

Экология, 2020, № 4, стр. 243-250
Индексы листовой поверхности древостоев в естественных и нарушенных лесах Приморского края
А. В. Иванов a, b, *, В. Ю. Покаместова a, А. С. Касаткин c, Д. Г. Замолодчиков d
a Приморская государственная сельскохозяйственная академия
692510 Уссурийск, ул. Блюхера, 44, Россия
b Дальневосточный НИИ лесного хозяйства
680020 Хабаровск, ул. Волочаевская, 71, Россия
c Рослесозащита
141207 Московская область, Пушкино, ул. Надсоновская, 13, Россия
d Центр по проблемам экологии и продуктивности лесов РАН
117234 Москва, ул. Профсоюзная, 84/32, Россия
* E-mail: aleksandrgg86@mail.ru
Поступила в редакцию 24.07.2019
После доработки 14.11.2019
Принята к публикации 30.12.2019
Аннотация
Определены индексы листовой поверхности (LAI) древостоев в естественных и нарушенных рубками и пожарами лесах Приморского края на основе сведений по фитомассе листвы и удельной листовой поверхности (SLA). Исходными данными послужили материалы инвентаризации 44 пробных площадей. LAI древостоев, нарушенных вследствие пожаров и рубок, более чем на 30% ниже, чем у малонарушенных лесов, близких по структуре к коренным хвойно-широколиственным насаждениям. Индекс листовой поверхности значимо связан с запасом древесины насаждений (R2 = 0.68), однако его следует считать самостоятельным, независимым показателем, расчет которого на основе таксационных характеристик древостоя связан с высокими неопределенностями.
В XXI в. экосистемные функции лесов, не связанные с производством древесины, становятся объектом экономических расчетов и учитываются при политических решениях. Постепенно индустриальная цивилизация начинает переосмысливать значение лесов, осознавая, что срубленное дерево может приносить меньше пользы, чем оставленное на корню. Наиболее важные в глобальном масштабе экосистемные функции лесов связаны с поддержанием циклов вещества и энергии в системе суша–атмосфера: водного баланса, включая регулирование стока и перераспределение осадков, круговоротов углерода, азота и других элементов [1, 2]. С позиции традиционного “древесинного” лесопользования лесная экосистема рассматривается прежде всего как совокупность стволов деревьев разной сортиментно-сортной структуры. Например, система лесопользования России преимущественно направлена на решение проблемы получения максимума древесины при минимуме усилий. Оценкой лесных насаждений как государственного древесного ресурса занимается специальная наука – лесная таксация. Подробные таксационные характеристики в длительной динамике представлены для всех лесов России по субъектам РФ и лесничествам, однако их использование для оценки экосистемных услуг лесов затруднительно. Накопленный объем новых знаний о лесах, базы данных об их продуктивности, биомассе, биологическом разнообразии, пространственной структуре, листовых индексах, потоках углерода, фенологическом развитии и др. формирует новое – экологическое – направление в таксации лесов [3].
Важным компонентом экологической таксации следует признать характеристику листовой поверхности древесного полога. Индекс листовой поверхности LAI (leaf area index) определяется как отношение площади поверхности листьев фитоценоза или его частей к площади территории, занимаемой этим фитоценозом. Обычно LAI измеряется в м2/м2 или в га/га, либо указывается как безразмерная величина [4]. Для лесов, как правило, приводятся оценки LAI древостоя без учета подчиненных ярусов. Согласно обзору В.И. Грабовского с соавт. [5], среднее значение LAI для лесов России по состоянию на июль находится в пределах 4–6.
Через поверхность листвы древесного полога происходит преобразование радиационной энергии в биомассу, обмен водой и потоками углерода между растениями и окружающей средой [6]. Таким образом, LAI может выступать одним из предикторов при определении продуктивности лесов и фактором в моделях экофизиологических процессов как на локальном, так и на глобальном уровнях [4, 6–9]. Установлена связь между валовой продукцией фотосинтеза (GPP) и значением LAI [10]. Индекс листовой поверхности может служить характеристикой степени деградации лесов после выборочных рубок и иных структурных нарушений в лесных насаждениях. Так, выполненное в буковых лесах Словакии исследование показало [6], что LAI вторичных лесов оказался на 1.6 ед. меньше, чем у коренных ненарушенных фитоценозов бука. У лесных насаждений LAI обычно находится в диапазоне 1–20 [11]. Расходы воды на транспирацию в пологе лесного насаждения на 90% определяются индексом листовой поверхности [4]. В работе “Площадь поверхности лесных растений” А.И. Уткин писал: “Вряд ли будет преувеличением утверждать, что уже в ближайшие годы LAI станет одним из важнейших таксационных показателей, таких как абсолютная полнота, класс бонитета, годичный прирост. Но для этого необходима фактологическая перевооруженность информационного обеспечения лесной науки, прежде всего за счет целеустремленного сбора данных о LAI для древостоев разных древесных пород с учетом их возрастной динамики, смен растительности, реакций на те или иные виды лесохозяйственных мероприятий и т.д.” [4, с. 39].
Индекс листовой поверхности насаждения меняется в зависимости от преобладающей породы, среднего возраста, режима управления лесами и положения на рельефе [6]; значение LAI для конкретного дерева зависит от размера листьев, их общего числа и архитектуры кроны [12]. Само по себе значение индекса листовой поверхности для конкретного насаждения не дает точной энергетической характеристики экосистемы [4]. Это возможно только при соотнесении LAI с таксационными показателями, величинами приростов, общей биомассой фитоценоза и фитомассой листвы, данными о фотосинтезе и дыхании, что все еще относительно редко встречается в научной литературе [10, 11].
LAI может быть определен следующими способами: 1) установление аллометрических зависимостей между площадью листвы и геометрическими параметрами дерева (обычно диаметр и высота); 2) оптический метод с использованием фотокамеры со сферическим объективом; 3) моделирование на основе данных космической съемки; 4) лазерное сканирование (TLS, terrestrial laser scanning), которое может быть как наземным, так и дистанционным [9, 10, 13, 14]. Последний метод является наиболее простым и одновременно точным, однако для верификации съемки системами типа LIDAR необходимы репрезентативные базы данных, полученные первыми двумя методами.
Цель настоящей работы – определение индекса листовой поверхности (LAI) весовым методом для древостоев лесных насаждений разных типов на территории Приморского края.
МАТЕРИАЛ И МЕТОДЫ
Работу выполняли на лесном участке Приморской государственной сельскохозяйственной академии – на территории, расположенной в южной части Приморского края, в 40 км к востоку от г. Уссурийска (лесной участок). В 1990-е и 2000-е годы на большей части участка были проведены выборочные рубки и рубки ухода. По таксационным характеристикам лесные насаждения здесь весьма разнообразны: от вторичных дубняков (доминирует Quercus mongolica Fisch. ex Ledeb.) до многопородных чернопихтарников (доминирует Abies holophylla Maxim.) и кедровников (доминирует Pinus koraiensis Siebold & Zucc.) [15]. Пересеченный рельеф, близость гор Пржевальского (перепады высоты 100–500 м над ур. м.) формируют здесь как пойменные, так и горные типы лесов. Климат района типично муссонный, с преобладанием солнечной морозной погоды зимой, пасмурной и влажной – летом. Среднегодовая температура воздуха колеблется от 3 до 5°С, годовая сумма осадков – от 637 до 894 мм.
Площадь поверхности листьев определяли как произведение удельной листовой поверхности SLA (specific leaf area) и фитомассы листвы в сухом состоянии для каждой породы на пробной площади; площадь листвы для пород древостоя получали суммированием площадей листовой поверхности всех деревьев (d1.3 > 6 см).
Определение удельной листовой поверхности. SLA представляет собой отношение площади поверхности листьев к их массе в абсолютно сухом состоянии и имеет размерность см2/г [4]. Удельная листовая поверхность была определена для 19 пород, формирующих древостои в условиях южного Сихотэ-Алиня: бархат амурский (Phellodendron amurense Rupr.), береза ребристая (Betula costata Trautv.), береза плосколистная (B. platyphylla Sukaczev), береза даурская (B. dahurica Pall.), граб сердцелистный (Carpinus cordata Blume), дуб монгольский (Quercus mongolica Fisch. ex Ledeb.), ильм японский (Ulmus japonica Sarg.), кедр корейский (Pinus koraiensis Siebold & Zucc.), клен ложнозибольдов (Acer pseudosieboldianum Kom.), клен маньчжурский (A. mandshuricum Maxim.), клен мелколистный (A. mono Maxim.), липа амурская (Tilia amurensis Rupr.), липа маньчжурская (T. mandshurica Rupr. & Maxim.), маакия амурская (Maackia amurensis Rupr.), ольха волосистая (Alnus hirsuta Rupr.), орех маньчжурский (Juglans mandshurica Maxim.), пихта цельнолистная (Abies holophylla Maxim.), осина (Populus tremula L.), ясень маньчжурский (Fraxinus mandshurica Rupr.).
Для каждого вида было выбрано по 10 модельных деревьев, у которых в нижней части крон с помощью секатора на штанге срезали модельные ветви в зоне 3–6 м от поверхности почвы. Растения выбирали на трансекте длиной 5 км, включающей пойменные и горные участки. Известно, что деревьям в лесном пологе присуща изменчивость индекса SLA по вертикальному профилю. Задача по выявлению вертикальной изменчивости этого показателя весьма затруднительна в связи с действующим законодательным запретом на рубку некоторых видов деревьев. Поэтому полученные результаты ценны из-за сравнения LAI между древостоями разных насаждений, и в дальнейшем могут быть уточнены с применением лазерного сканирования.
У лиственных пород сбор листвы выполняли в начале июля 2018 г., у хвойных – в конце августа. Листья с каждого дерева складывали отдельно и затем сканировали с пространственным разрешением 300 пикселей/дюйм (dpi). В программе ImageJ (версия 1.50d, разработчик National Institutes of Health) после обработки скана определяли площадь листьев (по соотношению черных и белых пикселей). После сканирования листья высушивали в сушильном шкафу при температуре 101°С. Для каждого дерева были получены отношения площади листвы к абсолютно сухой массе – удельные листовые поверхности SLA. При этом для лиственных пород использовали одностороннюю площадь листьев, а для хвойных – удвоенную площадь проекции хвоинок [4]. Так, каждая древесная порода характеризовалась средним значением SLA по 10 деревьям.
Определение фитомассы листвы. Рубка модельных деревьев и определение их надземной фитомассы по фракциям (ствол, кора, ветви и листва) были осуществлены на лесном участке ПГСХА в 2015 и 2016 гг. Подробно методы и результаты этих работ описаны в публикациях [16–19], здесь мы ограничимся описанием методических приемов, относящихся к определению фитомассы листвы.
Было выбрано 15 древесных пород, входящих в состав древостоев кедрово-чернопихтово-широколиственных лесов. Перед отбором модельных деревьев были определены таксационные показатели насаждения (состав, средние высоты и диаметры, полнота, запас и др.). Рубку модельных деревьев осуществляли в августе 2015 и 2016 гг. после полного формирования листвы и хвои, что соответствует рекомендациям [20]. Модельные деревья выбирали как средние по диаметру, высоте и размерам кроны для ступени толщины. У спиленного на уровне корневой шейки дерева определяли общую длину (высоту) и протяженность живой кроны, возраст дерева по числу годичных колец на пне. Живую крону хвойных пород делили на три равные по длине секции: верхнюю, среднюю и нижнюю. У лиственных пород выделяли одну секцию, так как плотность листвы на разных высотах значимо не отличается [20]. Обрезку кроны производили вдоль по стволу, каждую секцию взвешивали с точностью 50 г на электронных весах грузоподъемностью 50 кг. У отделенной от ствола кроны обрезали облиственные (охвоенные) побеги (древесная зелень). По разности общей массы кроны и скелета находили массу облиственных (охвоенных) побегов по секциям. Из обрезанных облиственных (охвоенных) побегов каждой секции формировали навеску массой 200–300 г. В камеральных условиях в навесках отделяли листву (хвою), оставшиеся побеги взвешивали. После сушки (до постоянной массы) проводили повторное взвешивание и определяли у побегов содержание сухого вещества и плотность. Полученные значения использовали для пересчета массы листвы и скелета кроны из свежего состояния в абсолютно сухое.
Таксационное описание пробных площадей. Исходными данными для определения LAI были перечетные ведомости древостоев на временных и постоянных пробных площадях (ПП), заложенных на лесном участке ПГСХА в разное время (36 ПП), в национальном парке “Бикин” (5 ПП) и Курортном участковом лесничестве (3 ПП). Перечетная ведомость древостоя представляет собой распределение числа деревьев по породам и диаметру на высоте 1.3 м над поверхностью почвы (d1.3) с шагом ступени толщины 4 см. На каждой ПП размерами от 50 × 50 м до 100 × 100 м определяли стандартный набор таксационных показателей: средний диаметр, среднюю высоту, абсолютную полноту, запас стволовой древесины в коре, густоту деревьев.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
В 1998 г. Л.С. Ермолова и А.И. Уткин опубликовали статью с расчетными данными SLA для основных пород лесообразователей СССР, дифференцированных преимущественно по родам растений [3]. Применение этих значений SLA для определения индексов листовой поверхности лесов юга Дальнего Востока может привести к ряду неточностей, связанных, в частности, с разнообразием видового состава древостоев. Это опасение послужило основанием для выполнения оригинальных измерений SLA на территории лесного участка ПГСХА, а данные из цитируемой работы использовали для сравнения. В табл. 1 приведены полученные значения SLA.
Таблица 1.
Удельная листовая поверхность некоторых пород лесообразователей южной части Приморского края в июне 2018 г. (М – среднее значение, m – ошибка среднего)
| Порода | SLA, см2/г | |
|---|---|---|
| М | m | |
| Пихта цельнолистная | 104.0 | 6.4 |
| Кедр корейский | 123.1 | 2.4 |
| Осина | 129.3 | 3.0 |
| Бархат амурский | 168.2 | 5.6 |
| Дуб монгольский | 180.3 | 10.3 |
| Береза плосколистная | 183.1 | 3.8 |
| Орех маньчжурский | 187.9 | 7.8 |
| Ильм японский | 191.1 | 12.8 |
| Ясень маньчжурский | 202.1 | 27.8 |
| Береза желтая | 210.1 | 9.5 |
| Береза черная | 210.3 | 13.1 |
| Липа амурская | 228.1 | 16.6 |
| Липа маньчжурская | 235.4 | 12.8 |
| Ольха волосистая | 251.8 | 12.7 |
| Маакия амурская | 272.3 | 84.0 |
| Клен маньчжурский | 285.3 | 18.5 |
| Клен мелколистный | 293.5 | 18.5 |
| Клен ложнозибольдов | 310.0 | 11.2 |
| Граб сердцелистный | 363.1 | 19.8 |
По данным Л.С. Ермоловой и А.И. Уткина [3], средняя SLA составила: кедр корейский – 152, ясень – 176, дуб – 148, липа – 276 см2/г. Отличия с нашими оценками составляют 23.5, –12.8, –17.8 и 17.4% соответственно. Таким образом, применение обобщенных коэффициентов SLA из работы [3] для лесных насаждений юга Приморья приводит к неточностям около 20%.
Все пробные площади, для которых определяли индекс листовой поверхности, были разделены на три группы. Пробную площадь относили к той или иной группе исходя из истории нарушений (рубки, пожары) и лесорастительных условий: первая группа – относительно ненарушенные хвойно-широколиственные леса, сохраняющие структуру полнодревесного насаждения; вторая группа – в насаждениях проводились выборочные рубки, в пологе сохраняются большие окна, которые занимают светолюбивые виды, также сюда отнесены постпирогенные дубняки; третья группа – пойменные леса, образованные преимущественно Fraxinus mandshurica, Ulmus japonica, Populus maximoviczii, Juglans mandshurica. Значения LAI для каждой пробной площади и их таксационные характеристики по трем описанным группам насаждений приведены в табл. 2.
Таблица 2.
Индексы листовой поверхности для насаждений пробных площадей на территории Приморского края
| № п.п. | LAI | Состав | N, шт/га | G, м2/га | M, м3/га |
|---|---|---|---|---|---|
| Относительно не нарушенные хвойно-широколиственные насаждения | |||||
| 1 | 15.41 | 6Д2Лип1К1Кл + П,Кпн,Клз | 712 | 37.0 | 350.3 |
| 2 | 16.24 | 3Я2К2Ор2Кл1Лип + Иг,Маа,Ол | 680 | 27.4 | 254.6 |
| 3 | 15.50 | 3К3Лип3Д1Кл + П,Клз | 692 | 37.4 | 355.2 |
| 4 | 15.13 | 7Д1К1Лип1Ил + Кл,Клз | 668 | 35.4 | 339.1 |
| 5 | 24.30 | 9К1Лип + П,Е,Кл,Бб,Яс,Ос | 444 | 56.4 | 680.1 |
| 6 | 16.96 | 8К1Лип1Кл + П,Е,Бб | 448 | 42.5 | 460.3 |
| 7 | 18.58 | 6К1Бж1Кл1Лп1Т + П,Е,Ия,Бх,Ор,Че | 749 | 41.2 | 434.2 |
| 8 | 15.45 | 4К2ЛП1П1КЛ1БЖ + Г,Ил,Бх,Яс,Тр,Чма | 436 | 41.3 | 418.4 |
| 9 | 20.23 | 3П2К2ЛП1Бж1Кл1Г + Я,Ил,Тр,Чр,Мел,Ор | 484 | 42.4 | 506.3 |
| 10 | 19.82 | 3П2К2Бж1Лп1Кл1Г + Ор,Д,Ил,Кол,Ма,Я,Бх,Тр | 436 | 37.1 | 466.5 |
| 11 | 21.00 | 3К3Пц2Д2Лп | 728 | 46.9 | 491.5 |
| 12 | 20.89 | 5Пц2К1Лп1Км1 + Д,Иг,Бб,Я,Бжр,Ор,Бч,Ос,Тм,Клжз | 764 | 28.6 | 517.3 |
| 13 | 16.33 | 3К3П1Лип1Бжр1Км1Т | 280 | 34.3 | 362.6 |
| 14 | 17.15 | 5Д2К1П1Лп1Кл | 870 | 38.3 | 355.5 |
| 15 | 17.82 | 2К2Пц2Д1Лп1Я1Кл1Бб | 622 | 35.2 | 319.7 |
| 16 | 15.51 | 5К1П3Д1Кл + Тр,Клз,Я,Г | 412 | 19.0 | 289.6 |
| 17 | 15.96 | 4К1П2Лип1Я1Кл1Бж + Ия,Д,Маа,Кп,Тр,Чер | 664 | 49.0 | 337.2 |
| 18 | 18.47 | 5К1П1Д1Лип1Кл1Г + Ия,Тр,Я,Бх | 496 | 45.1 | 340.0 |
| Среднее | 17.82 ± 0.60 | 588 ± 37 | 38.57 ± 0.2.01 | 404 ± 24 | |
| Нарушенные хвойно-широколиственные и вторичные леса | |||||
| 1 | 8.33 | 4Д3Кл2Лип1Ор + Ил,Маа | 552 | 17.0 | 128.0 |
| 2 | 9.44 | 5Лип3Д1Кл1Ор + Ил,Я,Бх | 1024 | 32.9 | 242.1 |
| 3 | 9.42 | 5Д5Лип + Ил,Кл,Маа,Ив | 816 | 28.1 | 211.5 |
| 4 | 10.08 | 5К1Е1Кл1Лип1Я1Ил + П,Д,Маа,Бх | 512 | 28.3 | 282.3 |
| 5 | 9.72 | 3Е2К12ЛпП1Кл1Яс1Бх + Бб,Д | 456 | 26.1 | 244.5 |
| 6 | 12.58 | 4Д 4ЛП 1Яс 1ПЦ + Кл,И,Бб,Бх,Ол,Ос | 530 | 34.1 | 334.8 |
| 7 | 10.25 | 6Ос2К1Яс1Бб + Кл,Ор,Бх,Д,Пц,Лип | 882 | 28.1 | 270.5 |
| 8 | 9.63 | 4Бб3Ос1К1Д1Кл + Яс,Ор,Лип, Ия,Маа,Бх | 774 | 25.5 | 241.7 |
| 9 | 6.01 | 3Лп2Кл2Д1П1Кол1Гр + К,Бр,Маа,КлЛ,Бп,Яс,Ил,Бж,Мпл | 500 | 23.1 | 206.7 |
| 10 | 12.73 | 6П2Д1К1ЛП + Бж,Г,Кл,Кол,Я,Бх,Тр | 508 | 29.1 | 283.2 |
| 11 | 13.18 | 6П2К1Бж1КЛ + Лп,Г,Я,ББ | 748 | 32.0 | 311.8 |
| 12 | 19.37 | 3К2П2Ил2Кл1Гр + Лп,Тр,Бж,Маак,Ор | 564 | 26.8 | 273.0 |
| 13 | 13.5 | 6Д3К1П | 792 | 31.9 | 296.3 |
| 14 | 14.44 | 5П3К1Д1Я + Лип,Кл,Клз,Бж,Маа,Г,Бб,Бх,Тр, | 452 | 29.0 | 261.2 |
| 15 | 9.71 | 4П3К2Кл1Д + Лип,Я,Бх,Г,Маа,Чм | 456 | 22.0 | 190.8 |
| 16 | 7.35 | 4К1П2Кл1Лип1Д1Бж + Г,Тр | 340 | 21.2 | 141.3 |
| 17 | 12.02 | 3К2П3Д1Кл1Г + Бх,Лип,Тр,Ос,Бб | 460 | 31.4 | 232.3 |
| 18 | 12.43 | 4К2П1Бж1Лип1Кл1Г + Ия,Тр,Я,Бх,Ос,Д,Ол,Ор | 496 | 36.8 | 264.3 |
| Среднее | 11.12 ± 0.72 | 588 ± 44 | 27.97 ± 1.17 | 245 ± 13 | |
| Пойменные леса | |||||
| 1 | 7.44 | 3Ия2Кл1Лип1Ор1К1П1Я + Д,Бх,Тр | 624 | 25.9 | 228.7 |
| 2 | 9.74 | 5ЯС1И1КЛ1К1ЛП1ББ + Д,Бх,Ор,ОЛ,СА | 486 | 23.7 | 214.8 |
| 3 | 14.00 | 4Д3Лп2П1Яс + Кл,П,Бч,Мпл,Бб,Ил,Ябл,Маа | 616 | 33.7 | 333.1 |
| 4 | 15.45 | 3Лп2Д2Я1К1П1Бб + Кл,Ил,Бч,Ма,Мпл,Бж,Кпн,Маа,Ор | 612 | 34.0 | 324.0 |
| 5 | 6.89 | 4Ор3Я2Ил1Бх + К,П,Кл,Маа,Тр,Лип,Яб | 596 | 22.0 | 189.7 |
| 6 | 6.36 | 6Т1Ор1Ия1Кл1Лип + К,Ол,Бх,Я | 542 | 41.6 | 383.8 |
| 7 | 5.86 | 6Ор1Ия1Клз1Лип1Тр + Бх,Маа | 588 | 19.4 | 169.3 |
| 8 | 4.43 | 4К1С2Ос1Д1Лип1Ил1Ив | 668 | 10.8 | 76.7 |
| Среднее | 8.77 ± 0.94 | 592 ± 13 | 26.37 ± 2.29 | 240 ± 24 | |
Значения LAI на пробных площадях изменяются от 4.43 (кедровый молодняк в пойме) до 24.30 (старовозрастный кедрач в национальном парке “Бикин”). Следует отметить, что лесные насаждения, имеющие запас древесины более 400 м3/га, могут считаться минимально нарушенными. Так, например, самый большой запас древесины и фитомассы зафиксирован в кедровнике в среднем течении р. Бикин, где длительное время отсутствовали лесные пожары, а рубка леса вообще никогда не проводилась. Поэтому здесь сформировался древостой, близкий по показателям к максимальной потенциально возможной для этой лесной формации продукции биомассы [19]. Этим объясняется и максимальное значение на данной пробной площади индекса LAI. При этом можно найти насаждения с одинаковыми запасами древесины, но отличающиеся по значениям LAI. Различия в средних значениях LAI, оцененные с помощью двухвыборочного t-критерия, являются значимыми между 1-й и 2-й и 1-й и 3-й группами насаждений, p < 0.001 (см. табл. 2). В целом можно сказать, что насаждения 2-й группы – это деградировавшие по разным причинам насаждения 1-й группы.
В Приморском крае в связи с развитием инфраструктуры, интенсивной работой лесной индустрии и регулярными лесными пожарами на протяжении более чем 150 лет происходит сокращение площадей коренных хвойно-широколиственных лесов и их трансформация во вторичные, как правило, с доминированием Quercus mongolica. Так, в Приморье зафиксировано двукратное уменьшение площадей кедровников с 4 до 2 млн га [15]. Наряду с утратой этими экосистемами своего коренного биологического разнообразия, снижения общей биомассы насаждений можно констатировать также и уменьшение LAI на 37%, что сопряжено с эквивалентным уменьшением интенсивности потоков вещества и энергии.
Относительно низкие значения индекса листовой поверхности у пойменных широколиственных видов по сравнению с хвойными, вероятно, можно объяснить особенностями распределения листвы. Листовая биомасса у ясеня, тополя, ильма преимущественно сосредоточена в верхней части дерева, и стволы в нижней части обычно хорошо очищены от сучьев, тогда как у деревьев кедра и пихты биомасса хвои находится и в средней части ствола, поэтому хвойные деревья имеют относительно большую площадь листовой поверхности.
Зависимости LAI от таксационных характеристик насаждений имеют разнообразный характер (рис. 1). LAI оказывается мало связанным с густотой леса (R2 = 0), поскольку основная часть листвы насаждения находится на крупных деревьях, а размер деревьев мало зависит от густоты их размещения в насаждении.
Рис. 1.
Связь между LAI и таксационными показателями насаждений: а – густотой; б – абсолютной полнотой; в – объемным запасом древесины.
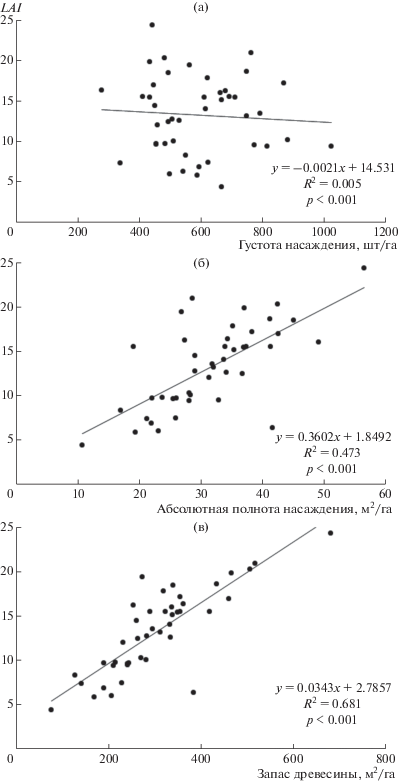
Абсолютная полнота и запас насаждения демонстрируют значимую линейную связь с индексом LAI. Запас древесины вычисляется на основе данных о диаметрах и высотах деревьев на пробной площади, поэтому коэффициент детерминации в уравнении, связывающем LAI с запасом древесины (R2 = 0.473), выше, чем в уравнении связи с абсолютной полнотой (R2 = 0.681), которая определяется только диаметрами деревьев (в обоих случаях p < 0.001).
Несмотря на достаточно высокое значение (0.68) коэффициента детерминации линейного уравнения связи между LAI и запасом насаждения, в исходном массиве данных имеются примеры значительных расхождений LAI при одном и том же запасе насаждения. Так, при запасе около 240 м3/га LAI на пяти пробных площадях меняется от 10 до 20. Можно согласиться с положением А.И. Уткина [4] о том, что LAI является самостоятельным независимым показателем, который невозможно определить по стандартным таксационным параметрам насаждения, поэтому он требует отдельного определения.
Полученные в ходе настоящего исследования значения LAI для древостоев пробных площадей на территории Приморского края в дальнейшем планируется использовать для верификации оптического и лазерного методов определения LAI.
ЗАКЛЮЧЕНИЕ
Выполненное исследование по оценке индексов листовой поверхности древостоев лесных насаждений Приморского края является одним из шагов к оценке экосистемных функций этих лесов, отличающихся максимальным для территории России биологическим разнообразием. Объемный материал с 44 пробных площадей позволил выявить закономерности изменения LAI в зависимости от типа насаждения и истории нарушений его структуры. Из всего массива данных минимальные значения LAI оказались присущи древостоям в поймах рек, максимальные свойственны высокополнотным, сложным по форме хвойно-широколиственным насаждениям с доминированием в составе древостоев хвойных пород.
Результаты нашей работы подтвердили негативное влияние нарушающих факторов – рубок и пожаров – на индекс листовой поверхности древостоев хвойно-широколиственных лесов. В среднем в результате нарушений площадь листвы на единице площади уменьшается на 37%. Если после выборочных рубок структура леса и LAI относительно быстро восстанавливаются, то регулярные лесные пожары, трансформирующие, в частности, кедровые насаждения в дубняки, уменьшают LAI необратимо. В масштабах региона вторичные леса могут рассматриваться как источник возобновляемой древесины, пусть и низкого качества. Однако многолетний процесс антропогенного преобразования коренных хвойно-широколиственных лесов во вторичные на обширных площадях ведет к изменению показателя LAI, что отражается на потоках вещества и энергии в лесных экосистемах.
Исследование выполнено в рамках темы ГЗ ЦЭПЛ РАН № АААА-А18-118052590019-7 “Методические подходы к оценке структурной организации и функционирования лесных экосистем” (полевые работы) и при поддержке проекта Российского научного фонда № 19-77-30015 “Разработка методов и технологии комплексного использования данных дистанционного зондирования Земли из космоса для развития системы национального мониторинга бюджета углерода лесов России в условиях глобальных изменений климата” (анализ данных).
Список литературы
Лукина Н.В., Исаев А.С., Крышень А.М. и др. Приоритетные направления развития лесной науки как основы устойчивого управления лесами // Лесоведение. 2015. № 4. С. 243–254.
Lähtinen K., Guan Y., Li N., Toppinen A. Biodiversity and ecosystem services in supply chain management in the global forest // Ecosystem Services. 2016. V. 21. P. 130–140. https://doi.org/10.1016/j.ecoser.2016.07.006
Ермолова Л.С., Уткин А.И. Удельная листовая поверхность основных лесообразующих пород России // Экология. 1998. № 3. С. 178–183.
Уткин А.И., Ермолова Л.С., Уткина И.А. Площадь поверхности лесных растений: сущность, параметры, использование. М.: Наука, 2008. 292 с.
Грабовский В.И., Зукерт Н.В., Корзухин М.Д. Оценка индекса листовой поверхности для территории России по данным государственного лесного реестра // Лесоведение. 2015. № 4. С. 255–259.
Glatthorn J., Pichler V., Hauck M., Leuschner Ch. Effects of forest management on stand leaf area: Comparing beech production and primeval forests in Slovakia // Forest Ecology and Management. 2017. V. 389. P. 76–85.
Nock C.A., Caspersen J.P., Thomas S.C. Large ontogenetic declines in intra-crown leaf area index in two temperate deciduous tree species // Ecology. 2008. V. 89. P. 744–753.
Liu Z., Jiang F., Zhu Yu. et al. Spatial heterogeneity of leaf area index in a temperate old-growth forest: Spatial autocorrelation dominates over biotic and abiotic factors // Science of the Total Environment. 2018. V. 634. P. 287–295.
Chen Y., Zhang W., Hu R. et al. Estimation of forest leaf area index using terrestrial laser scanning data and path length distribution model in open-canopy forests // Agricultural and Forest Meteorology. 2018. V. 263. P. 323–333.
Xin Q., Dai Y., Li X. et al. A steady-state approximation approach to simulate seasonal leaf dynamics of deciduous broadleaf forests via climate variables // Agricultural and Forest Meteorology. 2018. V. 249. P. 44–56.
Konôpka B., Pajtík J., Marušák R. et al. Specific leaf area and leaf area index in developing stands of Fagus sylvatica L. and Picea abies Karst. // Forest Ecology and Management. 2016. V. 364. P. 52–59.
Leuschner C., Voß S., Foetzki A., Clases Y. Variation in leaf area index and stand leaf mass of European beech across gradients of soil acidity and precipitation // Plant Ecology. 2006. V. 186. P. 247–258.
Calders K., Origo N., Disney M. et al. Variability and bias in active and passive ground-based measurements of effective plant, wood and leaf area index // Agricultural and Forest Meteorology. 2018. V. 252. P. 231–240.
Zhu X., Skidmore A.K., Wang T. et al. Improving leaf area index (LAI) estimation by correcting for clumping and woody effects using terrestrial laser scanning // Agricultural and Forest Meteorology. 2018. V. 263. P. 276–286.
Иванов А.В., Гамаева С.В., Панфилова Е.В. Оценка видового разнообразия растений и жужелиц на пробных площадях в кедрово-широколиственных насаждениях постпирогенного происхождения // Сибирский лесн. журн. 2018. № 3. С. 73–82.
Касаткин А.С., Жанабаева А.С., Акимов Р.Ю. и др. Надземная фитомасса и квалиметрия некоторых древесных пород южного Сихотэ-Алиня // Эко-потенциал. 2015. № 1(9). С. 41–50.
Касаткин А.С., Жанабаева А.С., Иванов А.В. и др. Надземная фитомасса деревьев в лесах южного Сихотэ-Алиня. Сообщ. 3 // Эко-потенциал. 2016. № 1(13). С. 32–36.
Касаткин А.С., Жанабаева А.С., Пауков Д.В. и др. Надземная фитомасса деревьев в лесах южного Сихотэ-Алиня. Сообщ. 2 // Эко-потенциал. 2015. № 4(12). С. 28–31.
Иванов А.В., Касаткин А.С., Мудрак В.П., Замолодчиков Д.Г. Надземная фитомасса древостоев хвойно-широколиственных лесов южного Приморья // Лесоведение. 2018. № 6. С. 454–463.
Усольцев В.А., Залесов С.В. Методы определения биологической продуктивности насаждений. Екатеринбург: УГЛТУ, 2005. 147 с.
Дополнительные материалы отсутствуют.