

Экология, 2020, № 5, стр. 365-376
Двадцатилетняя динамика экстразональных степей Южного Урала на фоне климатических изменений
Н. В. Золотарева *
Институт экологии растений и животных УрО РАН
620144 Екатеринбург, ул. 8 Марта, 202, Россия
* E-mail: nvp@ipae.uran.ru
Поступила в редакцию 10.04.2020
После доработки 15.04.2020
Принята к публикации 23.04.2020
Аннотация
С помощью прямых многолетних наблюдений прослежены изменения видового состава и структуры экстразональных степей в бореальной зоне Южного Урала, являющихся местообитанием большого числа редких растений. Показано, что за период 1999–2019 гг. произошла мезофитизация сообществ петрофитных степей: в 6.8 раза увеличилось проективное покрытие лугово-степных и степных видов, в то время как проективное покрытие петрофитно-степных видов, составляющих основу исследованных сообществ, возросло только в 2.2 раза. Часть петрофитных степей была поглощена окружающими луговыми степями. Выявлено, что наибольшим изменениям подвержены сообщества, имеющие площадь менее 40 м2. Наблюдаемые изменения, вероятно, связаны с возрастанием количества осадков в начале вегетационного сезона.
Изучение реакции растительного мира на происходящие климатические изменения важно для сохранения уязвимых сообществ, занимающих небольшие площади, но определяющих существенную долю регионального флористического разнообразия, таких как растительность серпентинитов, известняков, высокогорий [1–6]. На Урале к таким сообществам относятся петрофитные степи. Это характерный элемент растительности региона, местообитания большого числа редких и охраняемых растений, эндемиков и реликтов [7–9]. Из 64 эндемичных видов, встречающихся в Челябинской области – районе наших исследований, половина произрастает преимущественно в петрофитных степях [8]. Из 188 видов покрытосеменных растений, охраняемых на территории Челябинской области, третья часть также приурочена к петрофитным степям [9].
В бореальной зоне Среднего и Южного Урала петрофитные степи сохранились в экстремальных для древесной растительности почвенно-климатических условиях и являются экстразональной растительностью [10, 11]. Экстразональные степи – удобный объект для изучения динамики, так как относятся к сообществам, в местообитаниях которых рельефно проявляется влияние абиотических факторов.
Накоплено достаточное количество подтверждений возрастания среднегодовой температуры и количества осадков на Южном Урале с конца 1960-х годов [12, 13]. Хотя ряд исследований [3, 4, 14] свидетельствует о том, что ксерофитные типы растительности на бедных почвах устойчивы к повышению увлажнения, в других работах показана их чувствительность к возрастанию количества осадков [1, 15, 16]. Существуют опасения, что дальнейший рост увлажнения приведет к превращению экстразональных степей в луга и их облесению [17, 18]. В последние десятилетия в пределах бореальной зоны на Урале и в Забайкалье отмечено сокращение площадей степной растительности вследствие их замещения лесной растительностью [18–20].
Цель данного исследования – используя исторические фотоматериалы, оценить изменения в распределении степной и лесной растительности за последние 90 лет; с помощью прямых многолетних наблюдений выявить изменения видового состава и структуры экстразональных петрофитных степей Южного Урала, произошедшие за период 1999–2019 гг. на фоне климатических изменений, и определить их вероятные причины.
РАЙОН И МЕТОДИКА ИССЛЕДОВАНИЙ
Район исследования находится в северной части Южного Урала в пределах Ильменского государственного заповедника, на восточном макросклоне Уральских гор, в подзоне предлесостепных сосново-березовых лесов на границе с лесостепью [8]. Территория относится к Ильменскому округу Ильменско-Сакмарской провинции Южного Урала, характеризуется умеренно-прохладным климатом с достаточным увлажнением [21].
Исследования проводили на Демидовских сопках (высота 320–336 м над ур. м.) – восточных предгорьях Ильменского хребта. Они сложены серпентинитами, имеют куполообразную форму, довольно крутые склоны. Поверхность сопок занята плотно слежавшимся щебнем с примесью мелкозема, на глубине 10–15 см переходящим в коренную горную породу. С этими сопками связаны наиболее выраженные и богатые по флористическому составу степные сообщества Ильменского заповедника. Северные склоны сопок покрыты лиственнично-сосновым редколесьем с остепненным травяным покровом, а южные и юго-западные – комплексом степных фитоценозов [10].
В 1999 г. на 7 сопках от вершины к подножию степных склонов были заложены профили; в 1999, 2014 и 2019 гг. для каждого фитоценоза, отмеченного на профиле, выполнены геоботанические описания, измерена глубина почвы. Глубину почвенного слоя определяли путем погружения в почву металлической спицы до первого препятствия [22], в каждом сообществе делали 10 измерений. В зависимости от протяженности в 1999 г. профили разделены на две группы: “короткие” – длиной от 13 до 27.5 м и “длинные” – от 53 до 69 м (табл. 1).
Таблица 1.
Экотопические характеристики постоянных пробных площадей
| № пробной площади и сопки | Крутизна склона, град/ экспозиция, град | Длина профиля через степные сообщества, м | S пробной площади, м2 | Глубина почвенного слоя, см | |
|---|---|---|---|---|---|
| 1999 г. | 2019 г. | ||||
| 1 | 14/180 | 27.5 | 24 | 22 | 6.0 |
| 2 | 17/210 | 22 | 22 | 30 | 6.4 |
| 3 | 22/210 | 15 | 13 | 30 | 5.5 |
| 4 | 15/170 | 13 | 10 | 16 | 5.7 |
| 5 | 15/195 | 69 | 37 | 36 | 5.0 |
| 6 | 22/210 | 53 | 49 | 53 | 7.2 |
| 7 | 18/210 | 63 | 57 | 69 | 5.3 |
В верхней части каждого профиля находятся наиболее крутые, сухие и прогреваемые участки, которые заняты петрофитными степями с доминированием петрофитно-степных видов: Artemisia commutata, A. frigida, Centaurea sibirica, Echinops crispus, Dianthus acicularis, Festuca valesiaca. Эти сообщества были выбраны в качестве ключевых для осуществления детального мониторинга. В системе эколого-флористической классификации они относятся к ассоциации Stipo pennatae–Centauretum sibiricae Yamalov et al. ex Zolotareva, Korolyuk, Yamalov 2019 [11, 23]. Изученные степные сообщества являются коренными, их местообитания характеризуются условиями, экстремальными для существования древесной растительности.
В 1999 г. в 7 ключевых сообществах петрофитных степей были заложены постоянные пробные площади, ограничивающие наиболее типичную часть фитоценоза и отстоящие от его естественных границ с целью избежать экотонного эффекта. Размеры постоянных пробных площадей зависели от площади ключевого фитоценоза, в связи с чем различались на разных сопках (см. табл. 1). На каждой постоянной пробной площади в 1999, 2009, 2014 и 2019 гг. выполнены геоботанические описания; на 20 (в 1999 г.) и 24 (в остальные годы наблюдений) пробных площадках размером 50 × 50 см, заложенных рандомизированно, выявлен флористический состав, проективное покрытие (ПП) каждого вида в %. За весь период наблюдений на постоянных пробных площадях отмечено 60 видов высших растений, в том числе 4 одно-двулетних видов, злаков – 11 видов, осок – 2, разнотравья – 36, полукустарничков – 4, кустарников – 2, деревьев – 1.
Сообщества, описанные на профилях, можно отнести к двум типам: 1) петрофитные степи; 2) луговые степи, остепненные луга. Некоторые сообщества характеризуются как переходные между этими двумя типами. Для того чтобы определить их местоположение, а также выявить изменения, произошедшие за 20 лет, в программе Past 3.06 [24] был осуществлен анализ сходства сообществ, описанных в 1999 г. (31 сообщество), а затем в 2019 г. (29 сообществ). Луговые степи и остепненные луга отнесены к порядку Brachypodietalia pinnati Korneck 1974, петрофитные варианты луговых степей – к порядку Helictotricho-Stipetalia Toman 1969 класса Festuco-Brometea Br.-Bl. et Tx. ex Soó 1947 [11].
В каждом сообществе вычисляли ПП нескольких ценотических групп: 1) петрофитно-степных видов, составляющих основу петрофитных степей; 2) лугово-степных и степных видов, эколого-фитоценотический оптимум которых находится в луговых степях и остепненных лугах, окружающих петрофитные участки склонов; 3) луговых, лугово-лесных, лесных видов, характерных для луговых и лесных сообществ. Мы предполагали, что соотношение ПП этих ценотических групп может стать фитоценотическим индикатором происходящих изменений.
Принадлежность вида к ценотической группе определена согласно П.В. Куликову [8] и Л.В. Рязановой [25]. В случае, когда мнение автора статьи относительно ценотической характеристики вида не совпадало с информацией из вышеуказанных источников, проводили специальное исследование: на основе 29 описаний степных сообществ, выполненных на профилях в 1999 г., с помощью коэффициента корреляции Спирмена (rS) выявляли группы сопряженных видов и связь ПП вида с глубиной почвенного слоя. Специально рассмотрены 3 вида: Artemisia commutata, Festuca valesiaca, Seseli ledebourii. По мнению П.В. Куликова [8] и Л.В. Рязановой [25], Artemisia commutata относится к лугово-степным видам, в то же время известно, что на Урале этот вид ведет себя как петрофитно-степной, является характерным элементом петрофитных степей [23]. Это подтверждается положительными rS между ПП Artemisia commutata и петрофитно-степных видов (rS = 0.57–0.84, p = = 0.002), ценотическая приуроченность которых не вызывает сомнений: Alyssum obovatum, Artemisia frigida, Dianthus acicularis; отрицательным rS между ПП Artemisia commutata и глубиной почвы (rS = –0.72, p < 0.001). Festuca valesiaca и Seseli ledebourii относят к степным видам [8, 25], указывая при этом на предпочтение ими щебнистого субстрата. На исследуемой территории эти виды характерны именно для петрофитных степей [26]. Это подтверждается положительными rS между их ПП и ПП петрофитно-степных видов (для F. valesiaca rS = 0.54–0.74, p = 0.001–0.004; для S. ledebourii rS = 0.43–0.72, p = 0.001–0.026), а также отрицательными rS между их ПП и глубиной почвы (для F. valesiaca rS = –0.64, p < 0.001; для S. ledebourii rS= = –0.60, p < 0.001).
Таким образом, к группе петрофитно-степных видов отнесены: Allium rubens, A. strictum, Alyssum obovatum, Artemisia commutata, A. frigida, Carex pediformis, Centaurea sibirica, Clausia aprica, Dianthus acicularis, Echinops crispus, Elytrigia lolioides, E. reflexiaristata, Eremogone micradenia, Euphorbia gmelinii, E. segueriana, Festuca valesiaca, Helictotrichon desertorum, Lychnis sibirica, Onosma simplicissimum, Pilosella echioides, Scorzonera glabra, Seseli ledebourii, Thalictrum foetidum; к группе лугово-степных видов – Adonis vernalis, Aretmisia macrantha, A. latifolia, A. sericea, Aster amellus, Campanula sibirica, C. wolgensis, Carex praecox, Cerastium arvense, Cotoneaster melanocarpus, Dianthus versicolor, Dracoceptalum ruyschiana, Eupharisia pectinata, Fillipendula vulgaris, Fragaria viridis, Galatella angustissima, Galatella biflora, Galium tinctorium, G. verum, Genista tinctoria, Helictotrichon schellianum, Hypericum elegans, Medicago falcata, Myosotis immitata, Phleum phleoides, Poa angustifolia, Pulsatilla uralensis, Rumex thyrsiflorus, Scorzonera purpurea, Seseli libanotis, Silene amoena, Stipa pennata, Thalictrum minus, Thesium refractum, Veronica spicata, Vincetoxicum hirundinaria; к группе степных – Koeleria cristata, Potentilla humifusa, Spiraea crenata, Stipa capillata, S. zalesskii. Латинские названия таксонов приведены по П.В. Куликову [8].
Изменение распределения степной и лесной растительности в районе исследования оценивали также с использованием изображений на фотопластинах, выполненных Л.Н. Тюлиной в 1926–1928 гг., хранящихся в фотоархиве Ботанического музея БИН РАН.
При наличии в исследуемых степных сообществах сосны определяли ее возраст: у каждого дерева диаметром ствола менее 3–4 см – путем подсчета числа мутовок ветвей, у деревьев диаметром ствола более 3–4 см брали буровой образец древесины максимально близко к корневой шейке и подсчитывали число годичных колец [28].
Изменения видового состава фитоценозов оценивали коэффициентом Серенсена (CS). Статистический анализ данных осуществляли двухфакторным дисперсионным анализом с оценкой попарных различий с помощью критерия Тьюки в программе Statistica v.6. Для сравнения параметров сообществ, описанных на профилях в разные годы, использовали t-критерий Стьюдента для повторных переменных. Для выявления связи между признаками применяли коэффициент корреляции Пирсона (r). Классы встречаемости видов определены по шкале: I – 1–20%; II – 21–40%; III – 41–60%; IV – 61–80%; V – 81–100%. Гомогенность ключевых сообществ оценивали как отношение среднего числа видов на площадке 0.25 м2 к общему числу видов в сообществе, выраженное в процентах; чем выше его значение, тем однороднее горизонтальная структура сообщества [27].
РЕЗУЛЬТАТЫ
Изменение климатических условий
Проанализированы суточное количество осадков и средняя суточная температура воздуха за период 1936–2018 гг. на метеостанции Златоуст [29], располагающейся в 35 км к северо-западу от Демидовских сопок также в лесной зоне. В наибольшей степени изменения температуры воздуха и количества осадков проявились с конца 1960-х гг.: период 1970–2018 гг. по сравнению с периодом 1936–1969 гг. (табл. 2) характеризуется значительным ростом температуры в декабре–марте (на 1.3–2°C выше) и меньшим увеличением в апреле–ноябре (на 0.1–0.9°C выше). Средняя годовая температура воздуха за весь период наблюдений была равна 1.5°C; в 1970–2018 гг. по сравнению с 1936–1969 гг. она существенно выросла (1.8 и 1.0°C соответственно).
Таблица 2.
Средние значения температуры и количества осадков (M ± m) в 1936–1969 и 1970–2018 гг.
| Месяц | Средняя температура воздуха, °C | Среднее месячное кол-во осадков, мм | ||
|---|---|---|---|---|
| 1936–1969 гг. | 1970–2018 гг. | 1936–1969 гг. | 1970–2018 гг. | |
| Январь | –14.8 ± 0.6 | –13.5 ± 0.5 | 24.6 ± 2.3 | 33.3 ± 2.5 |
| Февраль | –13.6 ± 0.6 | –12.1 ± 0.5 | 19.3 ± 2.3 | 26.3 ± 1.9 |
| Март | –7.7 ± 0.5 | –5.7 ± 0.4 | 28.3 ± 2.8 | 30.5 ± 2.8 |
| Апрель | 2.5 ± 0.4 | 3.1 ± 0.4 | 33.1 ± 3.3 | 40.8 ± 3.1 |
| Май | 9.8 ± 0.4 | 10.3 ± 0.3 | 61.9 ± 5.6 | 59.4 ± 4.4 |
| Июнь | 14.8 ± 0.3 | 15.0 ± 0.3 | 76.5 ± 7.4 | 80.3 ± 5.3 |
| Июль | 16.4 ± 0.2 | 16.6 ± 0.3 | 110.1 ± 8.5 | 108.9 ± 7.8 |
| Август | 14.2 ± 0.2 | 14.3 ± 0.2 | 75.6 ± 7.4 | 81.1 ± 5.5 |
| Сентябрь | 8.6 ± 0.3 | 8.8 ± 0.3 | 68.0 ± 6.8 | 65.5 ± 4.6 |
| Октябрь | 0.8 ± 0.3 | 1.5 ± 0.3 | 56.3 ± 4.1 | 67.2 ± 4.7 |
| Ноябрь | –7.2 ± 0.5 | –6.3 ± 0.4 | 42.5 ± 3.5 | 49.8 ± 2.7 |
| Декабрь | –12.7 ± 0.6 | –11.3 ± 0.5 | 33.7 ± 2.8 | 38.3 ± 2.5 |
Средняя годовая сумма осадков за весь период наблюдений составляла 660 мм, на холодный период года (с октября по март) приходится 34.6% (228.5 мм) всех годовых осадков, а основное количество выпадает с мая по август (см. табл. 2). В 1970–2018 гг. по сравнению с 1936–1969 гг. выросло как среднее годовое количество осадков (681.3 и 630 мм соответственно), так и количество осадков в теплый (436.0 и 425.3 мм соответственно) и, особенно, холодный (245.3 и 204.7 мм соответственно) периоды.
Для степной растительности наиболее важным является весенне-раннелетний период (апрель–июнь), определяющий прирост биомассы степного сообщества. Для выявления изменений климатических условий в этот период сравнили климатические параметры за 18 лет, предшествовавших началу наблюдений, и за 18 лет, прошедших после начала наблюдений (табл. 3). По сравнению с предшествовавшим периодом в 2000–2018 гг. выросли эффективные температуры (сумма среднесуточных температур выше 5°C) апреля и мая (на 1.2 и 35°C соответственно), а также количество осадков. Наибольший рост количества осадков произошел в июне (на 21.2 мм), при этом сумма эффективных температур уменьшилась на 22.8°C. В июле на рассматриваемой территории выпадает наибольшее количество осадков (см. табл. 2), и, несмотря на их сокращение в 2000–2018 гг. на 16.4 мм (см. табл. 3), июль продолжает оставаться самым влагообеспеченным месяцем, при этом сумма эффективных температур увеличивается незначительно (на 9.8°C), что не создает экстремальных условий для экстразональной степной растительности.
Таблица 3.
Суммы эффективных температур и среднее месячное количество осадков (M ± m) в апреле–августе 1981–1999 и 2000–2018 гг.
| Месяц | Сумма эффективных температур, °C | Среднее месячное кол-во осадков, мм | ||
|---|---|---|---|---|
| 1981–1999 гг. | 2000–2018 гг. | 1981–1999 гг. | 2000–2018 гг. | |
| Апрель | 90.4 ± 18.0 | 91.6 ± 13.9 | 39.7 ± 5.1 | 41.0 ± 5.6 |
| Май | 283.0 ± 14.7 | 318.1 ± 15.5 | 62.1 ± 7.0 | 65.1 ± 6.7 |
| Июнь | 468.3 ± 14.1 | 445.5 ± 13.6 | 68.2 ± 6.5 | 89.4 ± 11.0 |
| Июль | 516.5 ± 15.0 | 526.2 ± 13.3 | 115.4 ± 15.0 | 99.0 ± 13.1 |
| Август | 430.4 ± 11.0 | 473.1 ± 16.2 | 86.1 ± 8.7 | 78.1 ± 10.0 |
Зарастание степных участков древесной растительностью
На территории Ильменского заповедника, так же как и на Среднем Урале, мы отмечаем наступление лесной растительности на степные участки. Метод повторных ландшафтных фотографий [30] позволяет продемонстрировать изменения в распределении лесной и степной растительности на склонах Демидовских сопок за последние 90 лет – на фотографиях 1926–1928 гг. видны обширные степные склоны, деревья представлены единичными экземплярами (рис. 1). Повторные фотографии двух сопок свидетельствуют о том, что на небольших по площади степных склонах происходят наиболее заметные изменения. На повторной фотографии сопки № 3, относимой к “коротким” профилям, незаросшим остался только короткий степной склон, в то время как протяженный степной склон сопки № 6 (“длинный профиль”) почти не изменился, изменения произошли только на вершине сопки, где увеличилась сомкнутость древесного яруса и развивается густой подрост сосны.
Рис. 1.
Изменение растительности Демидовских сопок за период 1926–2019 гг. Фото 1926–1928 гг. – Л.Н. Тюлиной, фото сопки № 6 2009 г. – М.П. Золотарева, фото сопки № 3 2019 г. – Н.В. Золотаревой.

Видимые изменения в соотношении степной и лесной растительности произошли также с начала наших наблюдений: во время первого обследования (1997–1999 гг.) подрост сосны в ложбинах и микропонижениях на склонах имел высоту не более 100 см и изреживался в результате усыхания. При повторном обследовании в 2009 г. небольшие ложбины по склону заросли преимущественно сосной, которая в 2019 г. достигала 6–7 м (рис. 2). За 20-летний период наблюдений массовое появление сосны произошло в фитоценозах остепненных лугов у подножия всех степных склонов, где к 2019 г. сформировался молодой сосновый лес возрастом 22–23 года.
Рис. 2.
Массовое появление сосны на степном склоне Демидовских сопок, вид со стороны оз. Демидовского: а – 2009 г., фото Н.В. Золотаревой; б – 2019 г., фото Е.Н. Подгаевской.

Сосна поселяется также на вершинах сопок в сообществах луговых степей. Так, в 1999 г. степной склон сопки № 5 характеризовался значительной протяженностью, на пологой, хорошо выраженной вершине существовала разнотравно-злаковая луговая степь, у подножия склона – наземновейниково-клубничный луг с редким подростом сосны, достигавшим 60 см. В 2019 г. вершина склона была занята подростом сосны, имеющим высоту 2.5 м, возраст 16–17 лет и ПП 40–60%, у подножия склона остепненный луг заместился молодым сосновым лесом, в связи с чем длина профиля сократилась более чем на 30 м (cм. табл. 1). Уменьшение длины профилей через степную растительность отмечено на всех исследованных сопках, в основном за счет облесения остепненных лугов у подножия. Поселение сосны произошло не только на вершинах и у подножия сопок, но и на степных склонах, в том числе и в сообществах петрофитных степей: в 2009 г. на сопках № 1 и 7 отмечены 10–11 экз. сосны минимальным возрастом 8–10 лет, в 2019 г. большая их часть сохранилась.
Динамика степной растительности на профилях
Чтобы проследить за изменениями растительности профилей, которые представлены не только облесением, описания сообществ, относимых к травяной растительности, мы проанализировали с помощью кластерного анализа и определили их принадлежность к одному из двух типов (рис. 3): петрофитным степям (кластер а) или луговым степям и остепненным лугам (кластер б).
Рис. 3.
Дендрограмма сходства видового состава степных фитоценозов. Использованы Евклидово расстояние и метод Варда: а – петрофитные степи, б – луговые степи и остепненные луга; 1 – луговые степи и остепненные луга, 2 – петрофитные степи; 3 – петрофитные степи, ставшие за время наблюдения луговыми.
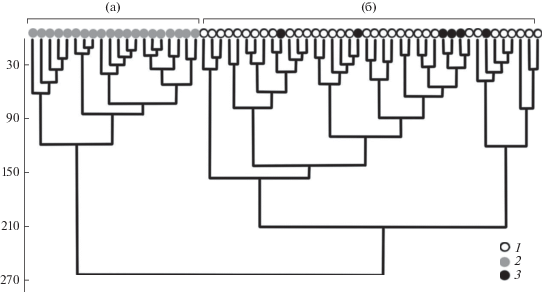
За прошедшие 20 лет к петрофитным степям перестали относиться 7 сообществ, перейдя в кластер б, 6 из них – это петрофитные степи небольшой площади, располагающиеся на выпуклых коротких участках склона в нижней части длинных профилей, в окружении луговых степей. Еще в 2014 г. контуры этих сообществ различались, хотя доминировавшие здесь в 1999 г. петрофитно-степные виды сменились лугово-степными. В 2019 г. ранее петрофитные сообщества уже не выделялись на фоне луговых степей и остепненных лугов, а доминантами в них стали лугово-степные и степные виды (Fragaria viridis, Medicago falcata, Stipa pennata, S. zalesskii и др.). Ключевые петрофитно-степные сообщества, наиболее ксерофитные и крупные по площади, не претерпели существенных изменений, за исключением сообщества сопки № 3, перешедшего в 2019 г. в кластер б. Также в 2019 г. перестала относиться к петрофиным степям часть ключевого сообщества сопки № 1, приуроченная к более пологому склону. К 2019 г. фитоценотическая неоднородность профилей, обусловленная особенностями микрорельефа, постепенно сгладилась. Объединение нескольких ранее дифференцирующихся сообществ в нижней части длинных профилей в одно увеличило однородность их растительности.
Чтобы оценить направление и степень произошедших изменений, была проанализирована ценотическая структура двух типов сообществ: петрофитных степей (14 пар описаний), а также луговых степей и остепненных лугов (16 пар описаний) за 1999 и 2019 гг. (табл. 4). Соотношение видов различных ценотических групп в сообществах луговых степей и остепненных лугов практически не изменилось, осталась прежней и группа доминирующих видов, значимо увеличилось (в 2 раза) лишь ПП видов, характерных для луговых и лесных сообществ. В основном это наблюдается в нижних частях склонов, где степные сообщества граничат с молодым сосновым лесом и большую часть дня затенены. Среди лесных видов, поселившихся в опушечных степных сообществах, оказался представитель семейства Orchidaceae – Neottianthe cuculata, характерный для сосновых лесов. Наибольшие изменения произошли в сообществах петрофитных степей: за 20 лет в 1.4 раза уменьшилась доля ПП петрофитно-степных видов, составляющих основу данных сообществ, и в 1.6 раза увеличилась доля ПП лугово-степных и степных видов (см. табл. 4).
Таблица 4.
Проективное покрытие (M ± m) основных ценотических групп видов* в степных фитоценозах с 1999 г. по 2019 г.
| Ценотические группы видов | Петрофитные степи (n = 14) | Луговые степи, остепненные луга (n = 16) | ||||
|---|---|---|---|---|---|---|
| 1999 г. | 2019 г. | p** | 1999 г. | 2019 г. | p | |
| Петрофитно-степные | 69.1 ± 3.9 | 50.0 ± 7.2 | 0.001 | 28.6 ± 3.3 | 23.4 ± 2.9 | 0.213 |
| Лугово-степные и степные | 27.0 ± 3.4 | 42.8 ± 5.8 | 0.002 | 60.6 ± 2.8 | 55.1 ± 3.9 | 0.256 |
| Луговые, лугово-лесные, лесные | 3.8 ± 0.9 | 7.2 ± 2.7 | 0.250 | 10.8 ± 1.8 | 21.5 ± 3.8 | 0.014 |
Динамика ключевых сообществ петрофитных степей
Наибольший интерес представляет динамика ключевых сообществ петрофитных степей, занимающих самые крутые, сухие и прогреваемые части склонов при дефиците увлажнения.
Смена доминантов. Во всех сообществах во все годы учетов регистрировали смену одного или двух доминантов. В 1999 и 2009 гг. доминантами были характерные виды петрофитных степей; с 2014 г. позиции доминантов начали занимать лугово-степные и степные виды (табл. 5). В 2019 г. только в двух ключевых фитоценозах, имеющих наибольшую площадь и относящихся к “длинным” профилям, доминанты представлены исключительно петрофитно-степными видами, в то время как в пяти других ключевых фитоценозах доминируют лугово-степные и степные виды.
Таблица 5.
Изменение состава доминантов в фитоценозах петрофитных степей за 20 лет
| № сопки | 1999 г. | 2009 г. | 2014 г. | 2019 г. |
|---|---|---|---|---|
| 1 | Artemisia frigida, 20* Dianthus acicularis, 15 Festuca valesiaca, 15 |
Artemisia frigida, 20 Festuca valesiaca, 20 |
Alyssum obovatum, 20 Artemisia frigida, 20 |
Festuca valesiaca, 25 Artemisia frigida, 7 Dianthus acicularis, 7 Stipa pennata, 7 |
| 2 | Artemisia frigida, 20 Dianthus acicularis, 15 |
Dianthus acicularis, 25 Festuca valesiaca, 30 |
Artemisia frigida, 30 Festuca valesiaca, 15 Dianthus acicularis, 15 |
Artemisia frigida, 15 Festuca valesiaca, 25 Stipa pennata, 15 |
| 3 | Artemisia frigida, 20 Echinops crispus, 20 |
Artemisia frigida, 25 Dianthus acicularis, 15 |
Stipa pennata, 20 Artemisia frigida, 30 |
Stipa pennata, 20 Artemisia frigida, 30 Helictotrichon desertorum, 20 |
| 4 | Artemisia frigida, 20 Echinops crispus, 40 |
Dianthus acicularis, 25 Echinops crispus, 40 |
Echinops crispus, 30 Medicago falcata, 15 Phleum phleoides, 15 |
Echinops crispus, 30 Medicago falcata, 20 Phleum phleoides, 20 |
| 5 | Alyssum obovatum, 15 Artemisia frigida, 15 Centaurea sibirica, 20 |
Centaurea sibirica, 20 Dianthus acicularis, 15 Festuca valesiaca, 15 |
Artemisia commutata, 10 Artemisia frigida, 15 Stipa pennata, 10 |
Artemisia frigida, 15 Festuca valesiaca, 10 Centaurea sibirica, 10 Stipa pennata, 10 |
| 6 | Artemisia commutata, 15 Artemisia frigida, 10 Echinops crispus, 15 |
Artemisia frigida, 15 Dianthus acicularis, 15 Festuca valesiaca, 15 |
Stipa capillata, 15 Artemisia commutata, 15 Artemisia frigida, 15 |
Dianthus acicularis, 15 Artemisia commutata, 15 Festuca valesiaca, 7 |
| 7 | Artemisia frigida, 15 Dianthus acicularis, 15 Festuca valesiaca, 15 |
Artemisia frigida, 20 Centaurea sibirica, 15 |
Artemisia frigida, 25 Festuca valesiaca, 20 |
Festuca valesiaca, 25 Artemisia commutata, 10 Artemisia frigida, 10 |
Изменение состава и встречаемости видов. Видовой состав исследованных фитоценозов не претерпел серьезных изменений: значения качественного коэффициента Серенсена в течение всего периода наблюдений находились в пределах 0.78–0.95. Число видов в сообществе и видовая насыщенность не демонстрировали однонаправленных изменений (рис. 4а). При этом гомогенность сообщества к 2009 г. возросла в 1.3 раза (p < 0.001) и слабо увеличивалась до конца наблюдений (рис. 4б). О возрастании гомогенности пространственной структуры ключевых фитоценозов свидетельствует также изменение распределения видов по классам встречаемости. От начала к концу наблюдений в ключевых сообществах в 1.3 раза уменьшилась доля видов с низкой встречаемостью (I и II классы), в то время как доля видов со средней и высокой встречаемостью (III–V классы) возросла в 1.5 раза (рис. 4в).
Рис. 4.
Показатели ключевых сообществ петрофитных степей. Приведены средние значения и 95%-ный доверительный интервал, одинаковыми буквенными индексами показаны гомогенные по критерию Тьюки значения: а – среднее число видов в сообществе (1) и среднее число видов на 0.25 м2 (2); b – виды I–II классов встречаемости (1) и III–V классов встречаемости (2).
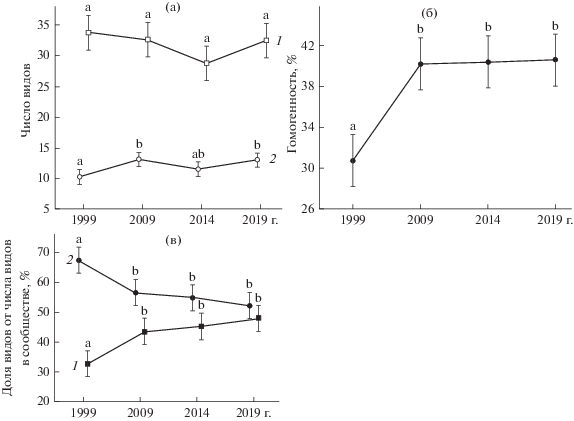
Выявлены 17 видов, встречаемость которых за время наблюдений превысила 40% (т.е. эти виды перешли из I–II в III–V классы встречаемости); все они, за исключением Elytrigia lolioides, размножаются исключительно семенным путем; 9 из них относятся к петрофитно-степным, а 8 – к лугово-степным и степным. Виды, перешедшие в III–V классы встречаемости, отмечены в каждом ключевом сообществе. В ключевых сообществах малой площади (менее 40 м2) в среднем выявлено 6.8 вида, в более крупных сообществах (более 50 м2) – в среднем 3.5 вида. Отмечена отрицательная корреляция числа видов, перешедших в III–V классы встречаемости, с площадью ключевого сообщества (r = – 0.76, p = 0.048). Это свидетельствует о стабильности более крупных степных сообществ.
Изменение ценотической структуры сообществ. Наблюдаемые изменения пространственной структуры петрофитных степей отражаются и в ценотической структуре сообщества. На первом этапе наблюдений на всех пробных площадях произошел рост ПП как петрофитно-степных, так и лугово-степных и степных видов (рис. 5). В 2014 г. отмечено снижение ПП петрофитно-степных видов и дальнейший рост ПП лугово-степных и степных. В 2019 г. ПП петрофитно-степных видов, а также лугово-степных и степных возросло на большинстве сопок. В среднем от начала к концу наблюдений ПП петрофитно-степных видов выросло в 2.2 раза, а лугово-степных и степных – в 6.8 раза.
Рис. 5.
Суммарное проективное покрытие и проективное покрытие основных ценотических групп видов в ключевых сообществах петрофитных степей с 1999 г. по 2019 г. Приведены средние значения и 95%-ный доверительный интервал; одинаковыми буквенными индексами для каждого параметра показаны гомогенные по критерию Тьюки значения: 1 – лугово-степные и степные виды; 2 – петрофитно-степные виды; 3 – суммарное проективное покрытие.
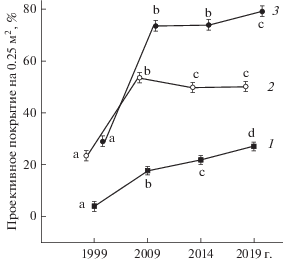
Результаты дисперсионного анализа на основе данных о ПП для 7 ключевых сообществ на 644 площадках размером 0.25 м2 свидетельствуют о том, что на суммарное ПП, возросшее за 20-летний период наблюдений (см. рис. 5), основное влияние оказывал фактор “год” (табл. 6), как и на ПП петрофитно-степных, лугово-степных и степных видов. Фактор “сопка”, т.е. локальные особенности местообитаний, и взаимодействие факторов влияли в меньшей степени. Выявлена высокая отрицательная корреляция степени увеличения ПП лугово-степных и степных видов на 0.25 м2 от начала к концу наблюдений (отношение ПП в 2019/1999 гг.) с площадью ключевого сообщества (r = –0.85, p = 0.015). Для петрофитно-степных видов такая зависимость не установлена.
Таблица 6.
Результаты двухфакторного дисперсионного анализа для проективного покрытия основных ценотических групп видов в ключевых сообществах петрофитных степей (приведено значение F-критерия Фишера)
| Проективное покрытие на площадке 0.25 м2 |
Источник варьирования | |||||
|---|---|---|---|---|---|---|
| год (dF = 3) | сопка (dF = 6) | взаимодействие факторов (dF = 18) | ||||
| F | p | F | p | F | p | |
| Петрофитно-степные виды | 202.2 | <0.001 | 20.1 | <0.001 | 8.5 | <0.001 |
| Лугово-степные и степные виды | 161.0 | <0.001 | 75.0 | <0.001 | 14.1 | <0.001 |
| Суммарное проективное покрытие | 517.8 | <0.001 | 84.5 | <0.001 | 5.8 | <0.001 |
ОБСУЖДЕНИЕ
Степные сообщества в пределах лесной зоны Урала имеют реликтовую природу [7]. В основном они представлены петрофитными степями; занимаемые ими местообитания отличаются экстремальными условиями, что способствует сохранению петрофитных степей в окружении лесной растительности: каменистые склоны южных экспозиций с неглубоким почвенным профилем характеризуются жестким радиационным режимом, резкими сезонными и суточными перепадами температуры и влажности почвы [10, 31]. Как и петрофитные степи Уральской горной страны в целом, петрофитные степи Демидовских сопок отличаются оригинальным флористическим комплексом, в котором присутствуют эндемичные (Dianthus acicularis, Elytrigia reflexiaristata, Euphorbia gmelinii) и реликтовые (Alyssum obovatum, Centaurea sibirica, Clausia aprica, Euphorbia seguieriana, Koeleria cristata и др.) виды, что подтверждает их коренную природу.
Формирование флористического комплекса каменистых степей началось в плейстоцене. Несмотря на существенные преобразования среды и растительного покрова, происходившие в последующие геологические периоды, каменистые степи продолжили свое существование, так как даже в теплые влажные эпохи, сопровождающиеся интенсивным расселением лесов, в местообитаниях, которые они занимали, была исключена конкуренция со стороны древесных растений [10]. Граница между степными и лесными сообществами неоднократно смещалась, о чем свидетельствует наличие почв черноземного ряда под лиственнично-сосновыми редколесьями, окружающими участки каменистых степей на Демидовских сопках [32].
Результаты 20-летних наблюдений свидетельствуют о наступлении лесной растительности на степные участки, что проявляется в поселении сосны на вершинах и у подножия степных склонов. Зарастание экстразональных степей сосной отмечено также на Среднем Урале, где начало этого процесса совпало с увеличением количества осадков с 1945 г. Менее чем за 50 лет степные участки малой площади полностью исчезли, а размеры крупных сократились на 80% [18].
Для района Ильменского заповедника выявлено возрастание среднегодовой температуры воздуха и годового количества осадков за последнее столетие, особенно заметное с 1970-х годов. Основные климатические изменения произошли в холодный период года, что согласуется с данными, полученными в южных районах Южного Урала [12, 13]. Согласно многолетним данным, полученным к концу 1950-х годов, в районе исследований весной и в начале лета нередки засухи и возвраты холодов [33]. Поэтому повышение эффективной температуры и количества осадков в апреле–июне 2000–2018 гг. благоприятно для степных сообществ. Особое значение имеет повышение количества осадков. Исследуемые экстразональные степи существуют на крутых склонах южных экспозиций, температура поверхности которых в летние месяцы может достигать 70°C [31], а поверхностный сток и интенсивное испарение приводят к дефициту влаги в жаркий период, при этом единственный источник увлажнения в данном местообитании – атмосферные осадки. В связи с этим обоснован вопрос, повлияли ли произошедшие климатические изменения непосредственно на степные сообщества, в том числе на наиболее ксеротермные из них, и как именно?
В результате 20-летних исследований обнаружены изменения в структуре сообществ петрофитных степей, в то время как луговые степи и остепненные луга не претерпели существенных изменений. Наблюдения на постоянных пробных площадях в сообществах петрофитных степей не выявили направленных изменений видового богатства и видовой насыщенности. Число видов в сообществе по годам изменялось в среднем на 12.7%, что подтверждает сведения о невысоком межгодовом варьировании видового состава травяных сообществ серпентинитов и известняков [4, 14, 15]. Показано [14], что видовая насыщенность травяных сообществ на серпентинитах в Калифорнии мало зависит от межгодовой изменчивости увлажнения. Это согласуется с представлением об устойчивости сообществ, формирующихся в местообитаниях с ограниченными ресурсами, к климатическим изменениям, включая прогнозируемое увеличение изменчивости осадков [3, 4]. В то же время исследование растительных сообществ на серпентинитах в штате Орегон с интервалом в 57 лет показало, что в условиях происходящих климатических изменений меняется не только обилие видов, но и видовой состав [1].
Ранее нами уже было обнаружено [18], что в условиях лесной зоны петрофитные степи очень чувствительны к увеличению увлажнения. Даже растительность наиболее сухих каменистых взлобков претерпевает сукцессионные смены, в ходе которых значительно уменьшается участие в сложении сообщества видов степного и лесостепного флористического комплекса и происходит смена доминантов – ксерофильных степных видов на мезофильные лесные.
Прямые наблюдения на территории Ильменского заповедника подтверждают, что возрастание количества осадков значительно меняет структуру петрофитных степей. Нами получены данные об увеличении ПП лугово-степных и степных видов за 20-летний период, что в совокупности с заменой петрофитно-степных доминирующих видов лугово-степными у части сообществ свидетельствует о происходящей мезофитизации исследованных петрофитных степей. Постепенно возрастает гомогенность сообществ, их горизонтальная структура становится более однородной. При этом внедрения новых видов не происходит, а увеличивается активность видов, уже имеющихся в сообществе.
Очень вероятно, что увеличение активности лугово-степных видов в петрофитных степях Ильменского заповедника связано с возрастанием увлажнения в апреле–июне за последние 20 лет. Известно, что в условиях экстразональных степей именно лугово-степные виды (Fillipendula vulgaris, Fragaria viridis, Stipa pennata и др.) особенно чутко реагируют на увеличение влагообеспеченности – их надземная фитомасса возрастает во влажные годы и снижается в засушливые [34]. Это также подтверждают результаты 12-летних наблюдений в южной Польше за реакцией остепненного луга на изменение климатических параметров: установлено, что ПП большинства видов и функциональных групп увеличивалось в ответ на возрастание осадков апреля–июня в год наблюдений [35]. Данные, свидетельствующие о мезофитизации экстразональных петрофитных степей на Южном Урале за последние 20 лет, совпадают с данными о возрастании продуктивности экстразональных луговых степей Приокско-Террасного заповедника в 1998–2011 гг. по сравнению с 1975–1985 гг. на 45%; при этом рост продуктивности происходил на фоне увеличения среднегодовой температуры воздуха, суммы эффективных температур и количества осадков за вегетационный период [16].
Определяющее значение для существования экстразональных сообществ, представляющих экологические изоляты, имеет размер местообитания. Так, для остепненной растительности диоритовых выходов различной площади в Центральной Германии показано [36], что крупные обнажения характеризуются более стабильными условиями, меньшими проявлениями краевого эффекта, что выражается в высоком числе редких растений и большем фитоценотическом разнообразии. В случае исследованных нами петрофитных сообществ от площади, которую они занимают, зависела степень мезофитизации – для ключевых сообществ малой площади отмечено наибольшее возрастание ПП лугово-степных и степных видов. В основном сообщества малой площади приурочены к коротким степным склонам, которые большую часть дня затенены окружающим лесом. Значительное влияние лесной растительности на прилегающие степные фитоценозы показано как в степной, так и в лесостепной зонах: вблизи лесного массива по сравнению с окружающей степью создается относительно влажный и менее континентальный мезоклимат [37, 38]. Ключевые сообщества длинных профилей имеют бóльшую площадь, находятся в более стабильных условиях, и изменения соотношения видов разных ценотических групп выражены здесь в меньшей степени.
ЗАКЛЮЧЕНИЕ
Наиболее надежные результаты исследования динамики растительности можно получить с помощью прямых многолетних наблюдений на постоянных пробных площадях. Используя этот подход, мы впервые показали изменение структуры экстразональных степей на фоне происходящих климатических изменений. Получены убедительные доказательства постепенной мезофитизации наиболее ксерофитных сообществ бореальной зоны Южного Урала в период 1999–2019 гг. С нашей точки зрения, возрастание гомогенности, а также участия в сложении сообщества лугово-степных и степных видов, замещение ими на доминирующих позициях петрофитно-степных видов связано с увеличением количества осадков в начале вегетационного сезона.
Таким образом, наблюдаемые изменения в первую очередь коснулись горизонтальной и ценотической структуры сообществ петрофитных степей, видовой состав на данном этапе сукцессионных смен не изменился. Устойчивость исследованных сообществ к климатическим изменениям напрямую связана с их площадью – растительность крупных участков более стабильна. Исторические фотографии и почвенные исследования [32] свидетельствуют о том, что соотношение и распределение степной и лесной растительности в районе Демидовских сопок неоднократно менялись, в связи с чем экстразональные степные участки представляют большую ценность в качестве геоботанического репера для выявления динамических тенденций современного растительного покрова.
Работа выполнена в рамках государственного задания Института экологии растений и животных УрО РАН № АААА-А19-119031890084-6. Автор признателен докт. биол. наук Д.В. Веселкину и канд. биол. наук Е.Н. Подгаевской (Институт экологии растений и животных УрО РАН) за конструктивное обсуждение рукописи, канд. биол. наук Н.Б. Куянцевой (Ильменский государственный заповедник) и канд. биол. наук Е.Н. Подгаевской – за помощь в организации полевых работ.
Список литературы
Damschen E.I., Harrison S.P., Grace J.B. Climate change effects on an endemic-rich edaphic flora: resurveying Robert H. Whittaker’s Siskiyou sites (Oregon, USA) // Ecology. 2010. V. 91. № 12. P. 3609–3619.
Damschen E.I., Harrison S., Ackerly D.D. et al. Endemic plant communities on special soils: early victims or hardy survivors of climate change? // J. Eco. 2012. V. 100. № 5. P. 1122–1130.
Grime J.P., Brown V.K., Thompson K. et al. The response of two contrasting limestone grasslands to simulated climate change // Science. 2000. V. 289. № 5480. P. 762–765.
Grime J.P., Fridley J.D., Askew A.P. et al. Long-term resistance to simulated climate change in an infertile grassland // Proceedings of the National Academy of Sciences USA. 2008. V. 105. № 29. P. 10028–10032.
Porro F., Tomaselli M., Abeli T. et al. Could plant diversity metrics explain climate driven vegetation changes on mountain summits of the GLORIA network? // Biodiversity and Conservation. 2019. № 28. P. 3575–3596.
Sternberg M., Brown V.K., Masters G.J., Clarke I.P. Plant community dynamics in a calcareous grassland under climate change manipulations // Plant Ecology. 1999. № 143. P. 29–37.
Горчаковский П.Л. Основные проблемы исторической фитогеографии Урала. Свердловск: УФАН СССР, 1969. 285 с.
Куликов П.В. Конспект флоры Челябинской области. Екатеринбург–Миасс: “Геотур”, 2005. 537 с.
Красная книга Челябинской области: Животные. Растения. Грибы. Изд. 2-е. М.: “Реарт”, 2017. 511 с.
Горчаковский П.Л., Золотарева Н.В. Реликтовая степная растительность Ильменских гор на Южном Урале. Екатеринбург: Гощицкий, 2004. 117 с.
Zolotareva N., Korolyuk A. Extrazonal steppes of forest belt on eastern macroslope of the Urals // Results and Prospects of Geobotanical Research in Siberia. BIO Web of Conferences. 2019. № 16, 00043. https://doi.org/10.1051/bioconf/20191600043 (Electronic resourse).
Агафонов Л.И., Кукарских В.В. Изменения климата прошлого столетия и радиальный прирост сосны в степи Южного Урала // Экология. 2008. № 3. С. 173–180. [Agafonov L.I., Kukarskikh V.V. Climate changes in the past century and radial increment of pine in the Southern Ural steppe // Rus. J. Ecol. 2008. V. 39. № 3. P. 160–167.]
Шкляев В.А., Шкляева Л.С. Оценка изменений температуры воздуха и осадков Среднего и Южного Урала в XX веке // Вестн. Челябинского гос. ун-та. 2011. № 5(220). С. 61–69.
Fernandez-Going B.M., Anacker B.L., Harrison S.P. Temporal variability in California grasslands: Soil type and species functional traits mediate response to precipitation // Ecology. 2012. V. 93. № 9. P. 2104–2114.
Cleland E.E., Collins S.L., Dickson T.L. et al. Sensitivity of grassland plant community composition to spatial vs. temporal variation in precipitation // Ecology. 2013. V. 94. № 8. P. 1687–1696.
Зеленская Н.Н., Керженцев А.С., Аблеева В.А., Терешонок Н.А. Динамика продуктивности луговых степей на северо-западном пределе их ареала (бассейна Оки) // Изв. РАН. Серия географич. 2012. № 6. С. 47–54.
Eurasian Steppes. Ecological problems and livelihoods in a changing world / Werger M.J.A., van Staalduinen M.A. Eds. New York, London: Springer, Dordrecht, Heidelberg, 2012. 565 p.
Золотарева Н.В., Золотарев М.П. Феномен облесения степных участков на Среднем Урале и его вероятные причины // Экология. 2016. № 6. С. 414–425. [Zolotareva N.V., Zolotarev M.P. The phenomenon of forest invasion to steppe areas in the Middle Urals and its probable causes // Rus. J. of Ecology. 2017. V. 48. № 1. P. 21–31.] https://doi.org/10.7868/S0367059716060159
Баландин С.В. Динамика степной растительности Уктусских гор (Средний Урал) // Ботан. журн. 2001. Т. 86. № 5. С. 103–110.
Сизых А.П. Модели таежно-степных сообществ западного побережья оз. Байкал // Экология. 2007. № 4. С. 255–258. [Sizykh A.P. Models of taiga-steppe communities on the western coast of Lake Baikal // Rus. J. of Ecology. 2007. V. 38. № 4. P. 234–237.]
Шакиров А.В. Физико-географическое районирование Урала. Екатеринбург: УрО РАН, 2011. 617 с.
Батчаева О.М., Захаров А.А., Онипченко В.Г. Зависимость распространения растений от глубины почвы в альпийских фитоценозах Тебердинского заповедника // Бюл. МОИП. Отд. биол. 2003. Т. 108. № 4. С. 53–57.
Золотарева Н.В., Королюк А.Ю., Ямалов С.М. Cообщества класса Festuco-Brometea Br.-Bl. et Tx. ex Soó 1947 Месягутовской и Красноуфимской лесостепей (Среднее Предуралье) // Растительность России. 2019. № 37. С. 29–76.
Hammer Ø., Harper D.A.T., Ryan P.D. PAST: Paleontological statistics software package for education and data analysis // Palaeontologia Electronica. 2001. V. 4. № 1. P. 1–9.
Рязанова Л.В. Конспект флоры степного юга Челябинской области. Челябинск: ЧГПУ, 2006. 445 с.
Дорогостайская Е.В. Конспект флоры цветковых растений Ильменского заповедника // Тр. Ильменского гос. заповедника. 1961. Вып. 8. С. 9–50.
Шенников А.П. Введение в геоботанику. Л.: Изд-во ЛГУ, 1964. 447 с.
Методы дендрохронологии. Ч. I. Основы дендрохронологии. Сбор и получение древесно-кольцевой информации: Учеб.-метод. пос. / Шиятов С.Г., Ваганов Е.А., Кирдянов А.В. и др. Красноярск: КрасГУ, 2000, 80 с.
Булыгина О.Н., Веселов В.М., Разуваев В.Н., Александрова Т.М. Описание массива срочных данных об основных метеорологических параметрах на станциях России. Свидетельство о государственной регистрации базы данных № 2014620549. http://meteo.ru/data/163-basic-parameters#описание-массива-данных
Шиятов С.Г. Динамика древесной и кустарниковой растительности в горах Полярного Урала под влиянием современных изменений климата. Екатеринбург: УрО РАН, 2009. 216 с.
Жирнова Т.В., Ямалов С.М., Миркин Б.М. Степи Башкирского государственного природного заповедника: анализ вклада ведущих факторов и синтаксономия // Бюл. МОИП. Отд. биол. 2007. Т. 112. №. 5. С. 36–45.
Тюлина Л.Н. К эволюции растительного покрова Восточных предгорий Южного Урала // Зап. Златоустовского общ-ва краеведения. Златоуст, 1928. Вып. 1. 186 с.
Агроклиматический справочник по Челябинской области. Л.: Гидрометеоиздат, 1960. 156 с.
Данилов В.И. О некоторых вопросах погодичной динамики степных фитоценозов // Бюл. МОИП. Отд. биол. 1981. Т. 86. № 5. С. 106–120.
Dzwonko Z., Loster S. Impact of weather on dynamics of plant functional groups in an abandoned limestone grassland // Acta Soc. Bot. Pol. 2011. № 80(4). P. 259–268.
Wesche K., Partzsch M., Krebes S., Hensen I. Gradients in dry grassland and heath vegetation on rock outcrops in Eastern Germany – an analysis of a large phytosociological data set // Folia Geobotanica. 2005. № 40. P. 341–356.
Русанов А.М. Влияние леса на состав и свойства органического вещества степных черноземов прилегающих биогеоценозов // Экология. 2012. № 1. С. 36–42.
Erdős L., Tölgyesi C., Horzse M. et al. Habitat complexity of the Pannonian forest-steppe zone and its nature conservation implications // Ecological Complexity. 2014. V. 17. P. 107–118.
Дополнительные материалы отсутствуют.