0.007) и несколько минимальных генетических дистанций на уровне II ранга сходства (DN78 = 0.009–0.015) с избранной серией 26 ГПР, в том числе на Чешско-Моравской возвышенности (рис. 2).
Рис. 2.
Распределение популяций внеледниковой зоны ареала P. sylvestris по рангам генетических дистанций Неи (DN78) с различными филогеографическими группами популяций ледниковой зоны: Шт – Шотландия, ЦЕ – Центральная Европа, Рр – Русская равнина, Ск – Скандинавия; I – ГПР первого ранга (DN78 ≤ 0.008), II – ГПР второго ранга (DN78 = 0.008–0.015). Шифры популяций см. в табл. 1.
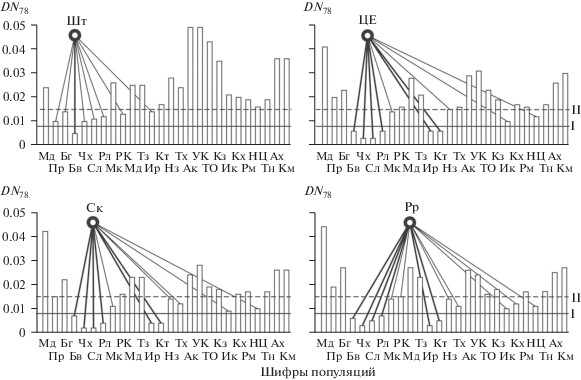
В ФГП Центральной Европы (куда включены выборки из Баварии, Чехии, Словакии, Калининграда, Пскова, Минска и Бреста, см. рис. 2) максимальное сходство выявлено с двумя географически смежными авангардными ГПР – в Чехии (DN78 = 0.003) и Словакии (Низкие Татры, 0.004), а несколько меньшее – с маргинальным южным (вероятно, анцестральным) в Балканах (Рила, 0.005). Не менее отчетливое, чем с Балканским рефугиумом, сходство центрально-европейской группы популяций P. sylvestris с двумя ГПР Южного Урала – маргинальным южным Крыктытау (0.005) и расположенным на 200 км севернее Иремель (0.005). Кроме того, на уровне II ранга найдены минимальные дистанции DN78 с ФГП Центральной Европы у Карпатского (Микуличин, 0.013), двух центрально-казахстанских (Наурзум, Тахтаброд, 0.014–0.015) и даже у двух сибирских – южно-прибайкальского (Иркутск, 0.010) и даурского (Нижний Цасучей, 0.011) рефугиумов. Вероятно, это отражает не только плейстоценовые, но и анцестральные, миоцен-плиоценовые генеалогические связи.
Почти аналогичные с Центральной Европой системы и соотношения ГПР характерны (см. рис. 2) для ФГП Скандинавии (Мурманск, Колари, Кемь, Осло, Упсала, Лунд, Петрозаводск, Выборг) и Русской равнины (Архангельск, Мезень, Сыктывкар, Вологда, Москва, Гомель, Воронеж, Киев). Обнаружен также крайне юго-западный пиренейский рефугиум II ранга для Скандинавии. В целом же здесь так же, как и в Центральной Европе, отчетливо выявляется сходство с основными южно-европейскими (Чехия, Словакия, Балканы), южно-уральскими (Крыктытау, Иремель) и, как ни парадоксально, с аддитивными сибирскими рефугиумами (Иркутск и даже Нижний Цасучей). Эти трансконтинентальные генетические связи во многом подтверждают гипотезу восточно-азиатского происхождения вида P. sylvestris L. [19, 29]. По-видимому, аллозимное наследование отражает не только (и не столько) недавние плейстоценовые связи популяций P. sylvestris в пределах ареала, но и кумулятивные анцестральные с самого начала расселения вида.
Далее, начиная с ФГП Урала (Печора, Печоро-Илычский заповедник, Соликамск, Ивдель, Павда, Исеть, рис. 3), формируется уже близкий к симметричному пучок направлений аллозимного сходства популяций с теми же рефугиумами, что и в Европе. Прежде всего это характерно для двух южно-уральских (Крыктытау, Иремель) горных популяций. Ослабляются связи со Словакией и появляются с казахстанским (Тахтаброд) и южно-прибайкальским (Иркутск) основными, а также с четырьмя второстепенными – южно-сибирскими (Телецкое озеро, Кызыл) и забайкальскими (Романовка, Нижний Цасучей) рефугиумами.
Рис. 3.
Распределение популяций внеледниковой зоны ареала P. sylvestris по рангам генетических дистанций Неи (DN78) с различными филогеографическими группами популяций ледниковой зоны: Ур – Урал, ЗС – Западная Сибирь, СС – Средняя Сибирь, Як – Якутия; I – ГПР первого ранга (DN78 ≤ 0.008), II – ГПР второго ранга (DN78 = = 0.008–0.015). Шифры популяций см. в табл. 1.
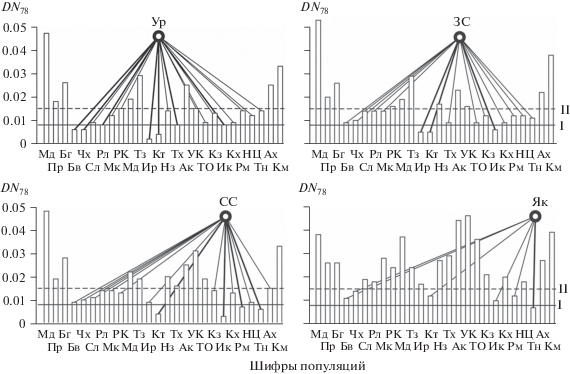
В Западной Сибири (см. рис. 3) европейские рефугиумы уже отчетливо утрачивают роль лидирующих. Пучок направлений минимальных генетических дистанций на основные рефугиумы – два южно-уральских и южно-прибайкальский, а также на серии второстепенных становится почти строго симметричным. В соответствии с гипотезой гидрохории хвойных [30] и результатами аллозимных исследований в бассейне р. Обь [31] можно предположить, что влияние южно-прибайкальского рефугиума на генофонд популяций Урала связано с гидрохорией семян сосны по р. Ангаре, а далее – после преодоления ими водораздела с Обью – и на северо-запад Западной Сибири.
В Средней Сибири (см. рис. 3), где наряду с южно-прибайкальским (Иркутск) впервые выявлена доминирующая роль двух забайкальских рефугиумов – Романовки и Тынды, формируется асимметричный восточный спектр рефугиумов. Еще более явно это выражено в Якутии, для генофонда которой удалось выявить лишь один основной рефугиум (Тында, DN78 = 0.007) в верховьях р. Олекмы, пересекающей Становой хребет. Здесь также подтверждается приоритетная роль фактора гидрохории семян сосны в голоценовой реколонизации ее популяций.
Общая картина локализации основных гипотетичных рефугиумов и линий их максимального сходства с группами популяций ледниковой зоны ареала P. sylvestris, отражающих направления их постгляциальной реколонизации, свидетельствует (см. рис. 1) о полирефугиальном происхождении и весьма широком разностороннем обмене генами в пределах всего ареала не только в плейстоцене, но, по-видимому, и в течение многих миллионов лет существования вида в Северной Евразии.
В целом по ареалу минимальные генетические дистанции I ранга (на уровне DN78 < 0.008) с северными группами популяций выявлены у девяти рефугиумов (табл. 2). Они представлены двумя маргинальными южными (наиболее вероятно, анцестральными третичными – балканским (Рила) и южно-уральским (Крыктытау)) и семью авангардными, форпосты которых расположены на несколько сотен километров севернее (Чехия, Словакия, Иремель, Тахтаброд, Иркутск, Романовка, Тында).
Таблица 2.
Формулы относительного вклада (%) инвестиционных индексов генофонда различных популяций внеледниковой зоны в генофонд ФГП P. silvestris L. ледниковой зоны ареала
| ФГП | Индексы инвестиции генофонда Iirf (1/(DN78×100)) | Сумма | Доминирующие рефугиумы, % |
|---|---|---|---|
| Шт | *2.0Бв **1.0Пр 1.0Чх 0.9Сл 0.8Рл 0.8РК 0.7Бг 0.7Ир 0.6НЦ 0.6Кт 0.5Рм 0.5Тн 0.5Кх 0.5Ик 0.4Мд 0.4Тх 0.4Аз 0.4Тз 0.4Мк 0.4Нз 0.3Кз 0.3Ах 0.2Км 0.2TO 0.2Ак 0.2УК |
15.0 | *13.3Бв **6.7Пр 6.7Чх 6.1Сл 5.6Рл 5.1РК 4.8Бг 4.8Ир |
| ЦЕ | *3.3Чх 3.3Сл 1.7Бв 1.7Рл 1.7Ир 1.7Кт **1.0Ик 0.8НЦ 0.7Мк 0.7Нз 0.6РК 0.6Тх 0.6Рм 0.6Кх 0.6Тн 0.5Кз 0.5Пр 0.5Тз 0.4Бг 0.4TO 0.4Ах 0.4Аз 0.3Ак 0.3Км 0.3УК 0.2Мд |
24.0 | *19.3Чх 19.3Сл 7Бв 7Рл 7Ир 7Кт **4.2Ик 3.5НЦ 3.0Мк 2.8Нз |
| Ск | *5.0Чх 5.0Сл 2.5Рл 2.5Ир 2.5Кт 1.4Бв **1.1Ик 1.0НЦ 0.9Мк 0.8Тх 0.7Нз 0.7Пр 0.6РК 0.6Кх 0.6Рм 0.6Тн 0.6Кз 0.5TO 0.5Бг 0.4Аз 0.4Тз 0.4Ак 0.4Ах 0.4Км 0.4УК 0.2Мд |
30.8 | *16.3Чх 16.3Сл 8.1Рл 8.1Ир 8.1Кт 4.6Бв **3.6Ик 3.3НЦ 3.0Мк 2.7Тх 2.3Нз 2.2Пр |
| Рр | *3.3Чх 3.3Ир 2.0Сл 2.0Кт 1.7Бв 1.4Рл **1.0Ик 0.9Тх 0.9НЦ 0.8Кх 0.7Мк 0.7Нз 0.7РК 0.6TO 0.6Рм 0.6Тн 0.6Кз 0.5Пр 0.4Тз 0.4УК 0.4Ах 0.4Ак 0.4Бг 0.4Аз 0.4Км 0.2Мд |
25.4 | *13.1Чх 13.1Ир 7.9Сл 7.9Кт 6.6Бв 5.6Рл **3.9Ик 3.6Тх 3.6НЦ 3.3Кх 2.8Мк 2.8Нз 2.6РК |
| Ур | *5.0Ир 2.5Кт 1.7Бв 1.7Чх 1.3Рл 1.3Тх 1.3Ик **1.1Сл 1.1TO 1.1Кх 0.8Мк 0.8НЦ 0.8Кз 0.7Нз
0.7Рм 0.7Тн 0.7РК 0.7УК 0.6Пр 0.5Аз 0.4Ак 0.4Ах 0.4Бг 0.3Тз 0.3Км 0.2Мд |
27.0 | *18.5Ир 9.3Кт 6.2Бв 6.2Чх 4.6Рл 4.6Тх 4.6Ик **4.1Сл 4.1TO 4.1Кх 3.1Мк 3.1НЦ 2.9Кз 2.6Нз 2.6Рм 2.6Тн 2.5РК 2.5УК |
| ЗС | *2.0Ир 2.0Кт 1.7Ик **1.1Бв 1.1Тх 1.1TO 1.1Кх 1.0Чх 0.9Тн 0.8Кз 0.8Рм 0.8НЦ 0.7Сл 0.7Рл 0.7Мк 0.6РК 0.6УК 0.6Нз 0.5Аз 0.5Пр 0.5Ах 0.4Ак 0.4Бг 0.3Тз 0.3Км 0.2Мд |
21.6 | *9.3Ир 9.3Кт 7.7Ик **5.1Бв 5.1Тх 5.1TO 5.1Кх 4.6Чх 4.2Тн 3.9Кз 3.9Рм 3.9НЦ 3.3Сл 3.3Рл 3.3Мк |
| СС | *3.3Ик 2.5Кт 1.7Тн 1.4Рм **1.1Бв 1.1Ир 1.1НЦ 1.0Чх 0.9Сл 0.8РК 0.8Кх 0.7Рл 0.7Мк 0.7Кз 0.7Ах 0.6Тх 0.5Пр 0.5Тз 0.5TO 0.5Нз 0.5Аз 0.4Ак 0.4Бг 0.3УК 0.3Км 0.2Мд |
23.3 | *14.3Ик 10.7Кт 7.2Тн 6.1Рм **4.8Бв 4.8Ир 4.8НЦ 4.3Чх 3.9Сл 3.3РК 3.3Кх 3.1Рл 3.1Мк 3.1Кз 2.9Ах |
| Як | *1.4Тн **1.0Ик 0.9Бв 0.8Кт 0.8Рм 0.7Чх 0.6Ир 0.6Рл 0.6НЦ 0.5Сл 0.5Кх 0.5Кз 0.4РК 0.4Тз 0.4Пр 0.4Бг 0.4Нз 0.4Ах 0.4Мк 0.3Тх 0.3TO 0.3Аз 0.3Мд 0.3Км 0.2Ак 0.2УК |
13.5 | *10.6Тн **7.4Ик 6.7Бв 6.2Кт 6.2Рм 5.3Чх |
Примечание. Полужирным выделены: * – популяции ГПР I ранга (DN78 ≤ 0.008), ** – популяции ГПР II ранга (DN78 = 0.008–0.015). Шифры популяций см. в табл. 1.
Менее значимую аддитивную донорскую роль (на уровне DN78 = 0.008–0.015) в ходе формирования современных популяций ледниковой зоны играли ГПР II ранга: Пиренейский и Карпатский – для Центральной Европы и Скандинавии, Тургайский – для Урала, Алтайский, Тувинский, Северо-Монгольский (Кяхта), Даурский (Нижний Цасучей) и Архара – для Урала и Сибири.
Анализ и оценка инвестиционных индексов рефугиумов. Относительный вклад (формулу) различных плейстоценовых рефугиумов в формирование генетического пула отдельных филогеногеографических групп популяций ледниковой зоны ареала наиболее отчетливо отражают доли их инвестиционных индексов Iirf = 1/(DN78 × 100) (рис. 4, табл. 2). Для каждой ФГП в направлении с запада на восток ареала они колеблются в пределах от 2 до 18% (от общей суммы в ФГП) в зависимости от удаленности ближайших популяций-ГПР и степени их миграционной изоляции (дистанционной, горной и гидрохорной).
Рис. 4.
Диаграммы долей вклада (% от общей суммы вкладов 1/DN78 всех ГПР) генофонда ГПР в генофонд ФГП Шотландии (Шт), Центральной Европы (ЦЕ), Скандинавии (Ск), Русской Равнины (Рр), Урала (Ур), Западной Сибири (ЗС), Средней Сибири (СС) и Якутии (Як). Шифры популяций см. в табл. 1.
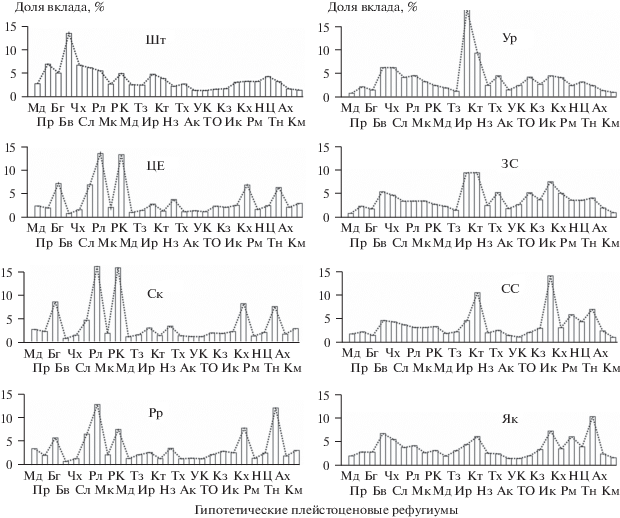
Соотношения параметров вклада различных ГПР в генофонд филогеографических групп популяций ледниковой зоны Северной Евразии, приведенные в табл. 2, отражают состав их “генетических сертификатов” по местонахождению рефугиумов происхождения. Графически они могут быть представлены в виде спектров индексов Iirf, которые являются зеркальным отражением спектров минимальных DN78, показанных на рис. 2 и 3. В принципе этот общий методический подход позволяет выполнить генетическую сертификацию любой популяции по данным ее аллозимного (или в перспективе ДНК) анализа и установленных инвестиционных индексов ГПР.
ЗАКЛЮЧЕНИЕ
Полицентричность географической локализации и относительный вклад популяций гипотетичных плейстоценовых рефугиумов (ГПР) P. sylvestris в формирование генофонда популяций ледниковой зоны ареала в первом приближении выявлены на основе аллозимного анализа в пределах всего ареала вида по минимальным генетическим дистанциям Неи (DN78) между ними и их обратным величинам – “инвестиционным индексам” (Iirf = 1/(DN78 × 100)).
Средние по ареалу минимальные генетические дистанции между популяциями ГПР (DN78 < 0.008), их максимальные инвестиционные индексы по отношению к популяциям P. sylvestris ледниковой зоны и вероятная доминирующая роль в формировании их генофонда выявлены на основе иерархического аллозимного анализа у четырех гипотетичных рефугиумов внеледниковой зоны ареала в Европе (Чехия, Словакия, Балканы и северные Альпы), двух – на Южном Урале (Иремель, Крыктытау) и двух – в Южном Прибайкалье (Иркутск) и Приамурье (Тында).
Меньшую (аддитивную) роль в формировании генетического пула популяций ледниковой зоны (на уровне минимальных DN78 = 0.008–0.014), вероятно, играли Карпатский, два Центрально-Казахстанских (Наурзум, Тахтаброд), три – из региона гор Южной Сибири (Телецкое озеро, Балгазын, Кяхта) и два – из Забайкалья (Романовка, Нижний Цасучей). Остальные ГПР, особенно расположенные на крайнем юго-западе (Иберийский и Южно-Альпийский) и юго-востоке (Нижне-Амурский) ареала, изолированные от его центральных регионов горными, морскими или степными барьерами, оказали незначительное влияние на генетический пул популяций ледниковой зоны.
Формулы относительного вклада 26 тестированных ГПР внеледниковой зоны ареала в формирование генофонда 8 ФГП ледниковой зоны, вычисленные по эмпирическим инвестиционным индексам, могут служить генетическими сертификатами современных популяций по месту их происхождения. По географическому положению все плейстоценовые рефугиумы можно подразделить на две группы: 1) маргинальные южные (анцестральные), сохранявшиеся в эпоху максимального оледенения, и 2) перигляциальные (авангардные), характеризующиеся различным инвестиционным вкладом в формирование генофонда популяций P. sylvestris ледниковой зоны ареала.
Работа выполнена в рамках государственного задания Федерального государственного бюджетного учреждения науки Ботанический сад УрО РАН и при поддержке РФФИ (проект № 16-04-00948). Авторы выражают благодарность профессору Л. Пауле (Зволенский технологический университет, Словакия) за предоставление образцов сосны из Центральной Европы.
Авторы подтверждают отсутствие конфликта интересов.
Список литературы
Bennett K.D., Tzedaki P.C., Willis K.J. Quarernary refugia of north European trees // J. of Biogeography. 1991. V. 18. P. 103–115.
Sinclair W.T., Morman J.D., Ennos R.A. The postglacial history of Scots pine (Pinus sylvestris L.) in western Europe: evidence from mitochondrial DNA variation // Molecular Ecology. 1999. № 8. P. 83–88.
Soranzo N., Alia R., Provan J. et al. Patterns of variation at mitochondrial sequence tagged-site locus provides new insights into the postglacial history of European Pinus sylvestris populations // Molecular Ecology. 2000. V. 9. P. 1205–1211.
Cheddadi R., Vendramin G.G., Litt T. et al. Imprints of glacial refugia in the modern genetic diversity of Pinus sylvestris // Global Ecol. Biogeogr. 2006. V. 15. P. 271–282.
Naidenov K., Senneville S., Beaulieu J. et al. Glacial variance in Eurasia: mitochondrial DNA evidence from Scots pine for a complex heritage involving genetically distinct refugia at mid-northern latitudes and in Asia minor // BioMed Central Evolutionary Biology. 2007. V. 7. http://creative commons.crd/lictnses/by/2.0
Pihajarvi T., Salmela M.J., Savolainen O. Colonization routes of Pinus sylvestris inferred from distribution of mito- chondrial DNA variation // Tree Genetics and Genomes. 2008. V. 4. P. 247–254.
Celepirovic N., Ivankovic M., Gradecki-Postenjak et al. Review of investigation of variability of nad 1 gene introne B/C of mitochondrial genome in Scots pine (Pinus sylvestris L.) // Periodicum Biologorum. 2009. V. 111. № 4. P. 453–457.
Dering M., Kosiński P., Wyka T.P. et al. Tertiary remnants and Holocene colonizers: genetic structure and phylogegraphy of Scots pine reveal higher genetic diversity in young boreal than in relict Mediterranian populations and a dual colonization of Fennoscandia // Diversity and Distributions. 2017. V. 23. P. 540–555.
Sannikov S.N., Paule L., Yegorov E.V. Identification of probable Pleistocene refugia of Pinus sylvestris L. in Europe // Forest Genetics. 2013. V. 13(4). P. 201–206.
Филиппова Т.В., Санников С.Н., Петрова И.В. и др. Феногеногеография популяций сосны обыкновенной на Урале. Екатеринбург: УрО РАН, 2006. 122 с.
Санников С.Н., Петрова И.В., Егоров Е.В. и др. Выявление системы плейстоценовых рефугиумов Pinus sylvestris L. в южной маргинальной зоне ареала // Экология. 2014. № 3. С. 174–181. [Sannikov S.N., Petrova I.V., Egorov E.V. et al. A system of pleistocene refugia for Pinus sylvestris L. in the southern marginal part of the species range // Rus. J. of Ecology. 2014. V. 45. № 3. P. 167–173].
Huntley B., Birks H.J. An atlas of past and pollen maps for Europe: 0–13 000 years ago. Cambridge: Cambridge Univ. Press, 1983. 667 p.
Lang G. Quartare Vegetationsgeschichte Europas: Methoden und Ergebnisse. Jena, Stuttgard, New York: Gustav Fischer Verlag, 1994. 462 S.
Санников С.Н., Санникова Н.С., Петрова И.В. и др. Гипотеза о Лофотенском плейстоценовом рефугиуме Pinus sylvestris L. // Экология. 2019. № 3. С. 172–181.
Kremenetski C.V., Kam-biu-Liu, MacDonald G.M. The late Quaternary dynamics of pines in Northern Asia // Ecology and biogeography of Pinus. N.Y.; Melbourne; Madrid; Cambridge: Cambridge Univ. Press, 1998. P. 95–106.
Бляхарчук Т.А. Новые палеопалинологические данные о динамике растительного покрова и климата Западной Сибири и прилегающих территорий в голоцене. Новосибирск: Академическое изд-во “Гео”, 2012. 138 с.
Семериков В.Л., Путинцева Ю.А., Орешкова Н.В. и др. Разработка новых маркеров митохондриальной ДНК сосны обыкновенной (Pinus sylvestris L.) для популяционно-генетических и филогеографических исследований // Генетика. 2015. Т. 51. № 12. С. 1386–1390.
Санников С.Н., Петрова И.В. Филогеногеография и генотаксономия популяций вида Pinus sylvestris L. // Экология. 2012. № 4. С. 252–260.
Санников С.Н., Санникова Н.С., Петрова И.В. Очерки по теории лесной популяционной биологии. Екатеринбург: УрО РАН, 2012. 270 с.
Санников С.Н., Петрова И.В., Санникова Н.С. и др. Дивергенция биогеоценозов в пределах типов сосновых лесов // Экология. 2017. № 4. С. 282–291.
Хотинский Н.А. Голоцен Северной Евразии. Опыт трансконтинентальной корреляции этапов развития растительности и климата. М.: Наука, 1977. 200 с.
Никифорова Л.Д. Изменение природной среды в голоцене на северо-востоке европейской части СССР: Автореф. дис. … канд. геол.-минерал. наук. М., 1980. 25 с.
Бляхарчук Т.А. Послеледниковая динамика растительного покрова Западно-Сибирской равнины и Алтае-Саянской горной области (по данным спорово-пыльцевого анализа болотных и озерных отложений): Автореф. дис. … докт. биол. наук. Томск, 2010. 43 с.
Nei M. Estimation of average heterozygosity and genetic distance from a small number of individuals // Genetics. 1978. V. 89. P. 583–590.
Корочкин Л.И., Серов О.Л., Пудовкин А.И. и др. Генетика изоферментов. М.: Наука, 1977. 275 с.
Семериков В.Л., Подогас А.В., Шурхал А.В. и др. Электрофоретическая изменчивость белков хвои сосны обыкновенной Pinus sylvestris L. // Генетика. 1991. Т. 27. № 9. С.1590–1596.
Swofford D.L., Selander R.B. BIOSYS-1: a FORTRAN program for the comprehensive analysis of electrophoretic data in population genetics and systematics // Heredity. 1981. V. 72. P. 281–283.
Rohlf E.J. Numerical taxonomy and multivariate analysis system. New York: Exter Publishing LTD, 1988. 16 p.
Санников С.Н., Петрова И.В. Дифференциация популяций сосны обыкновенной. Екатеринбург: УрО РАН, 2003. 248 с.
Санников С.Н., Санникова Н.С. Эскиз теории гидрохории некоторых видов хвойных // Докл. РАН. 2008. Т. 418. № 6. С. 847–849.
Санникова Н.С., Егоров Е.В. Гипотеза генетической интеграции популяций Pinus sylvestris L. в долине р. Обь // Сибирский лесной журн. 2016. № 5. С. 107–113.
Дополнительные материалы отсутствуют.