

Экология, 2021, № 6, стр. 439-449
Сезонное распределение и перемещения морского зайца, Erignathus barbatus nauticus, в Охотском море по данным спутниковой телеметрии
М. А. Соловьёва a, *, Д. М. Кузнецова a, Д. М. Глазов a, В. В. Рожнов a
a Институт проблем экологии и эволюции им. А.Н. Северцова РАН
119071 Москва, Ленинский просп., 33, Россия
* E-mail: solovjova.m@gmail.com
Поступила в редакцию 08.10.2020
После доработки 08.10.2020
Принята к публикации 01.02.2021
Аннотация
Распределение и образ жизни морского зайца, или лахтака (Erignathus barbatus), на большей части видового ареала тесно связаны с ледовой кромкой, положение которой меняется в течение года. Охотское море – сезонно замерзающая акватория, которая летом полностью освобождается ото льда, что определяет годовой цикл обитающего в нем тихоокеанского подвида лахтака (E. barbatus nauticus) и делает его уникальным по сравнению с животными из других частей ареала. С помощью спутниковой телеметрии выявлены основные сезонные местообитания и характер перемещений лахтаков в Охотском море. Установлено, что лахтаки использовали различные районы в безледный и ледовый периоды, совершая протяженную миграцию между ними. В безледный период охотоморские лахтаки используют наиболее мелководную часть Сахалинского залива. В ледовый период животные широко расходятся по акватории, занимая различные ее участки от западной части Сахалинского залива до восточного побережья о-ва Сахалин. На даты и направление сезонных перемещений лахтака наибольшее влияние оказывают сроки и характер формирования ледового покрова. Выбор лахтаком сезонных местообитаний определяется наименьшими доступными глубинами в течение всего года, а в зимне-весенний период – и ледовым покровом.
Морской заяц, или лахтак (Erignathus barbatus, Erxleben, 1777), распространен циркумполярно [1], в том числе по всей Российской Арктике. Он является облигатным пагофилом и тесно связан со льдом в периоды отдыха, размножения, лактации, спариваний и линьки [2–5]. Строго привязанный ко льду в зимний период [6], лахтак особенно чувствителен к изменениям количества, распределения и структуры льда, которые происходят в последние десятилетия [7, 8]. Особенности биологии делают морского зайца уязвимым по отношению к возрастающей антропогенной нагрузке. Хотя в рационе лахтака присутствуют различные объекты, предпочитаемой пищей, особенно в летнее время, являются бентосные виды беспозвоночных и рыб [9], что вынуждает его держаться прибрежных мелководных, наиболее подверженных антропогенному воздействию участков акватории [10]. Именно в таких местах ведутся разведка и добыча углеводородов, постоянно повышены судовой трафик и уровень антропогенного шума, что также может негативно повлиять на лахтака, поскольку он чувствителен к подводным звукам [11].
Особенности биологии лахтака изучены в разной степени. Много данных имеется о распределении вида в летний период [12–14], сроках и особенностях его размножения [15, 16], спектре питания [17, 18]. В Карском море занимающий вершину трофической цепи лахтак может быть видом-индикатором состояния морских экосистем [19]. В то же время сведения о сезонных перемещениях лахтака и влиянии на них отдельных факторов крайне скудны для Российской Арктики, хотя именно этим аспектам биологии вида в современной мировой литературе уделяется наибольшее внимание [1, 20].
Обитающий в Охотском море тихоокеанский подвид лахтака (E. barbatus nauticus) также изучен неравномерно. Основные исследования были проведены в северной зоне Охотского моря [16, 21, 22], и лишь немногие касались лахтака в его южной части [17, 23]. В отличие от других морей, входящих в ареал лахтака, Охотское море является закрытым и сезонно замерзающим. При таком ледовом режиме реализация характерной для лахтака из других популяций стратегии перемещаться вслед за ледовой кромкой [3, 24–26] оказывается невозможной. Вопросы миграций и выбора сезонных местообитаний в Охотском море, связи перемещений животных с параметрами окружающей среды остаются наиболее актуальными в изучении экологии этого вида.
Целью нашей работы было изучение распределения и перемещений лахтака в Охотском море с использованием спутниковой телеметрии в период с начала осени до конца весны.
МАТЕРИАЛ И МЕТОДЫ
Отлов и мечение. Животные были отловлены и помечены в окрестностях о-ва Чкалова (Охотское море, Сахалинский залив) в сентябре 2013 г. Отлов лахтаков проводили на залёжках с помощью сетей. Медицинские препараты для обездвиживания не применяли. У всех отловленных животных определяли пол и измеряли зоологическую длину, на основании которой с использованием таблиц Э.А. Тихомирова [27] определяли их половозрелость (возраст) (табл. 1).
Таблица 1.
Информация о помеченных животных
| № метки | Тип метки | Пол | Длина тела, см | Возраст | Дата установки метки | Дата получения последнего сигнала | Число дней работы метки | Число локаций после фильтрации |
|---|---|---|---|---|---|---|---|---|
| 39 497* | MK-10 | ♂ | 192 | subad | 16.09.2013 | 14.03.2014 | 179 | 3351 |
| 99 278* | MK-10 | ♂ | 175 | subad | 16.09.2013 | 02.04.2014 | 198 | 3514 |
| 66 976* | MK-10 | ♀ | 190 | ad | 16.09.2013 | 05.04.2014 | 201 | 3361 |
| 66 979* | MK-10 | ♀ | 210 | ad | 16.09.2013 | 28.04.2014 | 224 | 4207 |
| 99 276* | MK-10 | ♀ | 211 | ad | 16.09.2013 | 28.04.2014 | 224 | 4187 |
| 66 981 | SPOT-5 | ♀ | 180 | ad | 16.09.2013 | – | 0 | – |
| 66 984 | SPOT-5 | ♀ | 190 | ad | 16.09.2013 | – | 0 | – |
| 99 290 | SPOT-5 | ♀ | 208 | ad | 16.09.2013 | 14.11.2013 | 59 | 2 |
| 99 294 | SPOT-5 | ♀ | 190 | ad | 16.09.2013 | 04.12.2013 | 79 | 33 |
| 99 296/110 717 | SPOT-5/Пульсар | ♂ | 197 | ad | 09.09.2013 | 20.10.2013/04.11.2013 | 41/56 | –/265 |
Всего нами были помечены 10 лахтаков, на которых было установлено 11 передатчиков (меток), работающих в спутниковой системе Argos. Использовали три типа передатчиков: MK-10 (пять) и SPOT-5 (пять) производства Wildlife Computers Inc. (США), а также один “Пульсар” (ЗАО “Эс-Пас”, Россия), установленный дополнительно на лахтака с передатчиком SPOT-5.
Режим передачи данных для передатчиков MK-10 был запрограммирован 6/6 ч, для передатчика Пульсар – 6/6 ч или 6/18 ч (6 ч данные передаются на спутник, следующие 6 или 18 ч метки находятся в режиме ожидания). Ластовые метки SPOT-5 передавали данные в течение одних суток (круглосуточно) через каждые 6 сут. Передатчики MK-10 и Пульсар приклеивали на голову или между лопаток животного с помощью быстросохнущей эпоксидной смолы (шкуру тюленей предварительно очищали и обезжиривали), метку SPOT-5 крепили на межпальцевую перепонку задней ласты. Продолжительность работы приклеенных передатчиков определяется линькой, которая у лахтака на юге Охотского моря нередко начинается уже в апреле [17, 28]. Из 11 установленных передатчиков достаточно долго для получения необходимого для обработки количества данных проработало пять – срок их работы составил от 179 до 224 дней (см. табл. 1).
Обработка данных. Получаемые с передатчиков данные проходили предварительную фильтрацию с помощью фильтра Kalman Filtering algorithm на сайте системы Argos [29]. Дальнейшую их фильтрацию производили SDA-фильтром пакета argosfilter для R [30]. Параметры фильтрации: максимальная скорость перемещения тюленей – 3.8 м/с [31]; максимальный угол между двумя отрезками пути длиной больше 2.5 км – 15°, 5 км – 25° [32]. Затем в пакете программ ArcGIS вручную удаляли все точки, ошибочно попавшие на сушу и удаленные от береговой линии в глубь материка более чем на 1 км. После применения всех вышеописанных фильтров для последующего анализа мы получили от пяти передатчиков 18 620 локаций. Среднее количество локаций в день составило 18.02 (SD = 0.84).
Площадь участков обитания помеченных лахтаков определяли методом минимального выпуклого полигона (Minimum Convex Polygon, далее MCP100%) [33]. Для выявления ключевых для лахтака участков акватории применяли метод фиксированного контура (метод ядерных полигонов, kernel method) [34]. Как правило, используются полигоны 50 и 95% (kernel50% и kernel95%). Расчеты проводили с помощью пакетов Home range и Animal movements для программы ArcView GIS 3.2 [35].
Для анализа ледовой обстановки использовали ежедневные карты ледового покрытия c ресурса National Ice Center, показывающие границы сплошного и разреженного льда (http://www.natice.noaa.gov), а также карты Государственного научного центра “Арктический и антарктический научно-исследовательский институт” (http://www.aari.ru), поставляемые 1–2 раза в неделю в зависимости от сезона года и дающие представление о типе, толщине, сплоченности льда и других его характеристиках.
При анализе батиметрии применяли карты GebcoMaps для Охотского моря (http://www.gebco.net) с вертикальной точностью до 1 м и пространственным разрешением 00°00′30″. Лахтаки питаются преимущественно бентосными организмами [9], погружаясь на всю толщу воды. Поэтому, хотя передатчики, установленные на лахтака, не были снабжены датчиком глубины погружения, мы считали глубинами погружения животных глубину используемой ими акватории.
Дальнейшую обработку данных и картографический анализ материала проводили в пакете программ ArcGIS 9.3.1, а статистический анализ – в программе Statistica.
РЕЗУЛЬТАТЫ
Перемещения лахтаков и использование акватории. Описания перемещений пяти помеченных лахтаков, данные от которых вошли в анализ использования животными акватории Охотского моря, и их треки приведены на рис. 1.
Рис. 1.
Треки пяти лахтаков, помеченных передатчиками, за период их работы. Место мечения (о-в Чкалова) отмечено звездочкой (*).

Самец № 39 497. До начала ноября не покидал район мечения, 80% времени провел у берегов островов Чкалова и Байдукова с мористой части, не удаляясь от них более чем на 15 км, остальное время – в заливе Счастья. После 11.11.2013 г. начал регистрироваться мористее, к северу от места поимки. Декабрь, январь и февраль провел в западной части Сахалинского залива. В марте двинулся обратно, на юго-восток. Передатчик прекратил работу 14.03.2014 г.
Самец № 99278. До середины ноября не удалялся от места мечения более чем на 46 км. Перемещался главным образом в заливе Счастья и между островами Чкалова и Байдукова. Со второй половины ноября до конца декабря регистрировали к северу-востоку от места поимки, не далее 100 км от него. Начал движение на северо-восток 28.12.2013 г., обогнул о-в Сахалин и двинулся вдоль его восточного побережья на юг, преодолев за 10 дней более 850 км. К 19.01.2014 г. спустился еще на 120 км южнее и до прекращения работы передатчика (2.04.2014) перемещался в районе Набильского залива на расстоянии 30–130 км от побережья.
Самка № 99 276. До начала ноября практически не покидала район мечения, удаляясь от места поимки не более чем на 40 км. Большую часть времени провела между островами Чкалова и Байдукова и в заливе Счастья. Один раз переместилась до побережья о-ва Сахалин и обратно. Со второй декады ноября ее регистрировали в более мористой части к северо-востоку от места поимки, во второй половине декабря ушла на северо-восток от о-ва Чкалова. Пройдя около 80 км вдоль побережья, заняла район, в котором регистрировали до начала апреля. Вновь сменила район пребывания 6.04.2014 г. и переместилась немного северо-восточнее, где находилась до 23.04.2014 г. Передатчик прекратил работу 28.04.2014 г.
Самка № 66 976. До 6.11.2013 г. находилась в районе мечения, не удаляясь от места поимки далее 30 км. В течение 6–17.11.2013 г. переместилась на 100 км на запад вдоль побережья и вернулась в район мечения, оставаясь в юго-западной части Сахалинского залива и не удаляясь от побережья далее 50 км. Начала направленное движение на северо-восток 27.12.2013 г., к 1.01.2014 г. достигла северной оконечности о-ва Сахалин, обогнула его и продолжила движение вдоль его восточного побережья на юг, пройдя за 19 дней более 950 км. С 15.01.2014 г. по 29.03.2014 г. перемещалась вдоль восточного побережья о-ва Сахалин на расстоянии 100 км, после чего поднялась на 130 км к северу. Отмечена южнее Набильского залива 1.04.2014 г., где находилась до прекращения работы передатчика 5.04.2014 г.
Самка № 66 979. До 10.11.2013 г. находилась в районе мечения, практически не перемещаясь. По южной части Сахалинского залива перемещалась до 27.12.2013 г. на расстоянии не более 40 км от побережья. Затем прошла через его центральную часть и двинулась к северной оконечности о-ва Сахалин, достигнув ее 11.01.2014 г. В этом районе находилась до 19.04.2014 г., после чего двинулась на юг в обратном направлении. Передатчик прекратил работу 28.04.2014 г.
Расстояние, которое лахтаки преодолевали за день, за весь период наблюдений составило в среднем 95.6 км (SD = 26.7). Общее расстояние, пройденное животными за время работы передатчиков, составило 19 689 км (SD = 5403.8), самый длинный суммарный путь (25 142 км) принадлежит самке с передатчиком № 66 979.
Все время наблюдений ‒ с 16.09.2013 г. до прекращения работы передатчиков в марте – апреле 2014 г., можно разделить на четыре периода, которые отличаются особенностями распределения помеченных нами лахтаков по акватории Охотского моря, характером их перемещений и расстоянием от места поимки и мечения: 1 – период нахождения лахтаков в акватории Сахалинского залива в районе их поимки и мечения (залив Счастья, район островов Чкалова и Байдукова); 2 – период расширения используемой акватории до всей юго-западной части Сахалинского залива; 3 – период активного перехода из юго-западной части Сахалинского залива; 4 – период нахождения в акватории, которой лахтаки достигли в результате активного перехода. Использование акватории лахтаками в эти периоды показано на рис. 2.
Рис. 2.
Общая акватория (MCP100%) и ее ключевые участки, выявленные методом kernel (kd50% и kd95%), для помеченных лахтаков в разные периоды: а – с 16.09.2013 по 10.11.2013; б – с 11.11.2013 по 27.12.2013; в – с 27.12.2013 по 15.01.2014 (показаны треки перемещений); г – с 16.01.2014 до конца прослеживания; * ‒ место мечения (о-в Чкалова).
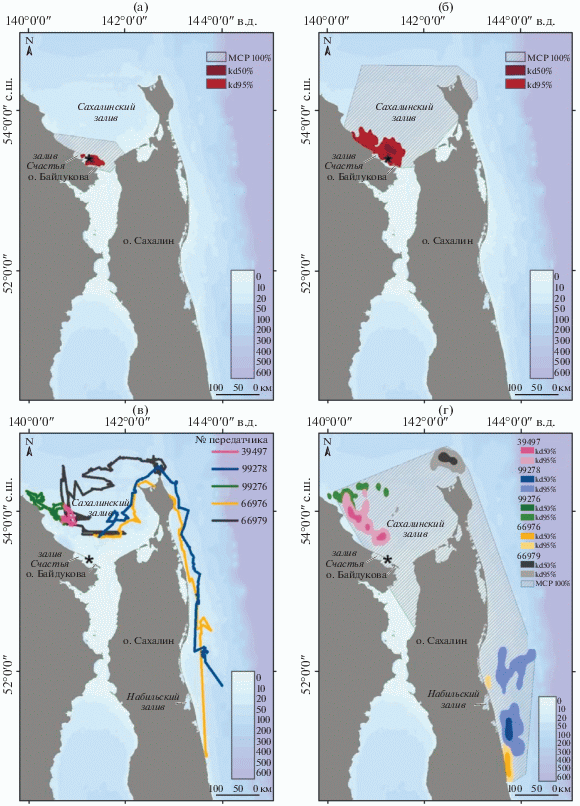
С момента установки передатчиков 16.09.2013 г. и до 11.11.2013 г. все помеченные животные практически не покидали район мечения и перемещались не далее 40 км от места отлова (M = 13.1 км; SD = 13.2, n = 6541 локация). В течение этого времени лахтаки активно использовали акваторию залива Счастья и прилегающую к островам Чкалова и Байдукова часть Сахалинского залива. Далее 40 км от места отлова животные удалились в этот период лишь дважды: самка № 99276 совершила радиальный переход до о-ва Сахалин, а самка № 66 976 – вдоль побережья в западную часть Сахалинского залива. Площадь используемой пятью помеченными лахтаками акватории в этот период составила: общая – 7649.3 км2 (MCP100%), ключевая ее часть – 847.2 км2 (kernel95%) (см. рис. 2а).
После 11.11.2013 г. все животные постепенно стали использовать всю юго-западную часть Сахалинского залива. Лахтаки стали удаляться дальше от места мечения (M = 38.6 км; SD = 27.5), возросла и площадь используемой ими акватории: как общая – 14 864.0 км2 (MCP100%), так и ключевая ее часть – 5052.8 км2 (kernel95%) (см. рис. 2б).
С середины и до конца декабря все лахтаки начали совершать относительно большие переходы (от 100 до 550 км), которые занимали у них от нескольких дней до трех недель, в другие акватории. Используемая ими за этот период перехода акватория показана на рис. 2в. К 15.01.2014 г. лахтаки распределились по трем районам, в которых затем пребывали до прекращения работы передатчиков: в западной части Сахалинского залива (№ 99 276 и 39 497), около северной оконечности о‑ва Сахалин (№ 66 979) и у его восточного побережья в районе Набильского залива (№ 66 976 и 99 278) (рис. 2г и 3). В этот период удаленность от места мечения у всех лахтаков была максимальной (M = = 222.3 км; SD = 115.0).
Ледовая обстановка. Одним из основных факторов, определяющих перемещения лахтаков, является ледовая обстановка. В период наблюдений в Сахалинском заливе припайный лед начал формироваться 10.11.2013 г. и в течение нескольких дней он полностью покрыл залив Счастья. Основная часть Сахалинского залива в течение ноября и декабря продолжала оставаться свободной ото льда: появляющийся нилас (тонкая ледовая корка толщиной до 10 см) держался несколько дней и затем таял. После 20.12.2013 г. весь Сахалинский залив начал закрываться сначала ниласом, затем молодым льдом (толщиной 10–30 см), а в начале января 2014 г. – толстым однолетним льдом (толщиной 30–200 см). По мере формирования льда и изменений его типа и сплоченности происходили изменения в местах регистрации лахтаков (рис. 3). В ноябре помеченные животные встречались и среди ниласа, и однолетнего льда, и припая, но еще довольно часто (65.2% всех локаций) находились среди открытой воды. В декабре доля локаций лахтаков среди открытой воды уменьшилась (24.3%), а среди молодого льда возросла (73.3%). В январе животных начали регистрировать среди толстого однолетнего льда (на него приходится почти половина всех локаций – 48.1%), и с января по апрель доля их локаций здесь росла, достигнув в марте–апреле более 80%. В апреле лахтаки вновь появились среди открытой воды. В районах с припайным льдом в течение всего времени прослеживания животных практически не регистрировали (см. рис. 3а).
Рис. 3.
Доля локаций лахтаков, зарегистрированных на различных типах льда и среди льда различной сплоченности: а – тип льда: 1 – чисто, 2 – нилас, 3 – молодой, 4 – однолетний, 5 – припай; б – сплоченность льда: 1 – 40–60%, 2 – 70–80%, 3 – 90–100%.

С начала формирования льда и до окончания прослеживания лахтаков сплоченность льда в местах их регистрации всегда была более 40% и по мере развития ледовой обстановки постепенно увеличивалась. В ноябре доля локаций среди льда сплоченностью 70–80% составила 83.6%, а при сплоченности 90–100% – 16.4%; в декабре при сплоченности льда 90–100% лахтаки стали регистрироваться чаще, а затем практически всегда находились на таком льду (см. рис. 3б).
Глубина акватории в районах перемещений лахтаков. Глубина акватории, на которой регистрировали лахтаков (рис. 4), в течение периода наблюдений изменялась от 0 до 160 м (однако за все время наблюдений в акваториях с глубинами более 100 м были зарегистрированы только 3.6% локаций животных и только у одного животного).
Рис. 4.
Средняя глубина акватории в местах регистрации лахтаков в течение суток. Вертикальной линией обозначена дата начала образования припая (11.11.2013).
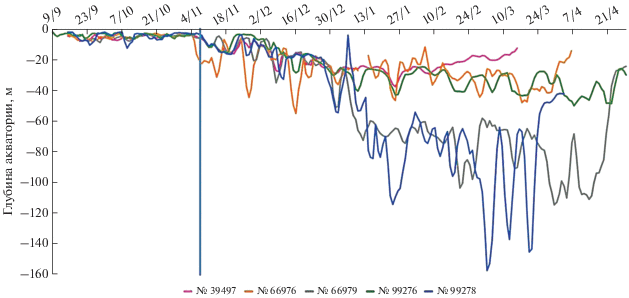
С момента мечения 16.09.2013 г. до 10.11.2013 г. животные находились в очень мелководных районах: 94% их локаций отмечены на акватории с глубинами менее 10 м. С 11.11.2013 г., когда начал образовываться припай, до начала января глубины акватории, на которых регистрировали лахтаков, несколько увеличились, но не превышали 40 м. В начале января часть лахтаков (№ 39 497, 66 976, 99 276) осталась в относительно неглубоководных районах и практически не регистрировалась в водах с глубинами более 40 м, а другая часть (№ 66 979 и 99 278) перешла в более глубоководные районы – от 60 до 180 м. Расстояние от места мечения и средние глубины, на которых регистрировали лахтаков в разные периоды, представлены на рис. 5.
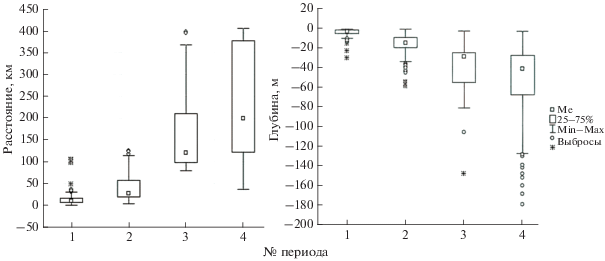
Рис. 5.
Расстояние от места мечения и глубины, на которых регистрировали лахтаков в разные периоды (Me – медиана, Min – минимальное значение, Max – максимальное значение): 1 – нахождение лахтаков в акватории Сахалинского залива в районе их поимки и мечения (залив Счастья, район островов Чкалова и Байдукова); 2 – расширение используемой акватории до всей юго-западной части Сахалинского залива; 3 – активный переход из юго-западной части Сахалинского залива; 4 – нахождение в акватории, которой лахтаки достигли в результате активного перехода.
ОБСУЖДЕНИЕ
Выделенные периоды (нахождение лахтаков в акватории Сахалинского залива в районе их поимки и мечения, расширение используемой акватории до всей юго-западной части Сахалинского залива, активный переход из юго-западной части Сахалинского залива и нахождение в акватории, которой лахтаки достигли в результате активного перехода) сопряжены с особенностями абиотических условий – состоянием акватории и характером ледового покрова, глубинами акватории в местах регистрации лахтаков. Основная причина, из-за которой лахтаки меняют свое местоположение, – изменение ледовой обстановки. Эта причина указывается как главная для перемещений лахтаков и в других частях ареала [36, 37]. Кроме того, отмечено изменение спектра питания лахтаков в течение года. Моллюски являются важнейшей частью рациона весной и летом, так как в это время года могут быть недоступны другие компоненты рациона [4, 9]. К осени моллюски становятся менее активными, лед может полностью закрыть для лахтаков доступ к этому ресурсу, в пище начинают доминировать креветки и крабы, а зимой в некоторых регионах лахтаки могут полностью переходить на питание рыбой [38]. Питание лахтаков в осенне-зимние месяцы в Охотском море не исследовалось, поэтому мы не можем оценить, изменяется ли рацион в выделенные нами периоды и что является первоочередным – смена рациона или смена местообитания. В приведенных выше работах влияние смены питания на перемещения лахтаков не отмечалось.
Первый период характеризуется отсутствием льда. Он заканчивается началом формирования прибрежного льда, который затрудняет нахождение животных в мелководном районе. Глубины, которых придерживаются в это время лахтаки, крайне малы (<10 м). На большей части ареала лахтак не делает залежек на берегу, всегда оставаясь привязанным к ледовой кромке [39], но там, где летом акватория полностью освобождается ото льда, он может образовывать скопления на низких берегах или банках. Такие залёжки известны для Охотского моря [13, 22], хотя привязанность лахтаков к ним не подтверждена. Из 8 животных, передатчики которых доработали до начала ноября, ни одно из них не ушло из района мечения. Этот период – время нагула лахтака, и такая привязанность его к Сахалинскому заливу объясняется высокой продуктивностью бентософауны данного района [40].
Второй период характеризуется формированием ледового покрова, который в 2013 г. в Сахалинском заливе начал появляться 10 ноября. Сразу после появления лед еще не способен выдержать вес взрослого лахтака, однако он преграждает тюленю выход на береговые залёжки и ограничивает добычу корма около побережья, что приводит к вытеснению лахтаков. Удаленность животных от места мечения возрастает – они находятся мористее и в более западной части Сахалинского залива, еще свободной ото льда. Незначительно возрастает и глубина акваторий, в которых находятся животные. В рационе лахтака летом и осенью доминируют бентосные организмы [41, 42]. Поэтому даже после формирования ледового покрова животные продолжали находиться в водах с наименьшей глубиной, которые еще не закрыты льдом. В декабре доля локаций лахтаков среди льда достигает 73.3%. По имеющимся данным мы не можем точно определить, залегал ли тюлень на льду или находился в воде среди льдин. Однако уже в декабре молодой лeд может достигать толщины 30 см [43], и лахтаки могут использовать его для залегания. С конца декабря, с развитием более плотного и толстого льда, который начал закрывать Сахалинский залив, начинается третий период, выделенный нами в перемещениях лахтаков.
Третий период – самый непродолжительный по времени (менее 3 недель) и проходит при установившемся ледовом покрове. Лахтаки в этот период направленно и быстро перемещаются из Сахалинского залива в другие районы, в которых они проводят зиму. Основная часть перемещений проходит вдоль берега, в акваториях с небольшими глубинами (<60 м) (пути перемещений двух помеченных животных проходили через районы добычи и транспортировки углеводородов, расположенные вдоль восточного побережья о-ва Сахалин). Перемещения к местам зимнего обитания заняли у лахтаков до 20 дней.
Четвертый период – время нахождения лахтаков на льду. При этом они предпочитают толстый однолетний лед высокой сплоченности (90–100%), хотя в акватории присутствует и лед меньшей сплоченности. Открытой воды животные избегают. Все лахтаки достигли мест своего зимнего обитания к середине января. Длительных быстрых переходов они больше не совершали. В этот период животные разделились на две группы по занимаемым глубинам. Сходным образом ведет себя лахтак и в Беринговом море, устраивая залёжки на льду с концентрацией более 75% [44]. Сплошного припайного льда лахтак, как и в других районах [45], избегает.
Следует отметить, что в годовом жизненном цикле лахтака в Охотском море в отличие от основной части его ареала, где этот вид тесно связан с ледовой кромкой и в течение года перемещается вслед за ней, в связи с полным освобождением моря ото льда в летнее время появляются дополнительные периоды – период формирования ледового покрова и быстрых перемещений к зимним местообитаниям. У лахтаков арктического региона выделяют только нагульный (летний) и репродуктивный (зимний) периоды [1].
Спутниковые передатчики позволили нам проследить пути перемещений лахтаков между нагульными и репродуктивными местообитаниями в Охотском море. Размножение лахтаков здесь приходится на конец марта–апрель (с пиком в первой половине апреля) [13, 14]. Помеченные нами лахтаки, летом нагуливающиеся в районе о-ва Чкалова в Сахалинском заливе, к зимнему периоду размножения разделились на три группы: одни остались в западной части Сахалинского залива, другие ушли к северной оконечности о-ва Сахалин, третьи – на восточное его побережье. Репродуктивное скопление лахтаков на восточном побережье о-ва Сахалин ранее было описано [21], а нахождение их в Сахалинском заливе зимой не отмечалось, хотя о перемещении туда лахтаков с севера и востока о-ва Сахалин по мере разрушения ледового покрова было известно [22].
Возвращение после сезона размножения к летним местообитаниям нам удалось зарегистрировать у одной особи лахтака (№ 66979). Эта половозрелая самка, активно перемещавшаяся до 25.03.2014 г., в течение следующего месяца (до 20.04.2014 г.) регистрировалась на очень небольшом участке к северу от о-ва Сахалин, а затем двинулась по направлению к центральной части Сахалинского залива, прошла за неделю 187 км, а 28.04.2014 г. ее передатчик прекратил работу. По-видимому, в период с 25.03.2014 г. по 20.04.2014 г. она родила и выкормила щенка (такой срок, около 24 дней, совпадает с известной продолжительностью лактации у этого вида [5]), а с разрушением ледового покрова она начала возвращаться к летним местообитаниям у о-ва Чкалова.
На всем ареале лахтак обычно использует акваторию в пределах 200-метровой изобаты [39, 45, 46]. В Охотском море помеченные нами лахтаки в течение всего времени прослеживания практически не регистрировались в водах с глубинами более 100 м: только 3.6% их локаций вышли за эту изобату. В некоторых районах Арктики лахтаки ведут себя таким же образом [47].
Таким образом, основными летними местообитания лахтаков, нагуливающихся в районе о-ва Чкалова, являются прибрежные районы Сахалинского залива. После становления льда все животные совершили протяженные миграции к зимним местообитаниям: в оффшорную часть Сахалинского залива, а также к северному и восточному побережьям о-ва Сахалин. Основными факторами, оказавшими влияние на сроки и характер перемещений лахтаков и выбор ими сезонных местообитаний, оказались толщина, сплоченность льда, даты становления ледового покрова и глубины акватории.
Полевая часть работы выполнена в рамках деятельности Постоянно действующей экспедиции РАН по изучению животных Красной книги Российской Федерации и других особо важных животных фауны России, а также при финансовой поддержке РФФИ (грант № 14-05-31440 мол_а).
Список литературы
Cameron M.F., Bengtson J.L., Boveng P.L. et al. Status review of the bearded seal (Erignathus barbatus) // U.S. Department of Commerce, NOAA Technical Memorandum NMFS-AFSC-211, 2010. 246 p.
Burns J.J. Remarks on the distribution and natural history of pagophilic pinnipeds in the Bering and Chukchi Seas // Journal of Mammalogy. 1970. V. 51. № 3. P. 445–454.
Fay F.H. The role of ice in the ecology of marine mammals of the Bering Sea // Oceanography of the Bering Sea. 1974. P. 383–399.
Nelson R.R., Burns J.J., Frost K.J. The bearded seal (Erignathus barbatus) // Marine mammal species accounts, wildlife technical bulletin. 1984. V. 7. P. 1–6.
Lydersen C., Kovacs K.M. Behaviour and energetics of ice-breeding, North Atlantic phocid seals during the lactation period // Marine Ecology Progress Series. 1999. V. 187. P. 265–281.
Moore S.E., Huntington H.P. Arctic marine mammals and climate change: impacts and resilience // Ecological Applications. 2008. V. 18. № sp2. P. S157–S165.
Poloczanska E.S., Brown C.J., Sydeman W.J. et al. Global imprint of climate change on marine life // Nature Climate Change. 2013. V. 3. № 10. P. 919–925.
Laidre K.L., Stern H., Kovacs K.M. et al. Arctic marine mammal population status, sea ice habitat loss, and conservation recommendations for the 21st century // Conservation Biology. 2015. V. 29. № 3. P. 724–737.
Lowry L.F., Frost K.J., Burns J.J. Feeding of bearded seals in the Bering and Chukchi Seas and trophic interaction with Pacific walruses // Arctic. 1980. P. 330–342.
Kovacs K.M., Lydersen C., Overland J.E., Moore S.E. Impacts of changing sea-ice conditions on Arctic marine mammals // Marine Biodiversity. 2011. V. 41. № 1. P. 181–194.
Richardson W.J., Greene C.R. Jr., Malme C.I., Thomson D.H. Marine mammal hearing // Marine Mammals and Noise. 1995. P. 205–240.
Фрейман С.Ю. Материалы к промысловой биологии тюленей Дальнего Востока // Труды ВНИРО. 1935. Т. 3. С. 188–203.
Тихомиров Э.А. Распределение и миграции тюленей в водах Дальнего Востока // Труды совещания по биологии и промыслу морских млекопитающих. 1961. № 2. С. 110–123.
Крылов В.И., Федосеев Г.А., Шустов А.П. Ластоногие Дальнего Востока. М.: Пищевая пром-ть, 1964. 59 с.
Тихомиров Э.А. О распределении и биологии ластоногих Берингова моря // Труды ВНИРО. 1964. Т. 53. С. 277–285.
Федосеев Г.А. О состоянии запасов и рациональном использовании тюленей Охотского моря // Изв. ТИНРО. 1966. Т. 58. С. 129–135.
Наумов С.П. Ластоногие Охотского моря // Уч. зап. Московского педагогич. ин-та. 1941. Т. 24. № 2. С. 19–74.
Федосеев Г.А., Бухтияров Ю.А. Питание тюленей Охотского моря // Тез. докл. V Всесоюз. совещ. по изучению морских млекопитающих. Ч. 1. Махачкала, 1972. С. 110–112.
Болтунов А.Н., Алексеева Я.И., Беликов С.Е. и др. Морские млекопитающие и белый медведь Карского моря: обзор современного состояния. М.: Печатный центр “Декарт”, 2015. 103 с.
Hammill M.O., Kovacs K.M., Lydersen C. Local movements by nursing bearded seal (Erignathus barbatus) pus in Kongsfjorden, Svalbard // Polar Biology (Print). 1994. V. 14. № 8. P. 569–570.
Федосеев Г.А. Популяционная биология ледовых форм тюленей и их роль в экосистемах Северной Пацифики. Магадан: МагаданНИРО, 2005. 179 с.
Гептнер В.Г., Чапский К.К., Арсеньев В.А., Соколов В.Е. Млекопитающие Советского Союза. Т. 2, ч. 3. Ластоногие и зубатые киты. М.: Высшая школа, 1976. 718 с.
Пихарев Г.А. Тюлени юго-западной части Охотского моря // Изв. ТИНРО. 1941. Т. 20. С. 61–99.
Потелов В.А. Распределение и миграции морского зайца в Белом, Баренцевом и Карском морях // Морские млекопитающие. М.: Наука, 1969. С. 245–251.
Simpkins M.A., Hiruki-Raring L.M., Sheffield G. et al. Habitat selection by ice-associated pinnipeds near St. Lawrence Island, Alaska in March 2001 // Polar Biology. 2003. V. 26. № 9. P. 577–586.
Frost K.J., Whiting A., Cameron M.F., Simpkins M.A. Habitat use, seasonal movements and stock structure of bearded seals in Kotzebue Sound, Alaska // Tribal Wildlife Grants Program, Fish and Wildlife Service, Tribal Wildlife Grants Study U-4-IT. Final report from the Native Village of Kotzebue, Kotzebue, AK, for US Fish and Wildlife Service, Anchorage, AK. 2008. P. 1–16.
Тихомиров Э.А. Рост тела и развитие органов размножения северотихоокеанских настоящих тюленей // Изв. ТИНРО. 1968. Т. 62. С. 216–243.
Слепцов М.М. О биологии размножения ластоногих Дальнего Востока // Зоол. журн. 1943. Т. 12. Вып. 2. С. 109–128.
Lopez R., Malardé J.P. Improving ARGOS Doppler location using Kalman filtering. CLS-DT-MEMO-11-65. 2011. 14 p.
R Core Team. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. 2019. URL https://www.R-project.org/
Boveng P.L., Bengtson J.L., Buckley T.W. et al. Status Review of the Spotted Seal (Phoca largha). U.S. Dep. Commer., NOAA Tech. Memo. NMFS-AFSC-200, 2009. 153 p.
Freitas C., Kovacs K.M., Ims R.A. et al. Ringed seal post-moulting movement tactics and habitat selection // Oecologia. 2008. V. 155. № 1. P. 193–204.
Hayne D.W. Calculation of size of home range // J. Mammal. 1949. V. 30. P. 1–18.
Worton B.J. Kernel methods for estimating the utilization distribution in home-range studies // Ecology. 1989. V. 70. P. 164–168.
Powell R.A. Animal home ranges and territories and home range estimators // Research techniques in animal ecology: controversies and consequences. New York: Columbia University Press, 2000. № 15. P. 65–110.
Breed G.A., Cameron M.F., Ver Hoef J.M. et al. Seasonal sea ice dynamics drive movement and migration of juvenile bearded seals Erignathus barbatus // Marine Ecology Progress Series. 2018. V. 600. P. 223–237.
Cameron M.F., Frost K.J., Ver Hoef J.M. et al. Habitat selection and seasonal movements of young bearded seals (Erignathus barbatus) in the Bering Sea // PloS one. 2018. V. 13. № 2. e0192743.
Johnson M.L., Fiscus C.H., Ostenson B.T., Barbour M.L. Marine mammals // Environment of the Cape Thompson Region, Alaska. U.S. Atomic Energy Commission, Oak Ridge, TN. 1966. P. 877–924.
Kovacs K.M. Bearded Seal: Erignathus barbatus // Encyclopedia of marine mammals. Academic Press, 2018. C. 83–86.
Чернявский В.И. Циркуляционные системы Охотского моря // Изв. ТИНРО. 1981. Т. 105. С. 13–19.
Потелов В.А. Питание морского зайца // Материалы рыбохоз. исслед. 1971. № 18. С. 107–121.
Бухтияров Ю.А. Питание тюленей в южной части Охотского моря // Изв. ТИНРО. 1990. Т. 112. С. 96–102.
Ледовые условия и методы их прогнозирования в Охотском море // Монографический справочник. Проект “Моря”. Гидрометеорология и гидрохимия морей. Т. IX. Охотское море. 1998. № 1. С. 291–331.
Ver Hoef J.M., Cameron M.F., Boveng P.L. et al. A spatial hierarchical model for abundance of three ice-associated seal species in the eastern Bering Sea // Statistical Methodology. 2014. V. 17. P. 46–66.
Burns J.J., Frost K.J. The natural history and ecology of the bearded seal, Erignathus barbatus // Environmental Assessment of the Alaskan Continental Shelf, Final Reports. 1979. V. 19. P. 311–392.
Косыгин Г.М., Трухин А.М., Велижанин А.Г. Зимнее распределение тюленей в Охотском море // Морские млекопитающие Дальнего востока. Владивосток: ТИНРО, 1984. С. 99–107.
Finley K.J., Renaud W.E. Marine mammals inhabiting the Baffin Bay North Water in winter // Arctic. 1980. P. 724–738.
Дополнительные материалы отсутствуют.