

Экология, 2021, № 6, стр. 430-438
Скорость роста щечных зубов узкочерепной полевки (Lasiopodomys gregalis) в зависимости от абразивности корма
Ю. Э. Кропачева a, *, Н. Г. Смирнов a, С. В. Зыков a
a Институт экологии растений и животных УрО РАН
620144 Екатеринбург, ул. 8 Марта, 202, Россия
* E-mail: KropachevaJE@yandex.ru
Поступила в редакцию 06.10.2020
После доработки 09.12.2020
Принята к публикации 22.12.2020
Аннотация
Изучена скорость роста первого нижнего моляра и особенности его стачивания у узкочерепных полевок (n = 39), получавших в лабораторных условиях жесткие и мягкие корма. В результате введения тетрациклиновой метки установлено, что за сутки высота зуба увеличивается на 0.33–0.56 мм. У полевок, получавших мягкий корм, по сравнению с животными, получавшими жесткий корм, наблюдалась меньшая скорость роста моляра, более низкая коронка, более тупой угол стачивания жевательной поверхности и фасетки боковых стираний. По мере развития этих признаков у животных, получавших мягкий корм, скорость роста моляра увеличивалась.
Зубная система растительноядных млекопитающих представляет собой комплекс взаимодействующих биомеханических структур, настроенный на переработку больших масс грубых кормов. Родовая и видовая специфика их реализации в щечных зубах проявляется в соотношении эмалевых, дентиновых и цементных элементов – их формы, количества и взаиморасположения. На эти особенности исследователи обращают внимание при решении задач происхождения и родства таксонов разного уровня, включая внутривидовой. Особое внимание уделяется эмалевому рисунку жевательной поверхности и другим особенностям, в изменчивости которых установлена высокая наследственная компонента [1, 2]. В тоже время зубная система имеет ряд характеристик, особенности которых формируются как реакция на средовые воздействия. Они могут носить адаптивный или патологический характер и быть обратимыми. Именно этот класс характеристик привлекает внимание в качестве индикатора кормовых условий, особенно в тех областях исследований, где затруднена или невозможна их прямая оценка, прежде всего в палеоэкологии.
Разработка косвенных методов реконструкций питания животных, основанных на особенностях стачивания жевательной поверхности зубов, является перспективным направлением исследований, так как позволяет значительно расширить возможности палеоэкологических реконструкций. Разные таксономические и экологические группы млекопитающих обладают существенной спецификой роста и стачивания зубов.
Объектом наших исследований была узкочерепная полевка (Lasiopodomys gregalis). Этот вид широко распространен в лесостепных и степных сообществах Евразии, и ее остатки представлены массовым одонтологическим материалом в палеонтологических сборах из местонахождений позднего плейстоцена и голоцена по всей Европе и Северной Азии [3, 4].
Коренные зубы серых полевок относятся к типу гипселодонтных, характеризуются отсутствием корней и обладают постоянным ростом и стиранием. Ранее были выявлены характерные особенности стачивания зубов узкочерепных полевок, содержавшихся на различных по абразивным свойствам кормах. Они проявляются в различиях величины угла стачивания жевательной поверхности, высоте зуба над альвеолами, появлении фасеток боковых стираний, особенностях микро- и мезорельефа жевательной поверхности [5–7]. Для данного вида показаны вариации в характеристиках мезорельефа верхних щечных зубов на трех участках вдоль широтного градиента экологических подзон полуострова Ямал в Российской Арктике [8].
В ряде работ [9–11] показано, что скорость роста зубов, не имеющих корней (как щечных, так и резцов), зависит от физических свойств употребляемого корма и замедляется на малоабразивных диетах. Выдвинуто предположение [8], что у узкочерепных полевок данная закономерность также проявляется. Настоящая работа посвящена проверке этой гипотезы. Данные о скорости роста щечных зубов необходимы для понимания процесса образования вариантов стачивания зубов в ответ на употребление пищи разной абразивности и какой временно́й интервал отражают разные варианты стачиваний.
Цель работы – оценить скорость роста первого нижнего щечного зуба (m/1) узкочерепных полевок, содержавшихся на мягком и жестком корме; сопоставить данные по скорости роста зубов с характеристиками стачивания коронки зуба – ее высоте над альвеолами, углу стачивания жевательной поверхности и выраженностью фасеток боковых стираний, а также с опубликованными ранее данными по микро- и мезорельефу жевательной поверхности у этих же животных.
МАТЕРИАЛ И МЕТОДЫ
Данные получены в результате эксперимента, проведенного на узкочерепных полевках (n = 46) из лабораторной колонии ИЭРиЖ УрО РАН в 2015 г. Животные-основатели колонии были отловлены в Белоярском р-не Свердловской обл., что соответствует области распространения подвида L. g. gregalis [12]. Целью этого эксперимента было выявление вариантов стачиваний зубов животных при употреблении кормов разной абразивности. В результате было показано, что у полевок, получавших малоабразивный корм, площадь повреждения эмали была больше, более низкий мезорельеф, меньшая высота коронки и более тупой угол стачивания, чем у животных, получавших более абразивный корм. Подробное описание эксперимента и результаты изучения микро- и мезорельефа опубликованы ранее [5, 6]. Для исследования скорости роста зубов использована часть выборки черепов животных, полученных в результате этого эксперимента (n = 39), пригодных по сохранности к измерению тетрациклиновой метки. Структура экспериментальных групп приведена в табл. 1. Полевки родились в мае и июне. Животных из каждого помета (9 пометов, 7 материнских линий) распределяли между двумя экспериментальными группами, по возможности соблюдалось равное соотношение полов. Эксперимент начинали в возрасте 1 мес. Ежемесячно у животных измеряли длину и массу тела.
Таблица 1.
Структура экспериментальных групп
| Экспериментальная группа | Мягкий корм | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Длительность содержания на диете, мес. | 1 | 2 | 3 | ||||||||
| Возраст на момент окончания эксперимента, мес. | 2 | 3 | 4 | ||||||||
| № материнской линии (n особей) | 1(3) | 2(2) | 1(1) | 3(2) | 4(3) | 5(1) | 1(2) | 3(1) | 5(3) | 7(2) | |
| № помета (n особей) | 20(3) | 22(2) | 17(1) | 16(2) | 18(3) | 19(1) | 17(2) | 16(1) | 15(3) | 14(2) | |
| Половой состав: самцы – 1, самки – 2 (n особей) | 1(2), 2(1) | 1(1), 2(1) | 2(1) | 1(1), 2(1) | 1(2), 2(1) | 2(1) | 1(1), 2(1) | 2(1) | 1(2), 2(1) | 1(2) | |
| Экспериментальная группа | Жесткий корм | ||||||||||
| Длительность содержания на диете, мес. | 1 | 2 | 3 | ||||||||
| Возраст на момент окончания эксперимента, мес. | 2 | 3 | 4 | ||||||||
| № материнской линии (n особей) | 1(2) | 2(1) | 9(2) | 3(2) | 4(3) | 5(1) | 1(2) | 3(1) | 5(3) | 7(2) | |
| № помета (n особей) | 20(2) | 22(1) | 21(2) | 16(2) | 18(3) | 19(1) | 17(2) | 16(1) | 15(2) | 19(1) | 14(2) |
| Половой состав: самцы – 1, самки – 2 (n особей) | 1(2) | 2(1) | 1(1), 2(1) | 1(1), 2(1) | 1(1), 2(2) | 2(1) | 1(1), 2(1) | 1(1) | 1(2) | 1(1) | 1(1), 2(1) |
Животных содержали в пластиковых контейнерах, закрытых мелкой металлической сеткой, в качестве подстилки использовали опилки. Применяли два состава кормов, различающихся по содержанию абразивных элементов, которым соответствовали две экспериментальные группы. Мягкий корм (n = 20) был максимально освобожден от внешних абразивов (частицы почвы, пыль) и содержал компоненты с низким содержанием внутренних абразивов (фитолиты растений): отмытые от пыли и частиц почвы листья одуванчиков, очищенная от частиц почвы путем снятия перидермы морковь, яблоки без сердцевины. Пища второй группы – жесткий корм (n = 19) – включала компоненты с высоким содержанием фитолитов растений и плотные компоненты: листья однодольных растений, сено, а также неочищенная морковь. С жестким кормом естественным образом могли попадать как внутренние, так и незначительное количество внешних абразивов (частицы почвы).
Рост коронки m/1 в высоту оценивали путем введения тетрациклиновой метки подкожно из расчета 20–25 мг/кг [13]. Через 15 дней после инъекции животных умерщвляли. Скорость роста пересчитывали на одни сутки (мм/сут). При исследовании зуба в ультрафиолетовом свете участки, которые росли в период циркуляции тетрациклина в организме, имеют желтую флуоресценцию. Положение тетрациклиновой метки измеряли по фотографиям от ее верхнего края до базальной части зуба вдоль третьего буккального выходящего угла – BSA3 (здесь и далее обозначения по [1]) (рис. 1, 1). Во избежание повреждения зуба и нарушения его положения в челюсти мы не вынимали его из альвеол. Чтобы сфотографировать метку, выпиливали кусок нижнечелюстной кости с помощью стоматологического бура и обнажали нижние 2/3 зуба. Часть зуба для измерения метки (BSA3) была выбрана из-за ее наибольшей доступности при таком подходе.
Рис. 1.
Схема промеров первого нижнего моляра (m/1): 1 – положение тетрациклиновой метки; 2 – угол стачивания жевательной поверхности; 3, 4 – высота коронки зуба над альвеолами, измеренная по BSA1 и BSA3 соответственно; 5 – высота зуба от базальной части до жевательной поверхности, измеренная по BSA3.
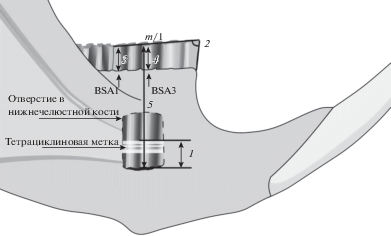
Угол стачивания жевательной поверхности измеряли относительно передней поверхности моляра (в градусах) (рис. 1, 2). Высоту коронки зуба над альвеолами измеряли с буккальной стороны от жевательной поверхности до края альвеол в двух местах – вдоль углов BSA1 и BSA3 (рис. 1, 3, 4). Были взяты два промера для установления взаимосвязи высоты передней и задней частей зуба с углом стачивания и закономерностей его формирования. Высоту зуба измеряли с буккальной стороны от жевательной поверхности до базальной части зуба вдоль BSA3 (рис. 1, 5). На основе данного промера высчитано теоретическое время обновления тканей зуба при условии постоянной скорости роста и стачивания.
Исследовали также фасетки боковых стираний – аномальное проявление стачивания зубов полевок, возникающее в результате нарушения окклюзии при употреблении мягких кормов. Фасетки встречаются как у животных, содержавшихся в лабораторных условиях, так и у полевок из природы [7]. Они располагаются на внутренних выходящих углах нижних моляров и внешних выходящих углах верхних моляров, что свидетельствует о возникновении фасеток боковых стираний в результате трения зубов друг о друга. Развитие боковых стираний, как правило, происходит от передней к задней части зуба и от передних к задним молярам (чаще фиксируются на m/1 и M1/) [7]. Степень развития фасеток оценена в баллах: 1 – начальная стадия, затрагивает только кромку жевательной поверхности зуба; 2 – стирание распространяется в верхней четверти боковой поверхности моляра; 3 – стиранием затронуто более 1/4 части моляра.
Челюсти полевок фотографировали в боковой проекции через бинокулярный микроскоп Leica EZ4. Измерения произведены с помощью пакета программ TPS (TPS Util и TPS Dig2).
Для оценки статистической значимости результатов сравнения угловых, размерных характеристик и скорости роста зубов животных из двух экспериментальных групп применен двухфакторный дисперсионный анализ (ANOVA, F) с факторами “экспериментальная группа” (мягкий и жесткий корм) и “длительность содержания на диете”. Для оценки возможного вклада наследственной компоненты в изменчивость скорости роста зубов использован двухфакторный дисперсионный анализ с факторами “материнская линия” и “экспериментальная группа”. Связи между характеристиками зубов в каждой из экспериментальных групп выявляли с помощью корреляционного анализа Пирсона (r). Для определения связи между угловыми, размерными характеристиками, скоростью роста моляра и степенью выраженности фасеток боковых стираний в группе животных, получавших мягкий корм, использован дисперсионный анализ с фактором “степень выраженности фасеток в баллах”. Статистический анализ данных проведен в программе Statistica 7.0.
РЕЗУЛЬТАТЫ
Длина и масса тела статистически не различались у животных из экспериментальных групп, получавших разный корм (фактор “экспериментальная группа”). Длина тела значимо увеличивалась в группах, содержавшихся на диетах разное количество времени (“длительность содержания на диете”) (F(2;33) = 6.47, p < 0.05).
Скорость роста m/1 у узкочерепных полевок, содержавшихся на мягком корме, была статистически значимо меньше, чем у животных, получавших жесткий корм (фактор “экспериментальная группа”) (F(1;33) = 23.27, p < 0.001) (табл. 2). Если допустить, что скорости прироста коронки и ее стачивания будут сохраняться постоянными, то полное обновление тканей зуба на мягком корме происходит в среднем за 75.7 сут (от 48.9 до 97.3 сут), на жестком корме – 65.1 сут (от 55.5 до 81.5 сут). Различия между скоростью роста зубов животных, которых содержали на диетах разное время, также статистически значимы (фактор “длительность содержания на диете” (F(2;33) = = 7.37, p < 0.05). Для обеих групп максимальная скорость роста наблюдается в группе животных, которых содержали на диетах 2 мес. (возраст зверей – 3 мес.) (рис. 2а). Двухфакторный дисперсионный анализ с факторами “экспериментальная группа” и “материнская линия” показал статистически значимые различия между экспериментальными группами (F(1;25) = 8.30, p < 0.05), а различия между материнскими линиями (F(5;25) = = 0.45, p = 0.81) и взаимодействие этих факторов (F(5;25) = 0.60, p = 0.70) статистически незначимы. На основании этих данных можно сделать вывод об отсутствии вклада наследственной компоненты в изменчивость скорости роста зубов.
Таблица 2.
Характеристики m/1 и тела узкочерепных полевок, содержавшихся на мягком (МК, n = 20) и жестком (ЖК, n = 19) корме
| Характеристика | Диета | M ± Std Dev. | min | max |
|---|---|---|---|---|
| Скорость роста m/1, мм/сут | МК | 0.040 ± 0.0058 | 0.033 | 0.056 |
| ЖК | 0.046 ± 0.0035 | 0.041 | 0.053 | |
| Высота m/1 над альвеолами, мм: | ||||
| BSA1 | МК | 0.74 ± 0.076 | 0.60 | 0.85 |
| ЖК | 0.82 ± 0.07 | 0.70 | 1.0 | |
| BSA3 | МК | 0.75 ± 0.09 | 0.60 | 0.94 |
| ЖК | 0.86 ± 0.07 | 0.75 | 1.05 | |
| Высота m/1, мм: BSA3 | МК | 2.96 ± 0.25 | 2.55 | 3.45 |
| ЖК | 2.99 ± 0.19 | 2.65 | 3.40 | |
| Угол стачивания жевательной поверхности m/1, град | МК | 78.65 ± 3.31 | 72.0 | 85.0 |
| ЖК | 72.42 ± 2.95 | 66.00 | 76.00 | |
| Масса тела, г | МК | 15.71 ± 3.40 | 11.10 | 22.50 |
| ЖК | 17.24 ± 3.82 | 13.00 | 26.80 | |
| Длина тела, мм | МК | 102.05 ± 6.10 | 95.00 | 116.00 |
| ЖК | 101.42 ± 4.20 | 92.00 | 115.00 |
Рис. 2.
Характеристики зубов узкочерепных полевок, содержавшихся на двух диетах разное время (средние значения и 95%-ный доверительный интервал): 1 – жесткий корм; 2 – мягкий корм.
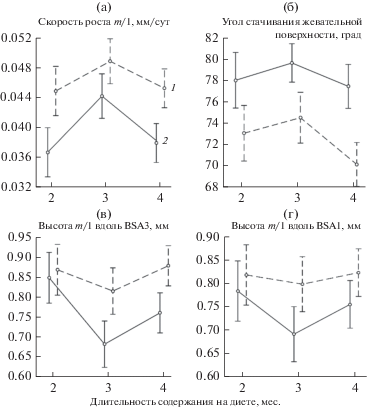
Угол стачивания жевательной поверхности статистически значимо больше (более тупой) у животных, получавших мягкие корма (фактор “экспериментальная группа”) (F(1;33) = 40.72, p < < 0.001). Фактор “длительность содержания на диете” также статистически значим (F(2;33) = = 5.29, p < 0.05) (рис. 2б).
Коронка зуба, измеренная как по BSA1, так и по BSA3, ниже у животных, получавших мягкие корма (фактор “экспериментальная группа”) (F(1;33) = 9.86, p < 0.05 и F(1;33) = 16.68, p < 0.001 соответственно) (см. табл. 1, рис. 2в, г). Фактор “длительность содержания на диете” статистически значим для BSA3 (F(2;33) = 8.03, p < 0.05) (рис. 2в).
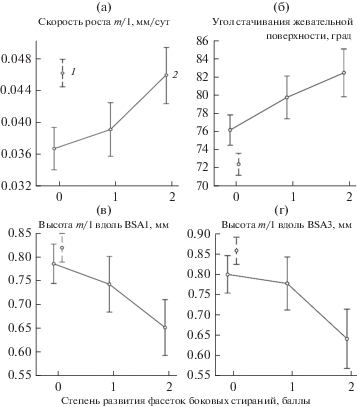
Фасетки боковых стираний обнаружены только у зверей, получавших мягкий корм (у 12 из 20 особей). Степень развития фасеток – 1, 2 и 3 балла. В ходе дальнейшего анализа зубы со степенью развития фасеток 2 и 3 были объединены в одну группу (2 балла) вследствие небольшого количества материала в каждой выборке.
Взаимосвязь характеристик роста и стачивания моляров в двух экспериментальных группах. В группе, получавшей жесткий корм, не выявлено значимых корреляций между скоростью роста зубов, высотой коронки и углом стачивания жевательной поверхности. В группе, которая получала мягкий корм, выявлена статистически значимая отрицательная корреляция между скоростью роста моляра и высотой коронки, измеренной как по BSA1, так и по BSA3 (r = –0.47, r = –0.44, p < 0.05), положительная между скоростью роста и значением угла стачивания жевательной поверхности (r = 0.47, p < 0.05), отрицательная – между высотой коронки, измеренной вдоль BSA1, и значением угла стачивания жевательной поверхности (r = = –0.55, p < 0.05). Таким образом, при более высокой скорости роста наблюдаются более низкая коронка и тупой угол стачивания. По мере развития фасеток от 0 до 2 баллов увеличивается скорость роста зубов (F(2;17) = 7.29, p < 0.05) (рис. 3а). Угол стачивания становится более тупым (F(2;17) = 16.26, p < 0.001) (рис. 3б), коронка зуба, измеренная по BSA1 и BSA3, – более низкой (F(2;17) = 9.55, p < 0.05 и F(2;17) = 8.63, p < 0.05 соответственно) (рис. 3в, г).
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Скорость роста m/1. Результаты оценки скорости роста m/1 были сопоставлены с литературными данными по скорости роста этого зуба у других видов грызунов, полученными с помощью тетрациклиновой метки. Средняя скорость роста m/1 у узкочерепной полевки медленнее, чем у других некорнезубых полевок, у которых во входящих углах коренных зубов откладывается цемент [14, 15].
При сравнении животных, получавших разные корма, выявлено, что в случае жесткого корма скорость роста зубов была выше. Масса тела не различалась у животных, содержавшихся на разных диетах, следовательно, более низкая скорость роста зубов у зверей, получавших мягкий корм, не обусловлена недостатком питательных веществ. Рост и развитие животных, рожденных в лабораторных условиях, так же, как и в природных, имеют сезонную изменчивость. Все животные, участвовавшие в эксперименте, родились в лаборатории в течение двух месяцев – в мае и июне, следовательно, влияние сезона рождения на ростовые процессы было сходно у всех особей. Не выявлено вклада наследственной компоненты в изменчивость скорости роста зубов. Таким образом, наиболее вероятно, что наблюдаемые различия роста зубов в двух экспериментальных группах обусловлены разным составом кормов, которые вызвали разную нагрузку на моляр при жевании.
Характеристики стачиваний m/1. Высота зубов с постоянным ростом определяется как скоростью роста, так и скоростью стирания. У узкочерепных полевок, содержавшихся на мягком корме, наблюдалась более низкая коронка. Это могло произойти вследствие, как минимум, двух взаимосвязанных причин: во-первых, низкая коронка могла формироваться от того, что скорость роста зубов на мягкой диете была меньше, во-вторых, из-за более интенсивного износа зубов. Высокоабразивная диета в нашем эксперименте была сходна с природным рационом (хоть и не аналогична ему из-за присутствия моркови). Ранее было показано сходство характеристик мезорельефа и высоты коронки у животных, получавших эту диету, и у животных, отловленных в природе [5]. Малоабразивная диета, вероятно, была экстремально мягкой для данного вида. У узкочерепных полевок, содержавшихся на этой диете, наблюдались более низкая коронка, более тупой угол стачивания и фасетки боковых стираний, которые возникают при трении боковых поверхностей зубов друг о друга. Ранее было показано [6], что у животных, получавших малоабразивный корм, площадь повреждения эмали в результате ее скалывания была больше, чем у животных, получавших более абразивный корм. Таким образом, характеристики стачиваний свидетельствуют о сильном износе тканей зуба, что при малом количестве абразивных компонентов в пище могло возникнуть только как результат трения зубов друг о друга.
Различия значений углов стачивания при содержании полевок на двух диетах, вероятно, связаны с особенностями биомеханики жевательных движений, направленных спереди назад, распределением пищевых частиц в ротовой полости и разной нагрузкой при жевании на переднюю и заднюю части зуба [12, 17, 18]. Кроме того, угол становится более тупым по мере развития фасеток, распространение которых начинается с передней части зуба. Можно предположить, что нарушение окклюзии усиливает затупление угла стачивания.
Длительность содержания на диете. Отмечены различия исследованных характеристик у животных, содержавшихся на двух диетах разное количество времени. Эти изменения нелинейны, но синхронны в обеих группах, что исключает их случайность. У 3-месячных зверей, содержавшихся на диетах 2 мес., в обеих экспериментальных группах наблюдались большая скорость роста зуба, меньшая высота коронки, более тупой угол стачивания. Обращают на себя внимание направления изменений признаков внутри групп, содержавшихся на разных диетах: при повышении скорости роста увеличивается угол стачивания и уменьшается высота коронки.
Интересно то, что и в группе, получавшей мягкий корм, были особи, имеющие острый угол стачивания, высокую коронку и половина животных из этой группы не имела фасеток боковых стираний. Это могло произойти вследствие предпочтения полевками какого-то одного компонента корма – моркови либо одуванчиков, и, что более вероятно, из-за адаптации части зверей к такому корму за счет особенностей анатомии и биомеханики жевательных движений. Обе эти гипотезы требуют проверки в специальных исследованиях.
Взаимосвязь изученных характеристик m/1 у животных, получавших мягкий корм, и сопоставление с литературными данными. Внутригрупповой анализ, проведенный для животных, получавших мягкий корм, выявил тенденцию к увеличению скорости роста коронки по мере возрастания износа зубов, вероятно, вследствие стачивания зуб о зуб. Крайним проявлением такого типа стачивания является возникновение фасеток боковых стираний. Развитие фасеток сочетается с большей скоростью роста зуба, более тупым углом стачивания и более низкой коронкой зуба.
Полученные нами данные были сопоставлены с результатами экспериментов, поставленных на гипселодонтных животных [9, 16, 19]. В исследовании, проведенном на морских свинках, показано, что при употреблении высокоабразивного корма (листья бамбука) зубы животных были ниже и имели более глубокие лунки в дентине, чем при менее абразивном корме (листья тимофеевки и люцерна) [19]. Узкочерепные полевки, получавшие жесткий корм, также имели более глубокий мезорельеф жевательной поверхности моляров, чем животные, содержавшиеся на мягком корме [5]. Однако высота зубов у узкочерепных полевок в случае жесткого корма была больше, так как жесткий корм не был экстремально абразивным для данного вида и не вызвал повышенного износа моляров.
В эксперименте, проведенном на кроликах [9, 16], показано, что при употреблении высокоабразивного корма, включающего внешний абразив (песок), зубы животных имели низкий и округлый мезорельеф жевательной поверхности и более выраженный микрорельеф, чем зубы животных, которые не получали внешний абразив (люцерна, листья однодольных растений и листья однодольных растений вместе с рисовой шелухой). При употреблении высокоабразивного корма скорость как роста, так и стирания была выше, чем в случае более мягких кормов. Коронка зубов у животных, которые получали песок с пищей, была ниже, чем у животных, которые его не получали, так как скорость роста не полностью компенсировала стирание [9, 16].
В нашем эксперименте высота, мезорельеф и микрорельеф зубов узкочерепных полевок, содержавшихся на мягком корме, носят некоторые черты сходства с таковыми у кроликов, получавших корм с песком [9, 16]: признаки сильного износа эмали [6], низкий мезорельеф [5], низкая коронка, а также признак, не обнаруженный у кроликов, – фасетки боковых стираний. Вероятно, при содержании на экстремально мягких кормах, вызывающих при жевании трение зуб о зуб, возникает тип стирания, сходный с сильным износом зубов, возникающим при содержании животных на экстремально жесткой диете, включающей внешний абразив, и преобладающем типе стирания зубов о пищу.
В случае мягкого корма в целом для экспериментальной группы отмечена более низкая скорость роста моляров, чем при содержании на жестком корме. По-видимому, малоабразивная диета провоцирует уменьшение нагрузки на зубы при жевании и вызывает снижение скорости их роста. В то же время в группе, получавшей мягкий корм, наблюдались признаки сильного износа тканей зуба. Степень проявления этих признаков была различной у разных особей. У животных, не имевших повышенного износа коронки, отмечена минимальная скорость роста зуба. Полевки, для которых отмечены признаки сильного износа моляра, имели более высокую скорость его роста. При максимальном износе скорость роста зуба приближалась к значению, полученному для животных, которые содержались на жесткой диете.
Таким образом, полученные данные свидетельствуют о том, что употребление крайне малоабразивной пищи, провоцировало снижение скорости роста моляра, но у части особей, вероятно, вызывало чрезмерное стирание зубов друг о друга. Это сопровождалось сильным износом зубов, что компенсировалось ускорением их роста. Скорость роста зубов полевок, получавших жесткий корм, была выше, чем в случае мягкого корма. Жесткий корм не провоцировал усиленного стирания зубов, так как не был экстремально абразивным для данного вида. Диапазон индивидуальных изменений скорости роста зубов и характера их стачиваний в ответ на употребление кормов разной абразивности можно оценить, исследуя одних и тех же животных при последовательной смене диет с разной абразивностью и прижизненной оценке изучаемых характеристик.
Наши результаты подтверждают гипотезу о том, что скорость роста коренных зубов узкочерепной полевки зависит от физических свойств корма. Проявление морфологических реакций зубов на отклонение от некой «нормальной» для вида пропорции кормов разной жесткости позволяет предположить, что существует видовая специфика характеристик щечных зубов, позволяющая перерабатывать корма определенного диапазона абразивности. Вероятно, выход за такой диапазон сказывается на характере стачиваемости жевательной поверхности и скорости роста зубов. Сравнительное изучение этих процессов у разных таксономических и экологических групп растительноядных млекопитающих является важной и актуальной задачей функциональной морфологии зубов, решение которой позволит существенно продвинуть возможности палеоэкологических реконструкций.
Авторы благодарят уважаемого рецензента за ценные замечания и П.В. Рудоискателя за предоставленное оборудование, без которого проведение исследования было бы невозможно.
Работа выполнена в рамках государственного задания Института экологии растений и животных УрО РАН при частичной поддержке РФФИ (проекты № 19-04-01008 и 19-04-00507).
Авторы подтверждают отсутствие конфликта интересов.
Список литературы
Бородин А.В. Определитель зубов полевок Урала и Западной Сибири (поздний плейстоцен–современность). Екатеринбург: УрО РАН, 2009. 100 с.
Маркова Е.А. Оценка сложности щечных зубов полевок (Arvicolinae, Rodentia): ранжированный морфотипический подход // Зоол. журн. 2013. Т. 92. Вып. 8. С. 968–980.
Смирнов Н.Г., Кузьмина Е.А., Головачев И.Б., Фадеева Т.В. Узкочерепная полевка (Microtus gregalis Pall.) в динамике зональных сообществ грызунов Северной Евразии // Экология. 2007. № 2. С. 117–123.
Smirnov N.G., Izvarin E.P., Kuzmina E.A., Kropacheva Y.E. Steppe species in the Late Pleistocene and Holocene small mammal community of the Urals // Quaternary International. 2016. V. 420. P. 136–144.
Kропачева Ю.Э., Сибиряков П.А., Смирнов Н.Г., Зыков С.В. Мезостачивания зубов серых полевок как индикаторы твердости и абразивности корма // Экология. 2016. № 6. С. 441–448.
Зыков С.В., Кропачева Ю.Э., Смирнов Н.Г. Микростачивание моляров узкочерепной полевки (Mic- rotus gregalis Pall., 1779) в зависимости от абразивности корма // Докл. РАН. 2018. Т. 478. № 3. С. 366–368.
Смирнов Н.Г., Кропачева Ю.Э. Распространение фасеток боковых стираний моляров у полевок (Arvicolinae) // Докл. РАН. 2015. Т. 460. № 1. С. 115–117.
Ungar P.S., Sokolova N.A., Purifoy J. et al. Assessing molar wear in narrow-headed voles as a proxy for diet and habitat in a changing Arctic // Mammalian Biology. 2020. P. 1–15. https://doi.org/10.1007/s42991-020-00079-x
Müller J., Clauss M., Codron D. et al. Growth and wear of incisor and cheek teeth in domestic rabbits (Oryctolagus cuniculus) fed diets of different abrasiveness // J. Exp. Zool. 2014. V. 321A. P. 283–298.
Müller J., Clauss M., Codron D. et al. Tooth length and incisal wear and growth in Guinea pigs (Cavia porcellus) fed diets of different abrasiveness // J. Anim. Physiol. Anim. Nutr. 2015. V. 99. P. 591–604.
Meredith A.L., Prebble J.L., Shaw D.J. Impact of diet on incisor growth and attrition and the development of dental disease in pet rabbits // J. of Small Animal Practice. 2015. V. 56. № 6. P. 377–382.
Громов И.М., Поляков И.Я. Млекопитающие. Л.: Наука, 1977. 504 с. (Фауна СССР. Т. 3, вып. 8).
Клевезаль Г.А. Регистрирующие структуры млекопитающих в зоологических исследованиях. М.: Наука, 1988. 288 с.
Голенищев Ф.Н., Кенигсвальд В. Скорость роста бескорневых зубов у Microtinae (Mammalia, Rodentia) // Функциональная морфология и систематика млекопитающих / Труды ЗИН. Л., 1978. Т. 79. С. 102–104.
Кропачева Ю.Э. Оценка скорости роста зубов полевки-экономки (Arvicolinae, Rodentia) // Экология: традиции и инновации: Мат-лы конф. молодых ученых. Екатеринбург, 2012. С. 66–69.
Martin L.F., Krause L., Ulbricht A. et al. Dental wear at macro-and microscopic scale in rabbits fed diets of different abrasiveness: A pilot investigation // Palaeogeography, Palaeoclimatology, Palaeoecology. 2020. V. 556. https://doi.org/10.1016/j.palaeo.2020.109886
Cox P.G., Rayfield E.J., Fagan M.J. et al. Functional evolution of the feeding system in rodents // PLoS One. 2012. V. 7. № 4. e36299.
Kesner M.H. Functional morphology of masticatory musculature of the rodent subfamily Microtinae // J. of Morphology. 1980. V. 165. P. 205–222.
Martin L.F., Winkler D., Tütken T. et al. The way wear goes: phytolith-based wear on the dentine–enamel system in guinea pigs (Cavia porcellus) // Proceedings of the Royal Society B. 2019. V. 286. №. 1912. https://doi.org/10.1098/rspb.2019.1921
Дополнительные материалы отсутствуют.