

Экология, 2021, № 6, стр. 403-411
Субфоссильные спорово-пыльцевые спектры лиственничных лесов Центральной Эвенкии: особенности интерпретации для целей палеоэкологических исследований
Е. Ю. Новенко a, b, *, Н. Г. Мазей a, Д. А. Куприянов a, Л. В. Филимонова c, Н. Б. Лаврова d
a Московский государственный университет им. М.В. Ломоносова
119991 Москва, Ленинские горы, 1, Россия
b Институт географии РАН
119017 Москва, Старомонетный переулок, 29, стр. 4, Россия
c Институт биологии Карельского научного центра РАН
185910 Петрозаводск, ул. Пушкинская, 11, Республика Карелия, Россия
d Институт геологии Карельского научного центра РАН
185910 Петрозаводск, ул. Пушкинская, 11, Республика Карелия, Россия
* E-mail: lenanov@mail.ru
Поступила в редакцию 16.10.2020
После доработки 23.02.2021
Принята к публикации 26.02.2021
Аннотация
Представлена характеристика 27 субфоссильных спорово-пыльцевых спектров современных растительных сообществ лиственничных лесов Центральной Эвенкии. Установлено, что, несмотря на преобладание лиственничных лесов в растительном покрове района исследования, основными компонентами спектров являются Betula sect. Betula и Alnus alnobetula subsp. fruticosa, тогда как доля пыльцы Larix занижена по отношению к ее участию в растительности и составляет от 0.6 до 13.5%. Доля пыльцы Picea возрастает в точках отбора проб, где ель входит в состав древостоя. Пыльца Pinus является заносным компонентом спорово-пыльцевых спектров, доля которой повышается в фитоценозах, где древесный ярус изрежен или отсутствует. Выявлены характерные виды таежных лесов, болотных и луговых местообитаний, индикаторы нарушенных грунтов и гарей, которые позволят более детально и объективно реконструировать палеорастительность.
Изучение растительности и условий окружающей среды прошлых эпох на основе палинологических данных – один из наиболее распространенных подходов в палеоэкологических исследованиях. Вопрос, насколько адекватно состав и соотношение компонентов спорово-пыльцевых спектров отражают растительные сообщества, их продуцирующие, чрезвычайно важен при реконструкции изменений растительности любого региона. Исследованию состава субрецентных и субфоссильных спорово-пыльцевых спектров посвящена обширная литература, начиная с первых работ [1–4] и до последних десятилетий [5–10]. Особенности формирования спорово-пыльцевых спектров в различных районах Сибири и Дальнего Востока детально рассмотрены в публикациях [11–14]. Изучению пыльцевого дождя и поверхностных проб в зонах тундры и лесотундры в Восточном секторе Российской Арктики посвящены работы [15–18]. При сопоставлении поверхностных спорово-пыльцевых спектров с составом растительных сообществ на севере Сибири, в Арктике и Субарктике выявлены определенные сложности в использовании этих данных при палеоэкологических реконструкциях [16, 19, 20]. Субфоссильные спорово-пыльцевые спектры лесных экосистем в криолитозоне Центральной и Восточной Сибири до настоящего времени практически не изучены из-за труднодоступности этих регионов и в целом небольшого количества палинологических работ.
История развития лиственничных лесов в прошлом и их современная динамика представляют большой интерес. В настоящее время эти леса занимают огромные пространства в Северной Евразии, а в период последнего оледенения значительно расширяли область своего распространения в перигляциальной зоне [21]. Из-за тонкой экзины пыльца лиственницы плохо сохраняется в отложениях, вследствие чего ее участие в ископаемых спорово-пыльцевых спектрах, как правило, занижено [22]. В связи с этим получение новых данных по составу современных спорово-пыльцевых спектров лиственничных лесов и степени адекватности их окружающей растительности очень значимо для интерпретации результатов палинологического анализа.
Цель настоящей работы – выявление региональных особенностей субфоссильных спорово-пыльцевых спектров лиственничных лесов в среднем течении р. Нижняя Тунгуска в Центральной Эвенкии. Для ее реализации были изучены поверхностные образцы, отобранные в различных типах лиственничных лесов, заболоченных рединах, пойменных растительных сообществах и на болотах.
МАТЕРИАЛ И МЕТОДЫ
Исследования проведены в центральной части Среднесибирского плоскогорья в среднем течении р. Нижняя Тунгуска (правый приток р. Енисей), в окрестностях пос. Тура. Абсолютные высоты местности составляет от 120 до 600 м над у. м., рельеф низкогорный, эрозионно-денудационный, климат умеренный, резко континентальный. По данным наблюдений метеостанции в пос. Тура (1936‒2018 гг.), температуры января, июля и среднегодовая составляют –35.7, +16.6 и –8.8°С соответственно [23]; осадков выпадет около 370 мм в год. Территория находится в пределах области сплошного распространения многолетней мерзлоты.
Район исследований принадлежит Нижне-Тунгусскому округу Ангаро-Тунгусской лесорастительной провинции Средне-Сибирской плоскогорной лесорастительной области [24]. Растительный покров образован преимущественно северотаежными светлохвойными лесами из Larix gmelinii (Rupr.) Rupr., находящимися в разных стадиях восстановительных постпирогенных сукцессий [25]. Межпожарный интервал зависит от рельефа и экспозиции склонов и варьирует от 61 до 139 лет [26, 27].
Для изучения субфоссильных спорово-пыльцевых спектров были отобраны поверхностные (1–2 см) слои почвы или мха (3–5 см) из центральной части моховых подушек. В местах отбора проведено детальное описание растительности [28]. Всего исследовано 27 растительных сообществ.
При подготовке проб для спорово-пыльцевого анализа их нагревали в 10%-ном растворе гидроксида калия до закипания, промывали дистиллированной водой через сито с размером ячеек 120 мкм для удаления крупных растительных остатков, а затем проводили ацетолиз [29]. Минеральные отложения дополнительно обрабатывали плавиковой кислотой для удаления силикатов. При проведении палинологического анализа в каждом образце подсчитывали не менее 500 пыльцевых зерен и спор.
Обработка данных и построение спорово-пыльцевых диаграмм проведены с использованием программ TILIA и TGView [30]. За 100% принята сумма пыльцы деревьев, кустарников (arboreal pollen, AP) и кустарничков, трав (non-arboreal pollen, NAP). Процентные соотношения спор подсчитаны относительно этой суммы.
РЕЗУЛЬТАТЫ
Локальная растительность в точках отбора поверхностных проб. Сомкнутые лесные сообщества представлены в четырех точках (т.) отбора: лиственничник голубично-зеленомошный с подлеском из Alnus alnobetula subsp. fruticosa (Rupr.) Raus. (т. 1), лиственничник багульниково-зеленомошный (т. 2) и лиственничник кустарничково-линнеевый с Picea obovata Ledeb и подлеском из Sorbus sp. (т. 3), которые расположены на склоне водораздела и на первой надпойменной террасе р. Нижняя Тунгуска, и березняк хвощевый, приуроченный к высокой пойме (т. 4). Сомкнутость крон древесного яруса в т. 1–4 составляет 0.5–0.6. Кустарниковый ярус образован в основном Alnus alnobetula subsp. fruticosa, Rosa acicularis Lindl., несколькими видами Salix: S. lapponum L., S. reticulata L., S. rosmarinifolia L. и др.
Проективное покрытие (ПП) травяно-кустарничкового яруса от 10 до 45%. Кустарнички представлены Ledum palustre L., Vaccinium vitis-idaea L. Среди трав наибольшую встречаемость имеют разные виды Carex sp., Equisetum arvense L., Е. palustre L., Calamagrostis langsdorffii (Link) Trin., С. lapponica (Wahlb.) Hartm. Мохово-лишайниковый покров образован различными видами зеленых мхов и лишайников, среди которых чаще всего встречаются Polytrichum commune Hedw., Pleurozium schreberi (Brid.) Mitt.
В редкостойных лиственничниках с сомкнутостью крон 0.3–0.4 отобраны пробы в 6 местообитаниях (т. 5–10). Места отбора проб 5, 6, 9, 10 расположены на первой надпойменной террасе Нижней Тунгуски, 7 и 8 – на склоне междуречья. Древесный ярус в этих сообществах формирует Larix gmelinii высотой 5–12 м. В точках отбора проб 7 и 8 отмечен подлесок из Alnus alnobetula subsp. fruticosa. В травяно-кустарничковом ярусе фоновым видом является Ledum palustre, но здесь же растут и другие кустарнички: Vaccinium uliginosum L., V. vitis-idaea, Betula nana L., Empetrum nigrum L. Травянистый ярус практически отсутствует (не более 3%). Мохово-лишайниковый ярус хорошо выражен и составляет до 95%: в т. 5 преобладают лишайники (80% ПП); в т. 7, 8, 10 – зеленые мхи (Polytrichum commune, Pleurozium schreberi и др.); в т. 6, 9 – сфагновые мхи.
В лиственничных рединах с сомкнутостью крон 0.1–0.2 исследованы 4 пробы (т. 11–14), в гари лиственнично-осоковой – одна проба (т. 15). Древесный ярус выражен слабо, высота деревьев (т. 11, 12, 14, 15) Larix gmelinii достигает 10 м. В точке отбора пробы 13 хорошо выражен подлесок из Alnus alnobetula subsp. fruticosa. Кустарниковый ярус образован различными видами ив. Среди кустарничков фоновым является Vaccinium uliginosum. Травянистый ярус в т. 11–13 слабо выражен (ПП до 8%), в т. 14 – ПП 35%, представлены Carex sp., Equisétum sp., Eriophorum sp., Poa sp., Hedysarum sp.
Пойменная растительность охарактеризована двумя точками в ивняках хвощево-вейниковых (т. 16, 17) и пробой, отобранной в ольховнике разнотравном (т. 18).
Для растительного покрова болот (т. 19–27) характерны мозаичность и комплексность, обусловленные различными типами микрорельефа болотных массивов [31]. При палеоэкологических исследованиях ископаемые пыльца и споры в основном выделяются из торфяных колонок, поэтому формирование спорово-пыльцевых спектров на болотах требует тщательного изучения. Пробы были отобраны в следующих фитоценозах: лиственничная редина осоково-кустарничково-сфагновая (т. 19), редкостойный лиственничник багульниково-сфагновый (т. 20–22), лиственничная редина кустарничково-осоково-зеленомошная с подростом ивы (т. 23), редкостойный лиственничник багульниково-зеленомошный (т. 24), лиственничные редины осоково-зеленомошная и голубично-зеленомошная с подлеском различных видов ив (т. 25, 26). Последняя точка отбора (т. 27) характеризует сомкнутый лиственничник грушанковый, расположенный в мезотрофной части болотного массива.
Для растительности болот характерно разнообразие травянисто-кустарничкового яруса, степень проективного покрытия которого достигает 30%. В этом ярусе обильны Vaccinium uliginosum L., Ledum palustre L., Oxycoccus microcarpus Turcz. ex Rupr., Empetrum nigrum, довольно обычны Carex, Eriophorum, Rubus chamaemorus L., Epilobium palustre L. Проективное покрытие мохово-лишайникового яруса составляет 60–100%. Лиственничники зеленомошные (т. 23–26) расположены на ровных участках болот, микрорельеф которых представлен плоскими моховыми буграми и моховыми кочками-подушками, доминируют виды Polytrichum cоmmune, P. strictum L., Pleurozium schreberi. Микрорельеф сфагновых лиственничников (т. 19–22) образован более высокими и большого диаметра буграми и приствольными подушками, доминируют мхи Sphagnum fuscum (Schimp.) H. Klinggr., Sph. angustifolium (C.E.O. Jensen ex Russow) C.E.O. Jensen.
Результаты изучения спорово-пыльцевых спектров поверхностных проб. В спорово-пыльцевых спектрах всех изученных поверхностных проб преобладает пыльца деревьев и кустарников (от 70 до 95%), доминирует пыльца Betula как древесной формы, так и кустарниковой. Весьма высок вклад пыльцы Alnus alnobetula subsp. fruticosa; присутствует также пыльца Larix, Picea, Pinus, Salix и др. (рис. 1).
Рис. 1.
Спорово-пыльцевая диаграмма поверхностных проб растительных сообществ из района пос. Тура (Центральная Эвенкия): 1 – лиственничники сомкнутые, 2 – березняк, 3 – лиственничники разреженные, 4 – лиственничные редины, 5 – пойменная растительность, 6 – болота.
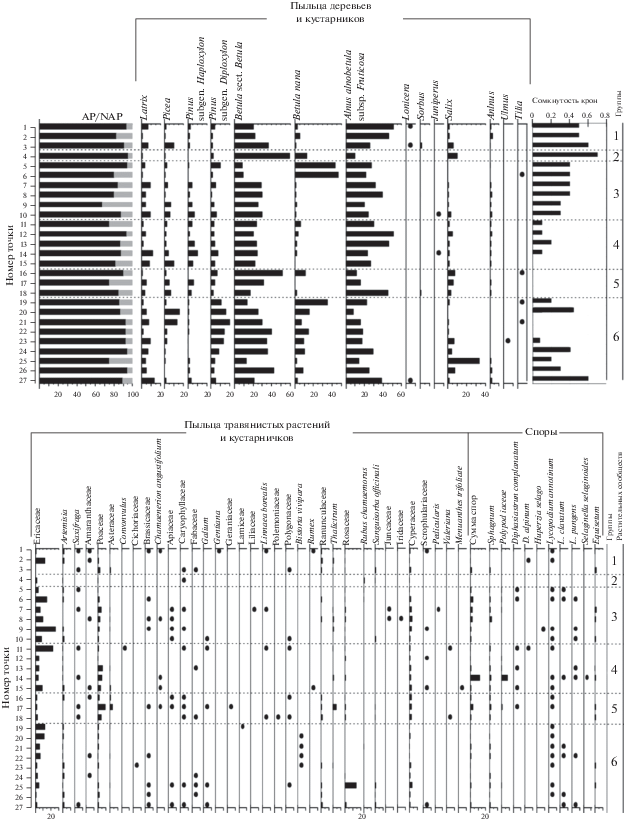
Несмотря на то, что большинство образцов отобрано в различных типах лиственничных лесов, доля пыльцы Larix варьирует от 0.6 до 13.5%, среднее значение 5.2% (рис. 2). Относительно высокие значения участия пыльцы Larix (7–10% и выше) обнаружены как в сомкнутых лиственничниках (т. 1, 3, 27), так и в лиственничных рединах, а также в точке, отобранной на гари, где древостой вообще отсутствует. В то же время в лиственничнике голубично-зеленомошном с сомкнутостью крон лиственницы 0.5 доля пыльцы Larix составляет всего 1.1%. Важно отметить, что значения доли пыльцы лиственницы в большинстве проб (значения между первым и третьим квартилем выборки – рис. 2) находятся в диапазоне от 2 до 7%.
Рис. 2.
Содержание пыльцы основных древесных пород и кустарников в спорово-пыльцевых спектрах поверхностных проб.

Пыльца Picea обнаружена во всех точках отбора проб, за исключением одного местонахождения на болоте, где ее доля колеблется от 0.3 до 15.9%, однако в большинстве проб она находится в диапазоне 0.6–5% при средних значениях 3.5% (см. рис. 2). В точках отбора проб, где ель присутствует в фитоценозе, содержание пыльцы возрастает до 6–9%, в остальных образцах ее доля не превышает 2%. Однако максимальные значения доли пыльцы ели в спектрах (13.1 и 15.9%) отмечены в т. 20 и 21 на болоте. Непосредственно в точках отбора проб ель отсутствует, но, согласно нашим маршрутным наблюдениям, встречается в окрестностях болота.
Во всех точках обнаружена пыльца сосны: доля пыльцы Pinus subgen. Haploxylon составляет от 0.2 до 10.3%, P. subgen. Diploxylon – от 1.2 до 19.6%. Максимальные значения содержания пыльцы обоих видов обнаружены в точках на болотах (т. 19–24), в лиственничных рединах или на открытых пространствах.
В районе исследований произрастают два вида древесных форм берез – Betula pubescens Ehrh. и B. pendula Roth. Поскольку пыльца этих видов морфологически трудно различима, в палинологии принято определять пыльцу древесной березы до секции Betula sect. Betula. Максимальное количество пыльцы древесной формы березы (59.4%) выявлено в спектре березняка хвощевого. В спорово-пыльцевых спектрах других проб ее доля колеблется от 16 до 40%, при этом наиболее высок ее вклад отмечается в тех местообитаниях, где береза присутствует в составе локального фитоценоза.
Участие в спектрах пыльцы Betula nana L. изменяется в широком диапазоне – от 0.8 до 47.3%, но в большинстве точек отбора проб ее доля не превышает 10% (см. рис. 2). Наивысших значений содержание пыльцы Betula nana L. достигает в палиноспектрах из редкостойных лиственничников кустарничково-лишайникового и шикшево-багульниково-сфагнового (т. 5, 6). Высок ее вклад в спектры проб из болотных экотопов (т. 19–24), особенно в тех фитоценозах, где береза карликовая имеет высокое обилие (cop.1, cop.2). В остальных пробах доля пыльцы резко падает.
Alnus alnobetula subsp. fruticosa доминирует в кустарниковом ярусе большинства описанных фитоценозов, и ее пыльца – один из основных компонентов спорово-пыльцевых спектров во всех изученных точках отбора проб. Ее содержание варьирует от 8.2 до 50.1% (пойменный ольховник, т. 18) и в целом выше в образцах, отобранных в местообитаниях, где вид имеет высокое обилие в фитоценозах. Размах значений между первым и третьим квартилями выборки составляет от 18 до 36% (см. рис. 2).
Содержание пыльцы ивы очень сильно зависит от локальных условий и ее участия в растительном сообществе. В спектрах пойменных ивняков и березняке хвощевом доля пыльцы Salix составляет 7–10%. В лиственничной редине осоково-зеленомошной с подлеском из различных видов Salix в пределах болотного массива доля пыльцы возрастает до 33.5%. Пыльца остальных кустарников – Lonicera, Ribes, Sorbus, Juniperus – присутствует единично в нескольких образцах.
В изученных поверхностных пробах обнаружена дальнезаносная пыльца древесных растений. В спектрах четырех точек отбора проб выявлена пыльца Tilia, в одной – пыльца Ulmus и в ряде местообитаний зафиксирована пыльца Alnus sp., принадлежащая видам с древесными жизненными формами, морфологически отличная от Alnus alnobetula subsp. fruticosa.
В группе пыльцы травянистых растений и кустарничков преобладает пыльца сем. Ericaceae, Poaceae, Cyperaceae. Доля пыльцы вересковых составляет от 8 до 24% в точках, где эти кустарнички обильны в фитоценозе, например в лиственничниках багульниково-зеленомошном, кустарничко-лишайниковом, кустарничково-сфагново-зеленомошном. В остальных пробах содержание пыльцы Ericaceae не превышает 2–3%. Пыльца злаков отмечена в количестве 4–5% в пойменных фитоценозах и сухих лиственничниках. В спектрах из ивняка хвощево-вейникового ее вклад составляет 9.4%, и, очевидно, принадлежит видам рода Calamagrostis. В прочих точках участие в спектрах пыльцы злаков колеблется в пределах 1–2%. Пыльца осоковых – постоянный компонент спектров в количестве от 0.8 до 3.2%. В некоторых точках отбора отмечена пыльца Artemisia и Amaranthaceae.
В изученных поверхностных пробах выявлена разнообразная пыльца представителей разнотравья (0.1–0.3%): Apiaceae, Asteraceae, Brassicaceae, Caryophyllaceae, Fabaceae, Geraniaceae, Gentianaceae, Lamiaceae, Polemoniaceae, Scrophulariaceae, Valeriana sp., Thalictrum sp., Bistorta vivipara (L.) Delarbre, Sanguisorba officinalis L. и др. В спектре т. 15 (лиственничная редина осоково-зеленомошная с подлеском из различных видов Salix) содержание пыльцы Rosaceae достигает 14.2%, что, очевидно, обусловлено локальными условиями. В остальных точках ее доля 0.2–1.5% или пыльца не зафиксирована. Пыльца растений нарушенных местообитаний – Convolvulus, Rumex – отмечена единично в спектрах в 1–2 точках, пыльца Chamaenerion angustifolium (L.) Scop. определена в 5 пробах. В пробах выявлена пыльца болотных видов – Rubus chamaemorus, Menyanthes trifoliata L., Pedicularis sp.
Сумма спор составляет от 0.5 до 11% по отношению к сумме AP+NAP. Наиболее часто выявлены споры Sphagnum (0.2–2.2%), папоротников (0.2–7.9%), Lycopodium annotinum L. (0.2–1.5%) и Equisetum (0.2–1.5%). В нескольких точках отмечены споры плаунов Diphasiastrum complanatum (L.) Holub, Lycopodium clavatum L., L. pungens (Desv.) Bach. Pyl. ex Iljin, а споры таких видов, как Huperzia selago (L.) Bernh. ex Schrank & Mart., Selaginella selaginoides (L.) P. Beauv. ex Schrank & Mart., встречены только в одном спорово-пыльцевом спектре. Участие спор сфагновых мхов и хвоща немного выше в тех точках, где они принимают участие в растительных сообществах, в остальных точках они отмечены единично или отсутствуют. Папоротники и плауны непосредственно в местах отбора проб не были зафиксированы, но все эти виды произрастают в районе исследований и могли поступить в спорово-пыльцевой спектр из лесных местообитаний в окрестностях точек отбора.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Полученные нами данные показали, что состав и соотношение компонентов в группе пыльцы деревьев и кустарников требуют тщательного анализа при интерпретации. Доля пыльцы Larix существенно занижена по отношению к ее участию в фитоценозе. Взаимосвязь между количеством пыльцы в спектрах и сомкнутостью крон лиственницы по нашим данным не выявлена. Несоответствие между участием пыльцы Larix в спектрах и ее ролью в растительных сообществах неоднократно отмечали палинологи, изучающие историю лиственничных лесов и изменения растительности в регионах, где они произрастают. Так, в поверхностных пробах из лиственничников Среднего Тимана (Республика Коми) содержание пыльцы Larix варьирует от 0.6 до 5.0% [22]. Здесь и далее процентные соотношения пыльцевых таксонов приводятся от суммы AP+NAP. В том случае, если авторы использовали другие способы расчета, соотношения были пересчитаны. В лиственничных лесах Северного Сахалина доля ее пыльцы может достигать 12.3% при средних значениях 1.5% [12]. Исследования Е.А. Рашке и Л.А. Савельевой [16] в притундровых лиственничных редколесьях в дельте р. Лена показали, что единично пыльца Larix встречается повсеместно, но на площадках, заложенных вблизи взрослых деревьев лиственницы, ее доля возрастает до 4–10%. Среднее значение доли пыльцы Larix в спектрах в лиственничниках в бассейне р. Индигирки – около 3% [17].
Дискуссионным остается вопрос – какую величину доли пыльцы Larix в спектрах можно считать предельной, чтобы сделать заключение о произрастании лиственницы на рассматриваемой территории? Для Европейского Севера О.В. Лисицина с соавт. [32] предлагают 0.5%, в то же время Дж. Клемм с соавт. [17], основываясь на большой выборке поверхностных проб на севере Восточной Сибири, считают эту величину заниженной, предлагая предельное значение 2%. Полученные нами результаты показали, что даже в сомкнутых лиственничных лесах доля пыльцы Larix может составлять 0.6–1.5%. При этом образцы характеризовались очень хорошей сохранностью пыльцы в целом и большой выборкой подсчитанных пыльцевых зерен, что снижает возможность ошибок при анализе.
Picea obovata на изучаемой территории произрастает в виде примеси в лиственничных лесах. Доля ее пыльцы в спектрах невысока, что в целом адекватно отражает участие ели в растительности. Прослеживается четкая тенденция к увеличению участия пыльцы Picea в спектрах в точках отбора проб, где ель входит в состав фитоценоза или произрастает поблизости.
Пыльца сосны (Pinus subgen. Haploxylon и P. subgen. Diploxylon) присутствует в спектрах всех изученных поверхностных проб. Область распространения Pinus sibirica Du Tour охватывает изучаемую территорию [33], однако эта древесная порода находится здесь на границе ареала и, видимо, очень редка. Северная граница ареала сосны обыкновенной Pinus sylvestris L. в среднем течении р. Нижняя Тунгуска проходит примерно на 200 км южнее изучаемой территории [33]. По данным А.С. Прокушкина (устное сообщение), изолированное местонахождение P. sylvestris расположено на расстоянии около 100 км на восток от района исследований (64°05′59.3′′ с.ш., 102°56′33.1′′ в.д.). Следует отметить, что доля пыльцы обеих видов сосен максимальна в точках на болотах, где лесной полог изрежен или отсутствует. Установлено [34], что под пологом леса участие пыльцы, принесенной ветром из отдаленных районов, значительно меньше, чем на открытых местообитаниях, благодаря фильтрующему эффекту крон. Закономерно, что доля пыльцы сосны возрастает в сообществах с очень низкой сомкнутостью крон или вообще лишенных древостоя, где выше доля заносной пыльцы в целом. Все виды сосен обладают высокой пыльцевой продуктивностью, а воздушные мешки позволяют их пыльцевым зернам переноситься по воздуху на значительные расстояния. Некоторые исследователи [18] используют содержание пыльцы Pinus или соотношение Pinus/Larix в качестве индикатора увеличения доли открытых пространств в Арктических районах.
Пыльца Tilia, Ulmus и Alnus также является заносной для изучаемого региона. Границы ареалов этих древесных пород находятся за тысячи километров от района исследований. Перенос пыльцы широколиственных пород и различных видов хвойных в Арктику и регионы с суровым климатом далеко за пределы их произрастания довольно часто фиксировали палинологи, работавшие в этих районах [35–37]. Например, единичные пыльцевые зерна липы и ольхи обнаружены в поверхностных пробах на Полярном Урале [7], на севере Таймыра [37]. Занос пыльцы различных видов сосен – типичное явление для Арктических районов как в Европе [10, 36], так и в Сибири [16, 37, 38].
Данные о соотношении основных компонентов спектров поверхностных проб из лиственничных лесов свидетельствуют о том, что их преобладающие компоненты – это береза и ольха кустарниковая, что закономерно для ветроопыляемых растений с высокой пыльцевой продуктивностью. В большинстве проб содержание пыльцы березы колеблется от 20 до 35%, а ольхи кустарниковой – от 18 до 36% (см. рис. 2). Доля пыльцы березы сильно завышена по отношению к ее участию в древостоях. Подобное явление было описано не только для лиственничных лесов, но и для ельников [8, 9] в лесах, где пыльцевая продуктивность основной лесообразующей породы ниже, чем у березы.
Ольха кустарниковая широко распространена на изучаемой территории, поэтому высокие доли ее пыльцы вполне предсказуемы. Однако прослеживается тенденция к увеличению вклада пыльцы Alnus alnobetula subsp. fruticosa до 45–50%, особенно в ее зарослях. Процентные соотношения пыльцы карликовой березы в целом адекватно описывают ее присутствие в фитоценозе и зависят от ее обилия в локальной растительности. Количество пыльцы ивы также обусловлено особенностями местных условий формирования спорово-пыльцевых спектров.
Состав и соотношение основных компонентов группы пыльцы травянистых растений и кустарничков, а также группы спор отображают основные закономерности видового состава и обилия различных видов в растительном покрове, насколько это позволяет метод спорово-пыльцевого анализа. Детальные исследования состава современной флоры и растительности района исследований показали, что среди кустарничков и травянистых растений лесных сообществ региона основное участие в спорово-пыльцевых спектрах принимают представители семейств Ericaceae, Rosaceae, Poaceae [25]. Пыльца этих семейств наиболее часто отмечена в спорово-пыльцевых спектрах. В болотных местообитаниях увеличивается доля пыльцы Сyperaceae, что обусловлено значительной ролью осок и пушицы в фитоценозах.
Пыльца растений, типичных для сорных местообитаний или участков с нарушенным или несформированным почвенно-растительным покровом, присутствует в большинстве точек. К ним мы отнесли пыльцу Amaranthaceae, Artemisia, Convolvulus, Rumex. Семейство Amaranthaceae представлено в районе исследования только тремя видами – Chenopodium acerifolium Andrz., Ch. rubrum L., Ch. suecicum Murr. [39], растущими по обочинам дорог или берегам рек и озер, на галечниках. Род Artemisia представлен пятью видами, среди которых наиболее часто встречаются Artemisia vulgaris L., растущая по гарям и на опушках лиственничных лесов, и A. laciniata Willd. – лугово-степной вид [40]. Пыльца Chamaenerion angustifolium, присутствующая в пробах, является индикатором гарей, которые часты в районе исследований.
Хотя группа спор немногочисленна, выявлены такие характерные виды таежных лесов, как Lycopodium annotinum, L. clavatum (наиболее часто встречаются в районе исследований), а также Diphasiastrum complanatum и Huperzia selago. Следует отметить находку в одном образце спор Selaginella selaginoides. Плаунок плаунковидный – циркумполярный гипоарктомонтанный вид, очень редко встречающийся в Ангаро-Тунгусской провинции [40], а в районе исследования его местонахождения не были отмечены.
ЗАКЛЮЧЕНИЕ
Анализ состава и соотношения компонентов спорово-пыльцевых спектров поверхностных проб различных типов растительных сообществ в области распространения лиственничных лесов в Центральной Эвенкии позволил сделать следующие выводы:
1. Несмотря на преобладание лиственничных лесов в растительном покрове района исследования, основными компонентами спектров являются Betula sect. Betula и Alnus alnobetula subsp. fruticosa, что объясняется различиями в пыльцевой продуктивности, особенностями распространения и фоссилизации представителей родов Alnus, Betula и Larix. Доля пыльцы Larix занижена по отношению к ее участию в растительности, что было неоднократно отмечено в палинологических работах. Наши исследования показали, что в спорово-пыльцевых спектрах, формирующихся преимущественно за счет поступления пыльцы из воздуха (при отсутствии привноса вещества речными водами или течениями), доля пыльцы Larix может составлять 0.6–1.5% даже в сомкнутых лиственничниках. Доля пыльцы Picea в спектрах невысока (3.5% в среднем) и возрастает в точках отбора проб, где ель входит в состав древостоя.
2. Пыльца Pinus является заносным компонентом спорово-пыльцевых спектров изучаемого региона. Установлено, что доля пыльцы как Pinus subgen. Haploxylon, так и P. subgen. Diploxylon повышается в фитоценозах, где древесный ярус изрежен или отсутствует, что позволяет использовать пыльцу сосны как один из показателей увеличения доли открытых местообитаний на изучаемой территории. Для более точной интерпретации ископаемых спорово-пыльцевых спектров и заключения о присутствии сосны в районе исследования в прошлом необходимы дополнительные доказательства (например, находки семян, хвои, коры, древесины и пр.).
3. Установлено, что на соотношение компонентов палиноспектров группы представителей травянисто-кустарничкового яруса большое влияние оказывают локальные условия. Выявленные характерные виды таежных лесов, болотных и луговых местообитаний, индикаторы нарушенных грунтов и гарей позволят более детально и объективно реконструировать палеорастительность.
Полевые исследования, пробоподготовка, анализ данных выполнены при поддержке проекта РНФ № 20-17-00043, количественный спорово-пыльцевой анализ – из средств федерального бюджета на выполнение государственных заданий Института биологии КарНЦ РАН (АААА-А19-119062590056-0), Института геологии КарНЦ РАН (АААА-А18-118020690231-1).
Авторы подтверждают отсутствие конфликта интересов.
Список литературы
Гричук В.П. Опыт характеристики состава пыльцы в современных отложениях различных природных зон европейской части СССР // Проблемы физической географии. 1941. Вып. 11. С. 101–129.
Гричук В.П., Заклинская Е.Д. Анализ ископаемой пыльцы и спор и его применение в палеогеографии. М.: Географгиз, 1948. 224 с.
Мальгина Е.А. Опыт сопоставления распространения пыльцы некоторых древесных пород с их ареалами в пределах европейской части СССР // Труды ИГ АН СССР. 1950. Т. 46. Вып. 3. С. 256–270.
Федорова Р.В. Количественные закономерности распространения пыльцы древесных пород воздушным путем // Труды ИГ АН СССР. 1952. Т. 52. С. 91–103.
Филимонова Л.В. Поверхностные и приповерхностные спорово-пыльцевые спектры из Среднетаежной подзоны Карелии // Актуальные проблемы палинологии на рубеже третьего тысячелетия: Мат-лы IX всероссийской палинологической конф. М.: Изд-во ИГиРГИ, 1999. С. 311–313.
Короткий А.М. Географические аспекты формирования субфоссильных спорово-пыльцевых комплексов (юг Дальнего Востока). Владивосток: Дальнаука, 2002. 271 с.
Лаптева Е.Г. Субфоссильные спорово-пыльцевые спектры современной растительности Южного Урала // Вестн. Башкирского ун-та. 2013. Т. 18. № 1. С. 77–81.
Новенко Е.Ю., Носова М.Б., Красноруцкая К.В. Особенности поверхностных спорово-пыльцевых спектров южной тайги Восточно-Европейской равнины // Изв. ТулГУ. Естественные науки. 2011. Вып. 2. С. 345–354.
Новенко Е.Ю., Мазей Н.Г., Зерницкая В.П. Рецентные спорово-пыльцевые спектры заповедных территорий европейской части России как ключ к интерпретации результатов палеоэкологических исследований // Nature Conservation Research. Заповедная наука. 2017. № 2(2). С. 55–65. https://doi.org/10.24189/ncr.2017.012
Мазей Н.Г., Кусильман М.В., Новенко Е.Ю. Встречаемость пыльцы Carpinus, Fagus, Tilia и Quercus в субрецентных спорово-пыльцевых спектрах Восточно-Европейской равнины: к вопросу о возможности дальнего заноса пыльцы // Экология. 2018. № 6. C. 431–439.https://doi.org/10.1134/S0367059718050074
Васильчук А.К. Региональная и дальнезаносная пыльца в тундровых палиноспектрах // Известия РАН. Сер. биологич. 2005. № 1. С. 88–99.
Микишин Ю.А., Гвоздева И.Г. Субфоссильные спорово-пыльцевые комплексы Сахалина и прилегающих территорий. Владивосток: Изд-во Дальневост. ун-та, 2009. 162 с.
Рябогина Н.Е., Якимов А.С. Палинологические и палеопочвенные исследования на археологических памятниках: анализ возможностей и методика работ // Вестник археологии, антропологии и этнографии. 2010. № 2. С. 186–200.
Ямских Г.Ю., Макарчук Д.Е. Субфоссильные спорово-пыльцевые спектры – основа реконструкций палинологических данных (на примере территории Красноярской котловины и ее горного обрамления) // Эволюция жизни на Земле: Мат-лы V межд. симпозиума. Томск, 2018. С. 229–230.
Ложкин А.В. Современный пыльцевой дождь в арктических районах Берингии и реконструкция растительности ледниковых интервалов плейстоцена // Четвертичная палеогеография Берингии. Магадан: СВКНИИ ДВО РАН, 2002. С. 13–27.
Рашке Е.А., Савельева Л.А. Субрецентные спорово-пыльцевые спектры и современная растительность дельты реки Лена, Российская Арктика // Сибирс-кий экологич. журн. 2017. Т. 24. № 4. С. 456–472. https://doi.org/10.15372/SEJ20170407
Klemm J., Herzschuh U., Pisaric M.F.J. et al. A pollen-climate transfer function from the tundra and taiga vegetation in Arctic Siberia and its applicability to a Holocene record // Palaeogeography, Palaeoclimatology, Palaeoecology. 2013. V. 386. P. 702–713. https://doi.org/10.1016/j.palaeo.2013.06.033
Klemm J., Herzschuh U., Pestryakova L.A. Vegetation, climate and lake changes over the last 7000 years at the boreal treeline in north-central Siberia // Quaternary Science Reviews. 2016. V. 147. P. 422–434. https://doi.org/10.1016/j.quascirev.2015.08.015
Эктова С.Н., Лаптева Е.Г., Трофимова С.С. Отражение флористического состава тундровой растительности долины р. Юрибей (Средний Ямал) в рецентных комплексах растительных остатков // Проблемы региональной экологии. 2013. № 4. С. 39–44.
Лопатина Д.А., Занина О.Г. Субрецентные спорово-пыльцевые спектры низовьев р. Колыма и их значение для реконструкции четвертичной палеогеографии региона // Стратиграфия. Геологическая корреляция. 2016. Т. 24. № 2. С. 203–211. https://doi.org/10.7868/S0869592X16020034
Палеоклиматы и палеоландшафты внетропического пространства Северной Евразии. Поздний плейстоцен–голоцен. Атлас-монография / Ред. Величко А. А. М.: ГЕОС, 2009. 120 с.
Непомилуева Н.И., Дурягина Д.А. К истории лиственничников среднего Тимана в голоцене (Коми АССР) // Ботанич. журн. 1990. №3. Т. 75. С. 326–335.
URL: https://climexp.knmi.nl/start.cgi (дата обращения 20.08.2020).
Коротков И.А. Лесорастительное районирование России и республик бывшего СССР // Углерод в экосистемах лесов и болот России / Ред. Алексеева В.А., Бердси Р.А. Красноярск: Изд-во ИЛ СО РАН, 1994. С. 29–47.
Кривобоков Л.В., Зверев А.А. Классификация растительности и особенности ценофлоры лиственничных лесов криолитозоны Средней Сибири (бассейн р. Нижняя Тунгуска) // Проблемы ботаники Южной Сибири и Монголии. 2015. № 14. С. 99–104.
Харук В.И., Двинская М.Л., Рэнсон К.Дж. Пространственно-временная динамика пожаров в лиственничных лесах северной тайги Средней Сибири // Экология. 2005. № 5. С. 334–343.
Knorre A.A., Kirdyanov A.V., Prokushkin A.S. et al. Tree ring-based reconstruction of the long-term influence of wildfires on permafrost active layer dynamics in Central Siberia // Science of the Total Environment. 2019. V. 652. P. 314–319. https://doi.org/10.1016/j.scitotenv.2018.10.124
Полевая геоботаника. Л.: Наука, 1976. Т. 5. 320 с.
Moore P.D., Webb J.A., Collinson M.E. Pollen Analysis. Oxford, Boston: Blackwell Scientific Publications, 1991. 216 p.
Grimm E.C. TILIA and TILIA GRAPH.PC spreadsheet and graphics software for pollen data // INQUA. Working Group on Data-Handling Methods. Newsletter. 1990. № 4. P. 5–7.
Карпенко Л.В. Растительный покров и оценка видового разнообразия заболоченных лиственничников и болот Центральной Эвенкии // Хвойные бореальной зоны. 2013. № 5–6. С. 22–28.
Lisitsyna O.V., Giesecke T., Hicks S. Exploring pollen percentage threshold values as an indication for the regional presence of major European trees // Review of Palaeobotany and Palynology. 2011. V. 166. P. 311–324. https://doi.org/10.1016/j.revpalbo.2011.06.004
Ареалы деревьев и кустарников СССР. Т. 1. Л.: Наука, 1977. 164 с.
Dimbelby G.W. Soil pollen analysis // Soil Science. 1961. V.12. № 1. P. 3–11.
Малясова Е.С. Методическая основа формирования спорово-пыльцевых спектров в донных осадках северных морей // Четвертичная палеоэкология и палеогеография северных морей / Ред. Матишов Г.Г., Тарасова Г.А. М.: Наука, 1988. С.162–169.
Руденко О.В. Изменения природной среды Восточно-Баренцевоморского региона в голоцене по материалам изучения субфоссильных и ископаемых пыльцевых спектров Новоземельского шельфа // Проблемы Арктики и Антарктики. 2011. № 3 (89). С. 33–42.
Andreev A.A., Tarasov P.E., Klimanov V.A. et al. Vegetation and climate changes around the Lama Lake, Taymyr Peninsula, Russia during the Late Pleistocene and Holocene // Quaternary International. 2004. № 122. P. 69–84. https://doi.org/10.1016/j.quaint.2004.01.032
Andreev A.A., Tarasov P.E., Siegert C. et al. Late Pleistocene and Holocene vegetation and climate on the northern Taymyr Peninsula, Arctic Russia // Boreas. 2003. № 32. P. 484–505. https://doi.org/10.1006/qres.2001.230
Флора Сибири: В 14 т. Т. 5: Salicaceae – Amaranthaceae / Ред. Красноборова И.М., Малышева Л.И. Новосибирск: Наука, 1992. 312 с.
Флора Красноярского края. Вып. 1 / Ред. Положиго А. В. Томск: Изд-во Томск. ун-та, 1983. 83 с.
Дополнительные материалы отсутствуют.