

Экология, 2021, № 4, стр. 243-253
Результат воздействия доминантов на видовое богатство растительных сообществ: упорядоченное или случайное исчезновение видов?
В. В. Акатов a, *, Т. В. Акатова b, Д. Ф. Афанасьев c, Т. Г. Ескина b, Е. Г. Сушкова c, С. Г. Чефранов a
a Майкопский государственный технологический университет
385000 Майкоп, ул. Первомайская, 191, Россия
b Кавказский государственный природный биосферный заповедник
385000 Майкоп, ул. Советская, 187, Россия
c Азово-Черноморский филиал Всероссийского научно-исследовательского института рыбного хозяйства
и океанографии (“АзНИИРХ”)
344002 Ростов-на-Дону, ул. Береговая, 21б, Россия
* E-mail: akatovmgti@mail.ru
Поступила в редакцию 20.03.2020
После доработки 17.01.2021
Принята к публикации 25.01.2021
Аннотация
Рассмотрен вопрос о характере (случайном или упорядоченном) исчезновения видов в растительных сообществах в результате роста участия доминантов. В качестве объектов изучения были выбраны 15 участков сообществ разных типов: прирусловых отмелей рек, лесных полян, степей, субальпийских и альпийских лугов Западного Кавказа, макрофитобентоса Азовского моря. Использованы два подхода: 1-й – данные по пробам биомассы, отобранным с микроучастков сообществ с разным участием доминирующего вида, были сопоставлены с сериями модельных ценозов со случайным распределением видов; 2-й – сравнили группы проб с разным участием доминанта, но благодаря разному числу проб в группах с равной суммарной биомассой сопутствующих видов. В результате хорошо выраженный эффект упорядоченного исчезновения видов был выявлен только на трех участках (с доминированием Rubus caesius, Glycyrrhiza glabra и Solidago сanadensis). В остальных случаях распределение видов по микроучасткам с разным участием доминанта имело случайный характер. Это означает, что рост участия доминантов ведет преимущественно к неизбирательному вытеснению других видов.
Считается, что доминирующие виды существенно ограничивают локальное видовое богатство растительных сообществ, по крайней мере в стабильных продуктивных местообитаниях [1–8], но не ясно, способны ли они влиять на видовое богатство крупных участков растительного покрова, а значит, при усилении их позиций представлять угрозу фиторазнообразию регионов в целом [9, 10]. Сделано предположение [11–13], что это может зависеть от степени различия видов растений по устойчивости к данному фактору. Если преимущества одних видов перед другими выражены хорошо, то рост степени доминирования на микроучастках ценозов будет сопровождаться исключением из них главным образом одних и тех же (менее устойчивых) видов растений (упорядоченное вымирание – [14, 15]), что приведет к их исчезновению на крупных участках сообществ. Если же такие преимущества выражены слабо, то рост участия доминантов приведет к исключению из микроучастков главным образом разных видов (случайное вымирание – [14, 15]), что не окажет значительного влияния на видовое богатство сообществ в целом (размер их видового пула), хотя снизит встречаемость сопутствующих видов и соответственно может сделать их более уязвимыми к воздействию других факторов, например фитофагов, флуктуаций среды, фрагментации и т.д.
Так преимущественно случайно или упорядоченно исчезают виды растений с микроучастков растительных сообществ при усилении позиции доминантов? На эту проблему обратили внимание лишь в последние годы, а в качестве объектов изучения были использованы в основном чужеродные доминанты и растительные сообщества, наиболее часто подвергающиеся заселению такими видами [2, 9–13, 16]. Мы рассмотрели этот вопрос на примере фитоценозов разных типов и преимущественно аборигенных доминантов.
МАТЕРИАЛ И МЕТОДИКА
Район и объекты исследования. Район исследования включал предгорье и горные массивы Западного Кавказа (бассейны рек Белая и Абин), западную часть Кубано-Приазовской низменности, мелководный участок Таманского залива Азовского моря. Объектом изучения были выбраны 15 однородных участков растительных сообществ с хорошо выраженным доминированием определенного вида, участие которого тесно связано с видовым богатством на микроплощадках: 1) три участка сообществ альпийских лугов с доминированием Geranium gymnocaulon, Kobresia capillifolia и Alchemilla retinervis (бассейн р. Белой, г. Оштен, 2230−2400 м над ур. м.); 2) три – субальпийских лугов с доминированием Inula grandiflora, Calamagrostis arundinacea и Alchemilla persica (бассейн р. Белой, г. Оштен, 2015−2025 м); 3) четыре – нижнегорных лесных полян с доминированием Botriochloa ischaemum и Calamagrostis epigeios (бассейн р. Белой, хр. Скалистый, 500 и 797 м), Solidago сanadensis и Rubus caesius (террасы р. Белой, 192 и 204 м); 4) один – горных степей с доминированием Agropyron pinifolium (бассейн р. Абин, г. Шизе, 530 м); 5) один – сообщества береговой зоны соленого лимана с доминированием Glycyrrhiza glabra (дельта р. Кубань, окрестности хут. Прорвенский, 1 м); 6) два – сообщества старой прирусловой отмели р. Белой с доминированием Echinochloa crus-galli и Setaria viridis (189 и 196 м); 7) один – макрофитобентос горизонта фотофильной растительности сублиторали Таманского залива Азовского моря с доминированием Zostera noltei (глубина 1.0−2.5 м).
В соответствии с полимодельной концепцией Б.М. Миркина [4] сообщества альпийских, субальпийских и нижнегорных лугов могут быть отнесены к CRS-модели организации [4, 17, 18]. Они сформированы в относительно стабильных и продуктивных местообитаниях и характеризуются доминированием многолетних конкурентно мощных видов. Из них Botriochloa ischaemum и Calamagrostis epigeios хорошо известны своей способностью ограничивать видовое богатство ценозов на небольших участках. Считается, что они сокращают ресурсы и пространство, доступные для других видов, а также ограничивают их проникновение в сообщества путем накопления мощного слоя ветоши [3, 6]. Agropyron pinifolium – многолетний злак, стерильные побеги которого образуют плотную дерновину [19]. Чужеродный вид Solidago сanadensis (родина Северная Америка) способен образовывать густые заросли с плотностью побегов более 300 экз/м2. Растение выделяет дитерпеноиды, которые ингибируют рост сеянцев других видов [20, 21]. Glycyrrhiza glabra также образует густые заросли высотой от 0.8 до 1.5 м, а Rubus caesius – многолетний полукустарник со стелющимися по субстрату и укореняющимися побегами, формирующими над его поверхностью плотный полог.
Сообщества макрофитобентоса с доминированием высших водных растений (Zostera noltei) формируются на песчаных и илистых отложениях в олигосапробных условиях, при относительно невысокой освещенности, испытывают достаточно мощное волновое изреживание. Сообщества прирусловых отмелей рек сформированы на низкопродуктивном субстрате (песок, галька) и характеризуются высокой частотой нарушений. Ценозы обоих типов могут быть отнесены к RS-модели организации. При этом проективное покрытие доминирующих видов в них местами достигает 80−90%.
Методы сбора и анализа фактического материала. В пределах однородных участков наземных сообществ с доминированием определенного вида было заложено по 25−30 площадок размером 0.5 × 0.5 м, в пределах морского участка – 20 таких же площадок. Часть площадок была заложена регулярным способом в виде одной или двух трансект, включающих 10 площадок, другие – сериями по 5−10 шт. на участок. Во втором случае выбирали варианты микроучастков сообществ с высоким и низким проективным покрытием доминирующих видов, которое оценивали визуально. Отобранные с каждой площадки пробы биомассы были разобраны по видам и взвешены. Для каждой пробы были определены степень доминирования (относительная биомасса доминирующего вида – D), сухая биомасса сопутствующих видов (Ws) и число видов (S). Тесноту связи между значениями этих показателей в сериях проб определяли путем расчета коэффициента корреляции рангов Спирмена. Общая характеристика участков сообществ приведена в табл. 1.
Таблица 1.
Характеристика участков растительных сообществ
| Доминанты (константные виды) | n | N | Smin−Smax | Dmin−Dmax | r |
|---|---|---|---|---|---|
| Alchemilla retinervis (Myosotis alpestris, Cirsium simplex, Phleum alpinum) | 25 | 28 | 3−16 | 0.24−0.99 | −0.788 |
| Kobresia capillifolia (Festuca ovina, Carex tristis, Polygonum carneum) | 25 | 28 | 4−14 | 0.23−0.96 | −0.839 |
| Geranium gymnocaulon (Rumex arifolius, Phleum alpinum, Ranunculus caucasicus) | 25 | 43 | 9−23 | 0.13−0.93 | −0.495 |
| Calamagrostis arundinacea (Vicia grossheimii, Betonica macrantha) | 30 | 39 | 5−17 | 0.20−0.94 | −0.565 |
| Inula grandiflora (Chaerophyllum roseum, Campanula latifolia, Poa longifolia) | 30 | 26 | 5−14 | 0.18−0.92 | −0.645 |
| Alchemilla persica (Betonica macrantha, Pimpinella rhodantha, Centaurea abbreviata) | 30 | 58 | 16−32 | 0.21−0.81 | −0.733 |
| Botriochloa ischaemum (Dorycnium intermedium, Lotus corniculatus, Polygala anatolica) | 25 | 48 | 6−17 | 0.23−0.93 | −0.647 |
| Calamagrostis epigeios (Carex tomentosa, Poa pratensis, Lathyrus pratensis) | 25 | 31 | 3−14 | 0.30−0.93 | −0.612 |
| Rubus caesius (Poa angustifolia, Calystegia silvatica, Coronilla varia) | 30 | 28 | 4−13 | 0.14−0.98 | −0.630 |
| Solidago сanadensis (Elytrigia repens, Daucus carota, Carex contigua) | 25 | 33 | 4−16 | 0.28−0.98 | −0.661 |
| Agropyron pinifolium (Convolvulus cantabrica, Sideritis montana, Allium saxatile) | 30 | 34 | 5−13 | 0.26−0.94 | −0.733 |
| Glycyrrhiza glabra (Elytrigia elongata, Asperula humifusa, Achillea nobilis) | 25 | 18 | 2−11 | 0.34−0.77 | −0.593 |
| Echinochloa crus-galli (Melilotus hirsutus, Setaria viridis, Polygonum persicaria) | 25 | 36 | 2−14 | 0.63−0.99 | −0.616 |
| Setaria viridis (Medicago lupulina, Ambrosia artemisiifolia, Melilotus hirsutus) | 25 | 21 | 2−9 | 0.45−0.98 | −0.647 |
| Zostera noltei (Zostera marina, Pneophyllum fragile, Cladophora albida) | 20 | 18 | 2−7 | 0.50−0.99 | −0.602 |
Примечание. n – число проб; N – общее число видов в серии проб; Smin−Smax – минимальное и максимальное число видов растений на микроучастках 0.25 м2 (в пробах); Dmin−Dmax– минимальное и максимальное значения степени доминирования; r – значения коэффициента корреляции рангов Спирмена между D и S (все значения статистически значимы на уровне Р < < 0.05). Названия видов сосудистых растений приводятся по А.И. Галушко [35], макроводорослей – согласно глобальной базе данных по таксономии и номенклатуре водорослей AlgaeBase [36].
Преобладающий характер воздействия (избирательный или неизбирательный) определенного доминанта на сопутствующие виды оценивали на основе двух подходов. В соответствии с первым для каждой серии проб, отобранной на участке растительного сообщества, были сформированы по три серии модельных ценозов со случайным распределением видов. Их построение производилось в среде Excel из пакета Microsoft Office версии 2013. Каждый модельный ценоз представлял собой бинарный вектор размера N (общее число видов, выявленных в пробах соответствующей серии). Каждый элемент вектора принимает значение 1 (присутствие определенного вида) или 0 (его отсутствие). Выбор конкретного значения осуществлялся в соответствии с формулой
(1)
$y\left( x \right) = \left\{ {\begin{array}{*{20}{c}} {1,{\text{\;если}}\,\,{\text{\;}}0 \leqslant x < Fr,} \\ {0,{\text{\;если\;}}Fr \leqslant x \leqslant 1,{\text{\;}}} \end{array}} \right.$Поскольку сопутствующие виды растений с низкой устойчивостью к воздействию доминанта не могут иметь высокую встречаемость в сериях проб биомассы, объектом тестирования стали виды со средней и низкой встречаемостью. Мы предположили, что в случае разной устойчивости этих видов к воздействию доминанта и преимущественно упорядоченного их исчезновения в ответ на рост его участия большая часть из них будет встречаться в пробах биомассы с более низкой степенью доминирования (пределы их распространения на градиенте D будут расположены преимущественно в области меньших значений), чем это можно было бы ожидать, если бы их распределение по пробам определялось случайными процессами, т.е. как в сериях модельных ценозов. Если распределение видов по пробам определяется случайными процессами, то соотношение числа случаев, когда они были бы обнаружены в пробах с более низким и более высоким участием доминанта, чем в модельных ценозах, должно быть близко к 1 : 1.
Сравнение серий природных и модельных ценозов осуществляли в следующей последовательности: 1) пробы каждой серии и соответствующие им модельные ценозы были ранжированы по росту участия доминирующего вида; 2) для каждого сопутствующего вида со средней и низкой встречаемостью (условно менее 0.6) определено наиболее высокое (пороговое) значение степени доминирования в пробах с его присутствием (DMF) и три аналогичных значения для модельных серий (DME); 3) для каждого вида определены три разности между значениями этих показателей (DME – DMF = ∆DM); 4) используя критерий χ2, оценена статистическая значимость отклонения соотношения числа положительных и отрицательных значений ∆DM для рассматриваемых видов от 1 : 1. Если данное отклонение оказывалось статистически незначимым, то соотношение положительных и отрицательных значений ∆DM оценивалось только для видов с низкой встречаемостью (менее 0.3).
Для проверки эффективности данного метода мы дополнительно сформировали 48 серий по 25 модельных ценозов. В основу формирования 12 из них был заложен механизм, предполагающий упорядоченное исчезновение видов в ответ на рост степени доминирования: 1) снижение по этой причине видового богатства ценозов (S) сопровождается снижением размера их видового пула (N) условно в 3, 2, 1.5 и 1.25 раза; 2) более крупные пулы полностью включают виды меньших пулов; 3) видовой состав ценозов с заданными значениями S является случайной выборкой из соответствующих им по размеру пулов (N). Максимальные значения N в модельных сериях (18, 31 и 48 видов) соответствовали общему числу видов растений в природных сериях с относительно низким, средним и высоким видовым богатством.
Для определения видового состава ценозов (набора значений 1 и 0 в таблице размером 1 × N) использовался генератор случайных чисел, описываемый функцией
(2)
$y\left( x \right) = \left\{ {\begin{array}{*{20}{c}} {1,{\text{\;если\;}}\,\,0 \leqslant x < S{\text{/}}N,} \\ {0,~{\text{если\;}}\,\,S{\text{/}}N \leqslant x \leqslant 1,~} \end{array}} \right.$Для каждого варианта серии модельных ценозов, сформированных на основе предположения об упорядоченном исчезновении видов в ответ на рост степени доминирования, используя алгоритм, описанный выше (1), были сгенерированы по три серии ценозов со случайным распределением видов. Сравнение серий модельных ценозов, сформированных на основе разных предположений, было выполнено таким же способом, как сравнение природных серий с модельными, т.е. путем анализа значений ∆DM, рассчитанных для видов со встречаемостью менее 0.6.
В основу второго подхода были положены представления энергетической (energy-diversity) гипотезы [22, 23]. В соответствии с ней количество доступной энергии (продуктивность местообитаний) может ограничивать число сосуществующих видов путем лимитирования плотности их особей. Это предполагает, что сообщества со сходной продуктивностью должны характеризоваться близким видовым богатством. В качестве суррогата показателя продуктивности сообществ недревесных видов растений обычно используют надземную сухую биомассу в период ее максимального развития.
Из данной гипотезы также следует, что чем значительнее участие доминирующих видов в формировании фитоценозов, тем меньше ресурсов остается другим (сопутствующим) видам, тем ниже их суммарная биомасса и соответственно меньшее число их особей оказывается на участках, тем ниже видовое богатство сообществ. В случае, если соотношение между участием доминанта и числом сопутствующих видов определяется только данным механизмом (неизбирательное вытеснение видов), вероятность присутствия каждого вида сообщества в группах проб с разным участием этого доминанта, но с равной суммарной биомассой сопутствующих видов (благодаря разному числу проб в группах) должна быть одинаковой. Соответственно соотношение числа видов сообщества, обнаруженных и необнаруженных в данных группах проб (с низким и высоким участием доминанта) также должно быть примерно одинаковым. Напротив, если часть видов сообщества не способна произрастать на участках с высоким участием доминанта (например, из-за значительного изменения им условий произрастания: сильного затенения, мощного слоя ветоши и т.д.), то число видов, обнаруженных на таких участках, должно быть меньше, а необнаруженных – больше, чем на участках с низким его участием, но со сходной суммарной биомассой.
Процедура оценки степени правомерности данных предположений включала следующие действия: 1) для каждого из 15 участков сообществ была сформирована группа из пяти проб биомассы с наиболее низким участием доминанта и для каждой из них определена суммарная биомасса сопутствующих видов; 2) сформирована группа проб с наиболее высоким участием доминанта, в которой суммарная сухая биомасса сопутствующих видов примерно соответствовала значениям этой характеристики в группе проб с низким его участием; 3) соотношение числа видов, обнаруженных и необнаруженных в пробах с низким участием доминанта, рассматривалось как ожидаемое для проб с высоким его участием. Статистическую значимость различий этих соотношений оценивали с использованием критерия χ2.
РЕЗУЛЬТАТЫ
Как видно из табл. 1, во всех сериях проб биомассы, отобранных на изученных участках растительных сообществ, относительная биомасса доминирующих видов (степень доминирования) и число видов в пробах варьируют в широких пределах. При этом связь между значениями этих характеристик на большинстве участков является высокой и во всех случаях статистически значимой.
Результаты сопоставления границ распространения видов на градиенте степени доминирования (D) в сериях модельных сообществ, сформированных на основе предположения об упорядоченном исчезновении видов, и в соответствующих им сериях модельных сообществ, сформированных на основе предположения о случайном исчезновении видов, приведены в табл. 2. Из нее следует, что в случае упорядоченного исчезновения видов с ростом участия доминанта большая часть видов со встречаемостью в сериях менее 0.6 или 0.3 обнаруживается в ценозах с более низкой степенью доминирования (в области меньших значений на градиенте D), чем в модельных сериях со случайным распределением видов (т.е. значения ∆DM являются преимущественно положительными). Причем если рост степени доминирования сопровождается трех- или двукратным снижением размера видового пула, то данное различие во всех случаях является статистически достоверным для P < 0.05; при снижении размера пула в 1.5 раза – в ценозах с высоким и средним видовым богатством (максимальный размер пулов равен 48 и 31 видам соответственно), в 1.25 раза – только с высоким. Рис. 1 иллюстрирует полученный результат на примере серий модельных ценозов 9−12 (см. табл. 2).
Таблица 2.
Результаты сопоставления границ распространения видов на градиенте степени доминирования (D) в сериях модельных сообществ, сформированных на основе предположения об упорядоченном исчезновении видов, и в соответствующих им сериях модельных сообществ, сформированных на основе предположения о случайном исчезновении видов
| № | Nmax –Nmin | Smin−Smax | F | n | Число (доля) случаев | χ2 | |
|---|---|---|---|---|---|---|---|
| ∆DM > 0 | ∆DM < 0 | ||||||
| 1 | 18–6 | 9–4 | <0.6 | 15 | 28(0.70) | 12(0.30) | 6.40* |
| 2 | 18–9 | 9–4 | <0.3 | 6 | 15(0.94) | 1(0.06) | 12.25* |
| 3 | 18–12 | 9–4 | <0.3 | 7 | 11(0.65) | 6(0.35) | 1.47 |
| 4 | 18–14 | 9–4 | <0.3 | 7 | 13(0.65) | 7(0.35) | 1.80 |
| 5 | 31–11 | 16–8 | <0.6 | 25 | 56(0.85) | 10(0.15) | 32.06* |
| 6 | 31–16 | 16–8 | <0.3 | 11 | 21(0.78) | 6(0.22) | 8.33* |
| 7 | 31–21 | 16–8 | <0.3 | 11 | 21(0.72) | 8(0.28) | 5.83* |
| 8 | 31–25 | 16–8 | <0.3 | 10 | 16(0.57) | 12(0.43) | 0.57 |
| 9 | 48–16 | 25–12 | <0.6 | 40 | 79(0.72) | 30(0.28) | 22.03* |
| 10 | 48–24 | 25–12 | <0.6 | 43 | 74(0.65) | 39(0.35) | 10.84* |
| 11 | 48–32 | 25–12 | <0.6 | 43 | 61(0.64) | 35(0.36) | 7.04* |
| 12 | 48–38 | 25–12 | <0.3 | 9 | 21(0.78) | 6(0.22) | 8.33* |
Примечание. № – номер серии модельных сообществ, сформированных на основе предположения об упорядоченном исчезновении видов; Nmax–Nmin и Smin−Smax – заданные пределы варьирования размера видового пула и видового богатства сообществ в данной серии (предполагалось, что варьирование размера видового пула и видового богатства обусловлено произвольно заданным ростом степени доминирования от 0.20 до 0.92); F – встречаемость видов; n – число видов с определенной встречаемостью; ∆DM = DME – DMOR, где DMOR – наиболее высокое значение степени доминирования в модельных ценозах с присутствием определенного вида, сформированных на основе предположения об упорядоченном исчезновении видов, DME – аналогичное значение в сериях модельных ценозов со случайным распределением видов; * – фактические значения χ2 превышают критические для P < 0.05.
Рис. 1.
Соотношение между встречаемостью видов в модельных сериях (F) и значениями показателя ∆DM: ∆DM = = DME – DMOR, где DMOR – наиболее высокое значение степени доминирования в модельных ценозах с присутствием определенного вида, сформированных на основе предположения об упорядоченном исчезновении видов; DME – аналогичное значение в сериях модельных ценозов со случайным распределением видов; а, б, в, г – рост степени доминирования сопровождается снижением размера видового пула в 3, 2, 1.5 и 1.25 раза (серии модельных ценозов 9, 10, 11 и 12 в табл. 2).
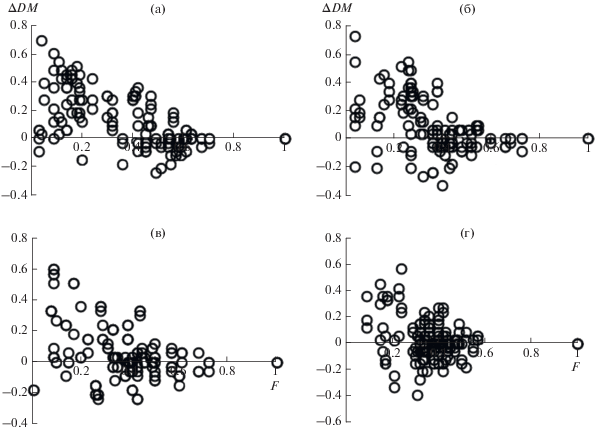
Таким образом, результаты табл. 2 свидетельствуют о том, что если рост участия доминанта ведет к снижению размера видового пула более чем в 1.25, 1.5 и 2 раза в ценозах с относительно высоким, средним и низким видовым богатством соответственно, то этот эффект может быть выявлен предложенным способом. Результаты его использования применительно к 15 участкам сообществ с доминированием определенных видов приведены в табл. 3, а в качестве иллюстрации для некоторых из них – на рис. 2. Видно, что в сериях проб с доминированием Rubus caesius и Glycyrrhiza glabra для видов со встречаемостью менее 0.6, а также с доминированием Solidago сanadensis для видов со встречаемостью менее 0.3 число положительных значений ∆DM выше, чем отрицательных, причем их соотношение статистически значимо отличается от 1 : 1 (P < 0.05).
Таблица 3.
Результаты сопоставления границ распространения видов на градиенте роста степени доминирования (D) в сериях проб биомассы и модельных ценозов, сформированных на основе предположения о случайном исчезновении видов
| Доминанты | F | n | Число (доля) случаев | χ2 | |
|---|---|---|---|---|---|
| ∆DM > 0 | ∆DM < 0 | ||||
| Alchemilla retinervis | <0.3 | 18 | 21(0.47) | 24(0.53) | 0.20 |
| Kobresia capillifolia | <0.3 | 20 | 30 (0.63) | 18(0.37) | 3.00 |
| Geranium gymnocaulon | <0.3 | 22 | 31(0.57) | 23(0.43) | 1.19 |
| Calamagrostis arundinacea | <0.3 | 29 | 42(0.58) | 30(0.42) | 2.00 |
| Inula grandiflora | <0.3 | 12 | 13(0.48) | 14(0.52) | 0.04 |
| Alchemilla persica | <0.3 | 31 | 45(0.52) | 42(0.48) | 0.10 |
| Botriochloa ischaemum | <0.3 | 31 | 31(0.42) | 43(0.58) | 1.95 |
| Calamagrostis epigeios | <0.3 | 24 | 27(0.50) | 27(0.50) | 0.00 |
| Rubus caesius | <0.6 | 15 | 30(0.73) | 11(0.27) | 8.81* |
| Solidago canadensis | <0.3 | 25 | 40(0.63) | 23(0.37) | 4.59* |
| Agropyron pinifolium | <0.3 | 20 | 25(0.52) | 23(0.48) | 0.08 |
| Glycyrrhiza glabra | <0.6 | 12 | 25(0.76) | 8(0.24) | 8.76* |
| Echinochloa crus-galli | <0.3 | 26 | 26(0.44) | 33(0.56) | 0.83 |
| Setaria viridis | <0.3 | 15 | 17(0.45) | 21(0.55) | 0.42 |
| Zostera noltei | <0.3 | 15 | 13(0.38) | 21(0.62) | 1.88 |
Примечание. F – встречаемость видов в пробах; n – число видов с определенной встречаемостью (менее 0.6 или 0.3); ∆DM = DME – DMF, где DMF – наиболее высокое значение степени доминирования в пробах с присутствием определенного вида, DME – аналогичное значение в сериях модельных ценозов со случайным распределением видов; * – фактические значения χ2 превышают критические для P < 0.05.
Рис. 2.
Соотношение между встречаемостью видов в серии проб биомассы с доминированием определенного вида (F) и значениями показателя ∆DM: ∆DM = DME – DMF, где DMF – наиболее высокое значение степени доминирования в пробах с присутствием определенного вида; DME – аналогичное значение в сериях модельных ценозов со случайным распределением видов.
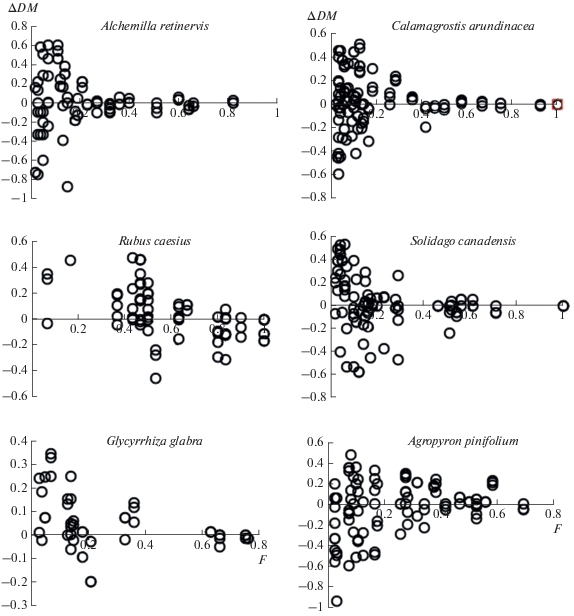
Похожая ситуация наблюдается на участках альпийских лугов с доминированием Kobresia capillifolia и Geranium gymnocaulon, а также субальпийских лугов с доминированием Calamagrostis arundinacea. Большая часть сопутствующих видов этих сообществ со встречаемостью менее 0.3 обнаруживается в пробах с более низкой степенью доминирования, чем в модельных сериях, однако наблюдаемые различия не достоверны для P < 0.05. В других сериях проб соотношение положительных и отрицательных значений ∆DM близко к 1 : 1, либо число отрицательных значений этого показателя несколько выше, чем положительных. Чтобы уточнить полученный результат для участков с доминированием K. capillifolia, G. gymnocaulon и C. arundinacea, мы дополнительно сгенерировали для каждого из них еще по три серии модельных ценозов и повторили процедуру тестирования. Однако, несмотря на рост числа ∆DM примерно в два раза, значения χ2 существенно не изменились. В частности, на участке с доминированием K. capillifolia для видов со встречаемостью менее 0.3 число положительных значений ∆DM оказалось равным 56, отрицательных – 39 (χ2 = 3.04); с доминированием G. gymnocaulon – 63 против 50 (χ2 = 1.50), C. arundinacea – 86 против 64 (χ2 = 3.23).
В табл. 4 приведены данные о среднем числе сопутствующих видов растений в отдельных пробах и общем числе таких видов в их группах, сформированных на основе двух признаков: 1) существенно разное участие доминанта (наиболее низкое и наиболее высокое); 2) равная суммарная (для групп проб) биомасса сопутствующих видов растений. Группы проб с низким участием доминантов состояли из 5 шт., с высоким участием – 9−18 шт. Как видно из таблицы, среднее число видов в пробах с низким участием доминантов ожидаемо выше, чем с высоким (в 1.4−2 раза). Однако общее число видов в сравниваемых группах проб отличается в меньшей степени и разнонаправленно. Причем только в группах проб с высоким участием Rubus caesius, Glycyrrhiza glabra, Solidago сanadensis и Geranium gymnocaulon обнаружено значительно меньшее число видов (разница достоверна для P < 0.05), чем в соответствующих им группах проб с низким их участием.
Таблица 4.
Видовое богатство в группах проб с разным участием доминантов, но c примерно равной суммарной биомассой сопутствующих видов
| Доминанты | Ns | Низкое участие доминанта | Высокое участие доминанта | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| n | D | Ws | S | $N_{{\text{s}}}^{'}$ | n | D | Ws | S | $N_{{\text{s}}}^{'}$ | ||
| Alchemilla retinervis | 27 | 5 | 0.39 | 280.5 | 11.4 | 24 | 17 | 0.85 | 280.8 | 6.8 | 21 |
| Kobresia capillifolia | 27 | 5 | 0.42 | 104.4 | 12.8 | 23 | 16 | 0.83 | 104.2 | 6.7 | 21 |
| Geranium gymnocaulon | 42 | 5 | 0.44 | 169.1 | 19.4 | 35 | 10 | 0.77 | 169.1 | 13.2 | 29* |
| Calamagrostis arundinacea | 38 | 5 | 0.25 | 751.2 | 11.6 | 22 | 18 | 0.77 | 751.8 | 8.1 | 28 |
| Inula grandiflora | 25 | 5 | 0.22 | 571.6 | 11.8 | 20 | 13 | 0.74 | 572.8 | 7.8 | 19 |
| Alchemilla persica | 57 | 5 | 0.27 | 340.3 | 28.0 | 49 | 13 | 0.74 | 339.7 | 18.5 | 47 |
| Botriochloa ischaemum | 49 | 5 | 0.37 | 197.4 | 14.4 | 25 | 15 | 0.76 | 198.1 | 10.5 | 43 |
| Calamagrostis epigeios | 30 | 5 | 0.38 | 303.8 | 9.8 | 19 | 13 | 0.79 | 303.2 | 6.2 | 21 |
| Rubus caesius | 27 | 5 | 0.23 | 427.8 | 10.0 | 20 | 10 | 0.78 | 427.2 | 5.5 | 9* |
| Solidago canadensis | 33 | 5 | 0.38 | 317.4 | 12.8 | 27 | 14 | 0.85 | 316.6 | 6.8 | 21* |
| Agropyron pinifolium | 33 | 5 | 0.30 | 230.5 | 11.4 | 24 | 15 | 0.74 | 230.9 | 7.5 | 25 |
| Glycyrrhiza glabra | 17 | 5 | 0.39 | 590.1 | 9.4 | 15 | 9 | 0.72 | 590.9 | 4.7 | 9* |
| Echinochloa crus-galli | 33 | 5 | 0.70 | 30.0 | 8.4 | 18 | 10 | 0.92 | 29.5 | 4.2 | 14 |
| Setaria viridis | 20 | 5 | 0.51 | 48.3 | 6.4 | 14 | 15 | 0.83 | 48.4 | 4.3 | 15 |
| Zostera noltei | 17 | 5 | 0.59 | 16.4 | 5.0 | 9 | 13 | 0.86 | 16.0 | 3.5 | 15 |
Примечание. n – число проб в группах; D – средняя для пробы степень доминирования; Ws – суммарная биомасса сопутствующих видов в группах проб; S – среднее для пробы число видов; Ns – общее число сопутствующих видов в сериях проб; $N_{{\text{s}}}^{'}$ – число сопутствующих видов, обнаруженных в группах проб с разным участием доминанта; * – соотношение $N_{{\text{s}}}^{'}$ и (Ns − $N_{{\text{s}}}^{'}$) в группах проб с высоким участием доминанта ниже, чем в группах проб с низким его участием, причем это различие статистически значимо (критерий χ2, Р < 0.05).
ОБСУЖДЕНИЕ
Итак, для оценки характера исчезновения видов в ответ на усиление позиции доминантов мы изучили 15 участков сообществ, сформированных в местообитаниях разных типов. Результаты анализа данных показали, что хорошо выраженный (выявленный на основе двух подходов) эффект упорядоченного исчезновения видов присутствует только на 3 из 15 участков, а именно с доминированием Rubus caesius, Glycyrrhiza glabra и Solidago сanadensis. Это может означать, что такие доминанты избирательно воздействуют на сопутствующие виды растений, и рост их участия может привести к существенному снижению размера видового пула сообществ. Два из них – S. сanadensis и G. glabra – способны образовывать высокие и густые заросли и, по-видимому, вытеснять в первую очередь виды, более уязвимые к изменению светового режима. По мнениию M. Hejda et al. [2], именно такие доминанты оказывают наиболее сильное воздействие на другие виды травяных ценозов. Скорее всего, такой же механизм (дифференцированная реакция видов на изменение освещенности) является причиной низкого видового богатства сообществ с сомкнутым пологом R. caesius.
Однако во всех остальных сообществах (в том числе с доминированием видов, известных как мощные конкуренты: Botriochloa ischaemum и Calamagrostis epigeios [3, 6]) сопутствующие виды, по-видимому, слабо отличаются по устойчивости к воздействию рассматриваемого фактора. В результате низкое видовое богатство ценозов с высокой степенью их доминирования является результатом преимущественно случайного, а не упорядоченного исчезновения видов. В этом случае сообщества с разным участием доминантов должны характеризоваться примерно одинаковыми по размеру видовыми пулами. Соответственно на небольших участках сообществ с низкой степенью доминирования потенциально может произрастать столько же видов, как и на более крупных участках с высокой степенью доминирования. Таким образом, наши результаты позволяют предположить, что во многих, возможно в большинстве растительных сообществ разных моделей организации, рост относительного участия доминантов может угрожать многим другим видам только в том случае, если площадь этих сообществ невелика.
Полученные нами данные согласуются с результатами оценки регионального воздействия доминантов на видовое богатство растительных сообществ, представленными в ряде других публикаций. В частности, ранее мы сопоставили [24] видовое богатство и размер видового пула древостоев Западного Кавказа с низкой и высокой степенью доминирования. Результаты показали, что первые характеризуются более высоким видовым богатством, чем вторые, при этом размер их видового пула за некоторым исключением является сходным. Аналогичные выводы были сделаны в ряде обзоров, посвященных воздействию на видовое богатство растительных сообществ чужеродных доминантов [11, 25]. Мета-анализу были подвергнуты данные полевых исследований и экспериментов, выполненных на площадках разного размера. Обнаружено, что такие виды обычно снижают видовое богатство сообществ, но сила этого эффекта ослабевает по мере увеличения пространственного масштаба исследования (размера используемой площадки). Было сделано предположение [11], что это следствие либо неизбирательного воздействия данного фактора на разные виды, либо большей устойчивости к такому воздействию редких видов по сравнению с обычными.
Дополнительно K.T. Powell et al. [12] сопоставили участки лесных биомов разных типов (штаты Гавайи, Миссури и Флорида, США) с доминированием чужеродных видов растений (проективное покрытие >90%) со сходными по условиям произрастания участками лесов, но с доминированием аборигенных растений, участие которых было ниже. Воздействие чужеродных и аборигенных видов на биоразнообразие в разном пространственном масштабе оценивали путем сопоставления кривых регрессии “площадь−число видов”. Результаты показали, что сообщества с доминированием чужеродных видов характеризуются относительно низким видовым богатством на небольших участках, но более высокой скоростью накопления видов при увеличении их площади по сравнению с сообществами с доминированием аборигенных видов. В итоге на крупных участках сообществ с доминированием чужеродных и аборигенных видов видовое богатство оказывается близким.
По мнению K.T. Powell et al., выявленная ими закономерность имеет универсальный характер, с чем, однако, не согласны авторы работ [10, 26]. На основе анализа двух групп данных по лесным, луговым и пустынным ценозам территории Северной Америки (США) они показали [26], что имеет место широкий континуум в соотношении реакции растительных сообществ на воздействие чужеродных доминантов в разном пространственном масштабе. Об этом свидетельствуют и некоторые другие результаты. Например, данные по растительным сообществам Чехии [2] показывают, что внедрение в сообщества чужеродных доминантов, достигающих более высокого покрытия, чем аборигенные, ведет к большему или меньшему (в зависимости от вида растения), но примерно пропорциональному снижению их видового богатства на участках разной площади. По мнению D.P. Rijal et al. [27], в растительных сообществах северной Скандинавии с доминированием Heracleum persicum (родина – Иран) способны произрастать лишь несколько высокорослых местных видов, что, по-видимому, является результатом избирательного вытеснения им других видов.
Неоднозначность как опубликованных ранее, так и наших результатов может быть вызвана особенностями воздействия тех или иных доминантов на сопутствующие виды растений [9, 28]. В одних случаях это выражается в сокращении ими пространства и ресурсов на микроучастках, в других – в трансформации ими местообитаний путем накопления слоя опада, изменения светового режима, физико-химических свойств почвы и т.д. Доминанты, способные существенно изменять условия произрастания для других видов, называют эдификаторами [29], в инвазионной экологии – “видами-трансформерами” [9, 28, 30]. Сделано предположение [9], что среди чужеродных видов растений “трансформеры” составляют около 10%. В частности, к ним относится изученный нами Solidago сanadensis [30]. Однако способность других таких видов избирательно воздействовать на сопутствующие виды растений требует проверки. Например, она не была выявлена у Acer negundo – древесного вида североамериканского происхождения, который относят к этой категории (исследование было проведено в урбанизированных лесах г. Екатеринбурга) [16].
Кроме того, необходимо учитывать, что результаты воздействия доминирующих видов на крупные участки растительных сообществ могут определяться не только особенностями их биологии (эдификаторными возможностями), но и условиями среды либо особенностью самих сообществ [31–33]. Этот аспект проблемы более актуален для видов, способных доминировать в местообитаниях разных типов, таких, например, как Botriochloa ischaemum, Solidago сanadensis или Rubus caesius. Известно также, что местные виды растений более устойчивы к воздействию аборигенных доминантов, чем чужеродных, поскольку имеют длительный период совместного существования [32–34]. В пользу этого представления свидетельствуют и наши результаты, поскольку одним из трех видов, предположительно способных избирательно влиять на другие виды, оказался Solidago сanadensis – пришелец из Северной Америки. Однако в целом знания об этом также очень ограниченны.
Исследования выполнены при финансовой поддержке Российского фонда фундаментальных исследований (гранты № 16-04-00228 и 20-04-00364). Мы благодарны профессору М. Рейманеку (M. Rejmánek) за рекомендацию обратить внимание на роль доминантов в ограничении видового богатства на крупных участках растительного покрова и за помощь в поиске публикаций по этой проблеме.
Авторы признательны неизвестным рецензентам за тщательный анализ статьи и рекомендации по ее улучшению.
Список литературы
Hillebrand H., Bennett D.M., Cadotte M.W. Consequences of dominance: a review of evenness effects on local and regional ecosystem processes // Ecology. 2008. V. 89. № 6. P. 1510–1520.
Hejda M., Pyšek P., Jarošik V. Impact of invasive plants on the species richness, diversity, and composition of invaded communities // J. Ecol. 2009. V. 97. P. 3393– 3403.
Somodi I., Virágh K., Podani J. The effect of the expansion of the clonal grass Calamagrostis epigejos on the species turnover of a semi-arid grassland // Applied Vegetation Science. 2008. V. 11. №. 2. P. 187–194.
Миркин Б.М., Наумова Л.Г. Современное состояние основных концепций науки о растительности. Уфа: АН РБ, Гилем, 2012. 488 с.
Csergő A.M., Demeter L., Turkington R. Declining diversity in abandoned grasslands of the Carpathian Mountains: do dominant species matter? // PLoS ONE. 2013. V. 8. №. 8: e73533.oi:10.1371.
Bartha S., Szentes Sz., Horváth A. et al. Impact of mid-successional dominant species on the diversity and progress of succession in regenerating temperate grasslands // Applied Vegetation Science. 2014. V. 17. № 2. P. 201–213.
Elumeeva T.G., Onipchenko V.G., Weger M.J.A. No other species can replace them: evidence for the key role of dominants in an alpine Festuca varia grassland // J. of Vegetation Science. 2017. V. 28. P. 674–683.
Акатов В.В., Акатова Т.В., Чефранов С.Г. Связь доминирования и выравненности с продуктивностью и видовым богатством в растительных сообществах разных моделей организации // Экология. 2018. № 4. С. 264–274. [Akatov V.V., Akatova T.V., Chefranov S.G. The relationship of dominance and evenness with productivity and species richness in plant communities with different organization models // Russ. J. Ecol. 2018. V. 49. № 4. Р. 296–305.]
Rejmánek M., Richardson D.M., Pyšek P. Plant invasions and invasibility of plant communities // Vegetation Ecology, Second Edition / Eds. van der Maarel E., Chichester J.F. United Kingdom: Wiley & Sons, Ltd., 2013. P. 387‒424.
Rejmánek M., Stohlgren T.J. Scale-dependent impacts of invasive species: a reply to Chase et al. (2015) // Biol. Lett. 2015. V. 11: 20150402. https://doi.org/10.1098/rsbl.2015.0402
Powell K.I., Chase J.M., Knight T.M. A synthesis of plant invasion effects on biodiversity across spatial scales // Amer. J. Bot. 2011. V. 98. № 3. P. 539−548.
Powell K.I., Chase J.M., Knight T.M. Invasive plants have scale-dependent effects on diversity by altering species-area relationships // SCIENCE. 2013. V. 339. P. 316–318.
Акатов В.В., Акатова Т.В., Чефранов С.Г. Степень доминирования и видовое богатство в растительных сообществах с высокой и низкой интенсивностью межвидовой конкуренции // Журн. общ. биол. 2017. Т. 78. № 4. С. 52–64. [Akatov V.V., Akatova T.V., Chefranov S.G. Degree of dominance and species richness in plant communities with high and low intensity of interspecies competition // Biol. Bull. Rev. 2018. V. 8. № 5. Р. 389–400.]
Solan M., Cardinale B.J., Downing A.L. et al. Extinction and ecosystem function in the marine benthos // Science. 2004. V. 306. P. 1177–1180.
Gross K., Cardinale B.J. The functional consequences of random vs. ordered species extinctions // Ecology Letters. 2005. V. 8. P. 409–418.
Веселкин Д.В., Дубровин Д.И. Разнообразие травяного яруса урбанизированных сообществ с доминированием инвазивного Acer negundo // Экология. 2019. № 5. С. 323–331. [Veselkin D.V., Dubrovin D.I. Diversity of the grass layer of urbanized communities dominated by invasive Acer negundo // Russ. J. Ecol. 2019. V. 50. № 5. Р. 413–421.]
Onipchenko V.G., Semenova G.V., van der Maarel E. Population strategies in severe environments: alpine plants in the northwestern Caucasus // J. Veg. Sci. 1998. V. 9. P. 27–40.
Миркин Б.М., Ямалов С.М., Наумова Л.Г. Синантропные растительные сообщества: модели организации и особенности классификации // Журн. общ. биол. 2007. Т. 68. № 6. С. 435–443.
Cabi E., Karabacak E., Çingay B. Agropyron pinifolium Nevski (Poaceae): a new species record for the flora of Turkey // Biological Diversity and Conservation. 2015. V. 8. № 1. P. 90–93.
Weber E. Biological flora of Central Europe: Solidago altissima L. // Flora. Switzerland. 2000. V. 195. P. 123–134.
Морозова О.В., Виноградова Ю.К. Solidago canadensis – Золотарник канадский // Самые опасные виды России (Топ-100) / Под ред. Дгебуадзе Ю.Ю., Петросяна В.Г., Хляп Л.А. М.: Тов-во научн. изд. КМК, 2018. 688 с.
Wright D.H. Species-energy theory: an extension of species-area theory // Oikos. 1983. V. 41. P. 496–506.
Srivastava D.S., Lawton J.H. Why more productive sites have more species: an experimental test of theory using tree-hole communities // Amer. Nat. 1998. V. 152. P. 510–529.
Акатов В.В. Состав, видовое богатство и размер видового пула моно- и олигодоминантных древостоев Западного Кавказа // Растительность России. 2018. № 32. С. 3–18.
Gaertner M., Breeyen A.D., Hui C., Richardson D.M. Impacts of alien plant invasions on species richness in Mediterraneantype ecosystems: A meta-analysis // Progress in Physical Geography. 2009. V. 33. P. 319–338.
Stohlgren T.J., Rejmánek M. No universal scale-dependent impacts of invasive species on native plant species richness // Biol. Lett. 2014. V. 10: 20130939. https://doi.org/10.1098/rsbl.2013.0939
Rijal D.P., Alm T., Inger L.N., Alsos G. Giant invasive Heracleum persicum: Friend or foe of plant diversity? // Ecology and Evolution. 2017. V. 7. P. 4936–4950.
Richardson D.M., Pyšek P., Rejmánek M. et al. Naturalization and invasion of alien plants: Concepts and definitions // Diversity and Distributions. 2000. V. 6. P. 93–107.
Работнов Т.А. Фитоценология. М.: Изд-во МГУ, 1983. 296 с.
Виноградова Ю.К., Майоров С.Р., Хорун Л.В. Черная книга флоры Средней России (Чужеродные виды растений в экосистемах Средней России). М.: ГЕОС, 2009. 494 с.
Vítková M., Müllerová J., Sádlo J. et al. Black locust (Robinia pseudoacacia) beloved and despised: A story of an invasive tree in Central Europe // Forest Ecology and Management. 2017. V. 384. P. 287–302.
Hejda M., Štajerová K., Pyšek P. Dominance has a biogeographical component: do plants tend to exert stronger impacts in their invaded rather than native range? // J. of Biogeography. 2017. V. 44. P. 18–27.
Meiners S.J., Pickett S.T.A., Cadenasso M.L. Effects of plant invasions on the species richness of abandoned agricultural land // Ecography. 2001. V. 24. P. 633–644.
Rejmánek M., Simberloff D. Origin matters // Environmental Conservation. 2017. V. 44. № 2. P. 97–99.
Галушко А.И. Флора Северного Кавказа. Ростов-на-Дону: Изд-во Ростовского ун-та, 1978. Т. 1. 320 с.; 1980. Т. 2. 352 с.; 1980. Т. 3. 328 с.
Guiry M.D., Guiry G.M. AlgaeBase, Worldwide electronic publication, National University of Ireland, Galway, 2020. http://www.algaebase.org
Дополнительные материалы отсутствуют.