

Экология, 2021, № 6, стр. 412-419
Экологическая характеристика Anemonastrum biarmiense (Juz.) Holub на Южном Урале
П. С. Широких a, *, О. В. Юсупова b, c, Л. М. Абрамова d, Л. Г. Наумова e
a Уфимский институт биологии Уфимского федерального исследовательского центра РАН
450054 Уфа, просп. Октября, 69, Россия
b Южно-Уральский государственный природный заповедник
453560 Реветь, ул. Заповедная, 1, Россия
c Национальный парк “Зюраткуль”
456910 Сатка, ул. Комсомольская, 13, Россия
d Южно-Уральский ботанический сад-институт – обособленное структурное подразделение Уфимского федерального исследовательского центра РАН
450080 Уфа, ул. Менделеева, 195/3, Россия
e Башкирский государственный педагогический университет им. М. Акмуллы
450000 Уфа, ул. Октябрьской революции, 3а, Россия
* E-mail: shirpa@mail.ru
Поступила в редакцию 01.02.2021
После доработки 14.05.2021
Принята к публикации 17.05.2021
Аннотация
С использованием эколого-флористического подхода и экологических шкал Д.Н. Цыганова проведен анализ влияния абиотических факторов среды на распространение эндемика Южного Урала – Аnemonastrum biarmiense (Juz.) Holub. Установлено, что по группе почвенных факторов A. biarmiense имеет широкую амплитуду и центрированное положение синэкологического оптимума вида. Поэтому изменение эдафических условий существенно не влияет на его продуктивность и виталитет. По группе климатических факторов А. biarmiense характеризуется узким диапазоном реализованной экологической ниши. Синэкологический оптимум смещен к границам реализованной ниши и соответствует горно-тундровых сообществам, где экологические условия местообитаний меняются на относительно коротких расстояниях. Поэтому реакция A. biarmiense на потепление климата будет связана в первую очередь со снижением плотности ценопопуляций, а впоследствии – с сокращением его местообитаний. Таким образом, сочетание узкой амплитуды толерантности и расположение синэкологического оптимума на границах реализованной ниши делает A. biarmiense хорошим индикатором глобального изменения климата.
Сохранение биоразнообразия растений является одним из приоритетных направлений современной ботаники. Особое внимание уделяется редким и эндемичным видам, как наиболее уязвимым компонентам растительного покрова в результате антропогенного воздействия. К таковым относится Anemonastrum biarmiense (Juz.) Holub – эндемичный вид Урала из семейства Ranunculaceae Juss. По происхождению он принадлежит к Уральской викарной расе полиморфных видовых комплексов, широко распространенных в высокогорьях различных горных систем Северной Евразии и Голарктики в целом, возникших в результате дифференциации и раздробления ареала общего предкового вида [1, 2].
На Южном Урале A. biarmiense встречается по горным хребтам с отметками от 1000 до 1640 м над ур. м., а также места его произрастания отмечаются в горно-лесном поясе, ниже 1000 м над ур. м. Вид включен в список таксонов Российской Федерации, нуждающихся в особом внимании к их состоянию в природной среде и мониторинге [3]. С третьей категорией редкости вид включен в Красные книги Свердловской области [4], Республики Коми [5], Ханты-Мансийского [6] и Ямало-Ненецкого [7] автономных округов.
Важное значение для видов, требующих особого внимания, имеет оценка влияния комплекса экологических факторов на изучаемые популяции. К наиболее значимым экологическим факторам, определяющим жизнедеятельность растений, относят климатические (солнечная радиация, температура, осадки и т.д.) и факторы местообитания (физический и химический состав почвы, ее увлажненность). Расчет их воздействия на конкретные популяции растений позволяет провести количественную оценку экологической толерантности видов, анализ их экологических предпочтений, рассмотреть экологические механизмы устойчивости [8–13] в тех случаях, когда прямое измерение параметров чрезвычайно сложно или невозможно.
Цель данной работы – анализ абиотических факторов среды, влияющих на распространение эндемичного вида Anemonastrum biarmiense, и оценка его устойчивости в связи с глобальным потеплением климата.
МАТЕРИАЛ И МЕТОДЫ
Район, где проводили исследования, относится к центрально-возвышенной части Южного Урала, расположенной на территории Белорецкого района Республики Башкортостан (РБ) и Катав-Ивановского района Челябинской области. Центрально-возвышенная часть является наиболее высоким и орографически сложным районом Южного Урала, в котором отчетливо выражена высотная поясность в распределении климатических условий и зависящих от них почв, растительности, а также ландшафтов в целом. Достаточно отчетливо выражены 3 пояса растительности – горно-лесной, подгольцовый и горно-тундровый [14].
По термическим условиям климат района характеризуется как умеренно холодный: среднегодовая температура воздуха составляет от +0.6 до +1.5°С, среднеиюльская +16…+17°С, среднеянварская –15…–16°С. Минимальная температура опускается до –41°С (абсолютный минимум в 1940 г. достиг –49°С). За год выпадает 600–800 мм осадков. Годовое испарение составляет от 480 до 500 мм [15].
С увеличением высоты изменяются все климатические характеристики: повышается радиационный баланс, понижается летняя температура, укорачиваются вегетационный и безморозный периоды, увеличиваются суммы осадков. В районе массива горы Большой Ямантау среднеиюльская температура понижается с 16 (на высоте 500 м) до 9°С (на высоте 1600 м над ур. м.), сумма активных температур – с 1600 до 800–400°С, безморозный период уменьшается с 90 до 60 дней, годовая сумма осадков увеличивается с 700 до 1100 мм [16].
Анализ фитоценотической приуроченности Anemonastrum biarmiense проводился в рамках разработанной классификации Южно-Уральского региона – ЮУР [17] с учетом последней сводки высших синтаксономических единиц [18]. В обработку были включены синтаксоны с участием изучаемого вида из опубликованных источников [19–21], базы данных УИБ УФИЦ РАН [22] и 67 описаний, выполненных О.В. Юсуповой в пределах Южно-Уральского заповедника, национальных парков “Зюраткуль” и “Таганай”, природного парка “Иремель” и на прилегающих к ним территориях. Общий массив данных составил 430 геоботанических описаний. Описания площадок выполняли по методике Браун-Бланке [23]. Для обработки материалов использовали базу данных TURBOVEG [24] и программу JUICE [25].
Для оценки экологических параметров местообитаний были применены диапазонные экологические шкалы Д.Н. Цыганова [8], адаптированные для территории ЮУР [26]. Подсчет значений экологических переменных проводили в интегрированной геоботанической системе IBIS [27] с использованием метода взвешенного усреднения [28]. Для количественной оценки каждого фактора использовали понятие “экологическая валентность”, а для комплекса факторов – “толерантность” видов [29].
Приспособленность популяции конкретного вида к изменению экологических факторов оценивали показателями потенциальной (PEV) и реализованной (REV) экологической валентности видов, эффективность освоения экологического пространства ценопопуляций Anemonastrum biarmiense – при помощи коэффициента экологической эффективности (Kec. eff., %), меру стено-эврибионтности – при помощи индекса толерантности (It). Подробная методика вычисления этих показателей была описана ранее [11, 29, 30].
Ординация сообществ в экологическом пространстве выполнена в программе CANOCO [31].
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Анализ абиотических факторов затруднен из-за их неразрывной связи, поэтому в каждом конкретном случае сложно определить относительную роль отдельного экологического фактора, не вырывая его из общей совокупности условий местообитания. С учетом этого анализ и интерпретацию результатов логичнее проводить по двум группам: климатическим и почвенным, с акцентом на отдельные наиболее важные факторы.
На основе рассчитанных данных (табл. 1) были построены лепестковые диаграммы (рис. 1), отражающие положение A. biarmiense в экологическом пространстве и являющиеся частью общего геометрического образа аутэкологического и синэкологического ареалов. Эти понятия аналогичны представлениям о фундаментальной и реализованной экологической нише Хатчинсона [32–34].
Таблица 1.
Экологическая валентность и коэффициенты экологической эффективности Anemonastrum biarmiense по шкалам Д.Н. Цыганова [8]
| Класс растительности | Экологический факторы | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| климатические | почвенные | Lc | |||||||||
| Tm | Kn | Om | Cr | Hd | Tr | Rc | Nt | Fh | |||
| PEV | 0.41 | 0.60 | 0.20 | 0.27 | 0.30 | 0.47 | 0.68 | 0.38 | 0.48 | 0.56 | |
| Juncetea trifidi | REV | 0.16 | 0.07 | 0.06 | 0.12 | 0.11 | 0.10 | 0.21 | 0.10 | 0.14 | 0.15 |
| Kec. eff. | 0.39 | 0.12 | 0.31 | 0.46 | 0.37 | 0.21 | 0.33 | 0.26 | 0.32 | 0.26 | |
| Loiseleurio-Vaccinietea | REV | 0.10 | 0.07 | 0.05 | 0.07 | 0.05 | 0.08 | 0.13 | 0.09 | 0.10 | 0.09 |
| Kec. eff. | 0.25 | 0.12 | 0.24 | 0.28 | 0.17 | 0.16 | 0.21 | 0.23 | 0.22 | 0.17 | |
| Mulgedio-Aconitetea | REV | 0.09 | 0.04 | 0.04 | 0.07 | 0.05 | 0.07 | 0.12 | 0.12 | 0.09 | 0.09 |
| Kec. eff. | 0.23 | 0.07 | 0.20 | 0.26 | 0.17 | 0.15 | 0.19 | 0.31 | 0.19 | 0.16 | |
| Molinio-Arrhenatheretea | REV | 0.07 | 0.06 | 0.06 | 0.06 | 0.11 | 0.08 | 0.16 | 0.07 | 0.11 | 0.14 |
| Kec. eff. | 0.17 | 0.09 | 0.30 | 0.22 | 0.36 | 0.17 | 0.25 | 0.17 | 0.24 | 0.26 | |
| Vaccinio-Piceetea | REV | 0.07 | 0.02 | 0.04 | 0.07 | 0.05 | 0.05 | 0.12 | 0.06 | 0.08 | 0.09 |
| Kec. eff. | 0.16 | 0.03 | 0.19 | 0.25 | 0.18 | 0.11 | 0.19 | 0.15 | 0.18 | 0.17 | |
| Brachypodio-Betuletea | REV | 0.05 | 0.03 | 0.03 | 0.05 | 0.06 | 0.05 | 0.11 | 0.05 | 0.07 | 0.08 |
| Kec. eff. | 0.11 | 0.04 | 0.17 | 0.21 | 0.19 | 0.11 | 0.17 | 0.14 | 0.17 | 0.15 | |
| Asaro-Abietetea | REV | 0.02 | 0.01 | 0.01 | 0.02 | 0.01 | 0.02 | 0.03 | 0.03 | 0.03 | 0.03 |
| Kec. eff. | 0.05 | 0.02 | 0.07 | 0.08 | 0.03 | 0.03 | 0.06 | 0.09 | 0.07 | 0.05 | |
| Обобщенные данные по всем классам растительности | |||||||||||
| REV | 0.19 | 0.14 | 0.12 | 0.14 | 0.24 | 0.16 | 0.37 | 0.16 | 0.22 | 0.21 | |
| Kec. eff. | 0.45 | 0.24 | 0.60 | 0.53 | 0.79 | 0.43 | 0.82 | 0.42 | 0.30 | 0.37 | |
| It | Itклим = 0.37 | Itпочв = 0.46 | |||||||||
| Экологическая позиция вида | |||||||||||
| Потенциальная | 2–8 | 7–15 | 7–9 | 4–7 | 9–15 | 1–7 | 6–10 | 3–7 | 3–7 | 1–5 | |
| Реальная | 3.8–7.8 | 8.0–9.7 | 7.7–9.1 | 5.0–7.0 | 10.4–14.7 | 4.0–6.8 | 4.6–7.8 | 3.9–5.9 | 4.0–6.1 | 3.4–4.8 | |
| Оптимум вида | 5.1 | 8.9 | 8.6 | 5.1 | 12.6 | 5.1 | 5.9 | 4.9 | 4.9 | 4.1 | |
Примечание. REV – реальная экологическая валентность, PEV – потенциальная экологическая валентность, It – индекс толерантности, Kec. eff. – коэффициент экологической эффективности. Фракции валентности и группы толерантности видов: стеновалентная и стенобионтная – показатель валентности, или индекс толерантности <0.33; гемистеновалентная и темистенобионтная – от 0.34 до 0.45; мезовалентная и мезобионтная – от 0.46 до 0.56; гемиэвривалентная и гемиэврибионтная – от 0.57 до 0.66; эвривалентная и эврибионтная – от 0.67 и выше [29]. Обозначения экологических факторов соответствуют рис. 1.
Рис. 1.
Графическая модель фрагмента экологической ниши Anemonastrum biarmiense на Южном Урале по группе почвенных факторов (а) и по группе климатических факторов, включая освещенность (б): 1 – диапазон (min–max) потенциальной экологической ниши, 2 – диапазон (min–max) реализованной экологической ниши, 3 – оптимум вида. Экологические факторы: Tm – термоклиматический, Kn – континентальность климата, Om – аридность-гумидность, Cr – суровость зимнего периода, Hd – увлажнение, Tr – солевой режим почв, Rc – кислотность почв, fH – переменность увлажнения, Nt – богатство почв азотом, Lc – освещенность (далее в табл. 1, 3 и на рис. 2 для экологических факторов используются те же обозначения). Цифрами на оси отображены значения экологических факторов.
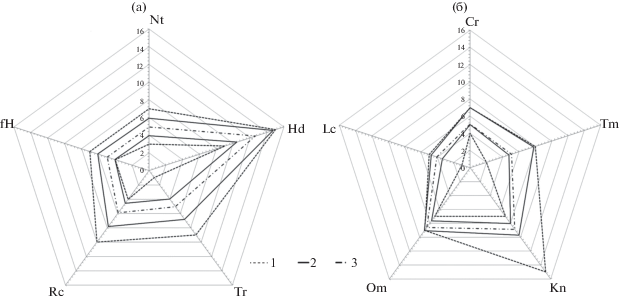
По группе эдафических факторов ветренник пермский имеет довольно широкую амплитуду изменений реализованной ниши (см. рис. 1а), произрастает в различных почвенных условиях (от каменистых и бедных азотом почв до хорошо развитых, богатых элементами минерального питания лесных и луговых почв). Наибольшая эффективность освоения экологического пространства выявлена по факторам увлажнения и степени кислотности почв (см. табл. 1), диапазон которых варьирует от кислых слаболесолуговых до почти нейтральных сыролесолуговых почв.
По группе климатических факторов вид имеет более узкий экологический диапазон (см. рис. 1б). Верхний предел реализованной экологической ниши, за исключением фактора континентальности (Kn), практически полностью совпадает с границами максимальной потенциальной экологической ниши, а нижний предел приближается к ее минимальным пороговым значениям, что свидетельствует о более высокой эффективности освоения экологического пространства по сравнению с группой эдафических факторов. По степени толерантности к климатическим факторам ветренник пермский больше тяготеет к стенобионтам (см. табл. 1).
При оценке устойчивости вида к абиотическим факторам среды важно также учитывать не только пределы толерантности, но и положение оптимума вида (аутэкологического и синэкологического) [35]. По группе почвенных факторов синэкологический оптимум A. biarmiense занимает центрированное положение в реализованной экологической нише (см. рис. 1а). Учитывая довольно широкую амплитуду толерантности, смещение синэкологического оптимума при изменении эдафических условий существенно не повлияет на продуктивность и виталитет вида. В группе климатических факторов синэкологический оптимум вида смещен к нижней границе реализованной ниши, которая отражает наиболее холодный и суровый климат, в особенности по термоклиматическому (Tm) и криоклиматическому (Cr) факторам. Именно в этих сообществах A. biarmiense имеет максимально высокие обилие (см. табл. 2) и продуктивность [38, 39].
Таблица 2.
Встречаемость Anemonastrum biarmiense в сообществах Южного Урала
| № п.п. |
Класс | Союз | Число ассоциаций/ сообществ | Обилие вида, % | Частота встречае- мости, % |
Число описаний |
|---|---|---|---|---|---|---|
| Горно-тундровый пояс | ||||||
| 1 | Juncetea trifidi | Lagotido uralensis-Caricion ensifoliae | 4/6 | 5–15 | 91 | 95 |
| 2 | Loiseleurio procumbentis-Vaccinietea | Loiseleurio procumbentis-Vaccinion | 2/3 | <1 | 63 | 35 |
| 3 | Juniperion nanae | 1 | <1 | 30 | 15 | |
| Горно-тундровый и подгольцовый пояса | ||||||
| 4 | Mulgedio-Aconitetea | Calamagrostion villosae | 2/2 | 1–5 | 84 | 37 |
| 5 | Calamagrostion arundinaceae | 2/– | <1 | 43 | 30 | |
| Горно-лесной пояс (высокогорье) | ||||||
| 6 | Molinio-Arrhenatheretea | Polygonion krascheninnikovii | 1/5 | 1–5 | 92 | 50 |
| 7 | Vaccinio-Piceetea | Brachypodio pinnatae-Pinion sylvestris | 1/– | <1 | 100 | 33 |
| 8 | Piceion excelsae | 2/– | <1 | 95 | 42 | |
| Горно-лесной пояс (среднегорье и низкогорье) | ||||||
| 9 | Brachypodio pinnati-Betuletea pendulae | Veronico teucrii-Pinion sylvestris | 2/– | <1 | 20 | 35 |
| 10 | Trollio europaea-Pinion sylvestris | 2/– | <1 | 22 | 28 | |
| 11 | Asaro europaei- Abietetea sibiricae | Aconito septentrionalis-Piceion obovatae | 1/– | <1 | 20 | 30 |
Подобное смещение оптимумов, как и амплитуды, часто встречается у разных видов растений. Сильные конкуренты и виды, значительно изменяющие среду, могут вытеснять более слабые из благоприятных для них условий местообитания, что приводит к сужению реализованной экологической ниши и смещению оптимумов. Классическим примером является межвидовое взаимодействие сосны (Pinus sylvestris), которая в культуре лучше растет на средних по увлажнению почвах, в то время как в природе распространена преимущественно на сухих (пески, скалы) либо переувлажненных (болота) почвах. Поэтому сосна имеет два синэкологических оптимума, что обычно объясняется конкурентным вытеснением ее елью со средних по увлажнению участков [35]. Большая часть видов рода Ranunculus по ряду экологических факторов также имеют значительную степень асимметрии оптимума по отношению к реализованной экологической нише [36, 37].
Взаимодействие A. biarmiense с другими видами в данной статье мы не рассматриваем, но совершенно очевиден факт взаимного влияния климатических и биотических факторов на его распространение. Данное явление подтверждает ординация сообществ с участием A. biarmiense (рис. 2, табл. 2), отражающая динамику биологической продуктивности (обилия, частоты встречаемости) и смены фитоценозов в зависимости от изменений абиотических факторов среды на градиенте высотной поясности. Ведущими факторами в распространении вида являются климатические показатели, коэффициенты корреляции которых превышают 90% (табл. 3). В более “холодных” климатических условиях распространены сообщества классов Loiseleurio-Vaccinietea и Juncetea trifidi, объединяющие растительность горных тундр и травяно-моховых низкотравных тундровых лугов на кислых почвах субальпийского пояса и бореально-неморальных высокогорий. Сообщества встречаются на хребтах Зигальга, Машак, Нары, Нургуш, Кумардак и на массиве Ямантау, в местообитаниях с суровым морозным климатом, неразвитой кислой почвой и резко переменным режимом увлажнения. В этих сообществах A. biarmiense встречается с высоким постоянством и достигает максимальной плотности в тундровых сообществах союза Lagotido uralensis-Caricion ensifoliae (проективное покрытие вида 5–15%).
Рис. 2.
Ординация сообществ с участием Anemonastrum biarmiense до уровня союза/подсоюза. Класс Loiseleurio-Vaccinietea: 1 – союз Loiseleurio-Vaccinion; класс Juncetea trifidi: 2 – союз Lagotido uralensis-Caricion ensifoliae; класс Mulgedio-Aconitetea: 3 – союз Calamagrostion villosae, 4 – союз Calamagrostion arundinaceae; класс Molinio-Arrhenatheretea: 5 – союз Polygonion krascheninnikovii; класс Vaccinio-Piceetea: 6 – союз Brachypodio pinnatae-Pinion sylvestris, 7, 8 – союз Piceion excelsae (7 – подсоюз Eu-Piceenion abietis, 8 – подсоюз Atrageno sibiricae-Piceenion obovatae); класс Brachypodio-Betuletea: 9 – союз Veronico teucrii-Pinion sylvestris, 10 – союз Trollio europaea-Pinion sylvestris; класс Asaro-Abietetea: 11 – союз Aconito septentrionalis-Piceion obovatae.

С понижением вертикальной поясности повышается общий температурный режим, горные тундры сменяются “тундроподобными” сообществами, высокогорными лугами и редколесьями классов Molinio-Arrhenatheretea и Mulgedio-Aconitetea. В данных сообществах A. biarmiense встречается с высокой частотой, но очень низким обилием. В горно-лесном поясе эти сообщества замещаются бореальными лесами класса Vaccinio-Piceetea. С понижением высоты над уровнем моря на западном макросклоне Южного Урала они замещаются гемибореальными травяными лесами класса Brachypodio-Betuletea, а на восточном – темнохвойными субнеморальными лесами класса Asaro-Abietetea. В этих сообществах A. biarmiense встречается единично.
На градиенте снижения высотной поясности и повышения температурного режима эффективность освоения экологического пространства также уменьшается (см. табл. 1). Наиболее эффективно экологическое пространство осваивается видом в горно-тундровых сообществах, высокогорных лугах и редколесьях классов Juncetea trifidi, Loiseleurio-Vaccinietea, Mulgedio-Aconitetea. В этих сообществах по ряду климатических (Tm, Om, Cr) и почвенных (Hd, Rc, Fh) факторов получены наибольшие значения коэффициента, которые уменьшаются от горно-тундровых сообществ до лесных фитоценозов низкогорий Южного Урала.
Предполагается, что реакция видов на изменение климата будет активной в экотонах и пограничных экосистемах, где растительные формы жизни и почвенные условия быстро изменяются на относительно коротких расстояниях. Таковыми являются сообщества горно-тундрового пояса. Начало этих изменений уже фиксируется в региональном [40] и глобальном [41–45] масштабах. В частности, реакция A. biarmiense на потепление климата будет связана со снижением численности и плотности популяций, а затем и с сокращением местообитаний. Следовательно, оценивая состояние таких видов с узким экологическим диапазоном, как A. biarmiense, можно достаточно эффективно отслеживать климатогенные изменения.
ЗАКЛЮЧЕНИЕ
Согласно полученным данным, распределение Anemonastrum biarmiense в экологическом пространстве связано с комплексным воздействием климатических и почвенных факторов, обусловленного изменением положения сообществ с его участием в высотно-поясном ряду. В фитоценотическом отношении распространение вида охватывает довольно широкий диапазон – от криофильных горно-тундровых сообществ до темнохвойных субнеморальных и ксерофитных травяных светлохвойных гемибореальных лесов. Лимитирующими является группа климатических факторов. Синэкологический оптимум вида расположен в пограничных экосистемах – горно-тундровых сообществах, большая часть которых находится на территории ООПТ различного уровня, что вызывает оптимизм в отношении сохранности этого вида.
Тем не менее прогнозируемые в будущем климатические изменения в сторону потепления и уменьшения количества годовых осадков могут привести к опасным для существования вида и его местообитаний последствиям. В первую очередь будет происходить зарастание лесом горно-тундровых сообществ (основных местообитаний вида) с последующим замещением на бореальные темнохвойные леса, которые в долгосрочной перспективе сменятся травяными гемибореальными и субнеморальными хвойными лесами. Данная сукцессия приведет к исчезновению местообитаний эндемичного вида A. biarmiense, а в глобальном масштабе – к снижению разнообразия на видовом и ценотическом уровнях.
В совокупности все климатические факторы находятся в тесной многосторонней взаимосвязи, отражающие состояния сложной термодинамической системы – атмосферы. Изменение одного показателя приведет к сдвигу других, что повлечет за собой снижение численности ветренника пермского и уменьшению местообитаний, возможных для его обитания. Узкий диапазон реализованной экологической ниши A. biarmiense и наличие нескольких лимитирующих факторов делает его хорошим индикатором глобального изменения климата.
Работа выполнена в рамках государственного задания ЮУБСИ УФИЦ РАН по теме АААА-А18-118011990151-7 и УИБ УФИЦ РАН № 075-00326-19-00 по теме № АААА-А18-118022190060-6.
Список литературы
Мамаев С.А., Князев М.С. Ветреницы в природе и культуре. Екатеринбург: Изд-во УрО РАН, 1995. 18 с.
Куликов П.В., Золотарева Н.В., Подгаевская Е.Н. Эндемичные растения Урала во флоре Свердловской области. Екатеринбург: Гощицкий, 2013. 612 с.
Красная книга Российской Федерации (растения и грибы) / Гл. редколл.: Трутнев Ю.П. и др.; Сост. Камелин Р.В. и др. М.: Тов-во научных изд. КМК, 2008. 855 с.
Красная книга Свердловской области: животные, растения, грибы. Екатеринбург, 2018. 450 с.
Красная книга Республики Коми. Сыктывкар, 2019. 768 с.
Красная книга Ханты-Мансийского автономного округа – Югры: животные, растения, грибы. Екатеринбург, 2013. 460 с.
Красная книга Ямало-Ненецкого автономного округа: животные, растения, грибы. Екатеринбург, 2010. 308 с.
Цыганов Д.Н. Фитоиндикация экологических режимов в подзоне хвойно-широколиственных лесов. М.: Наука, 1983. 196 с.
Булохов А.Д. Фитоиндикация и ее практическое применение. Брянск: Изд-во БГУ, 2004. 245 с.
Shirokikh P.S., Martynenko V.B. Comparison of different ecological scales with respect to efficiency in assessing ecological conditions in forests of the Southern Ural Region // Russ. J. of Ecology. 2009. V. 40. № 7. P. 457–465.
Жукова Л.А., Дорогова Ю.А., Турмухаметова Н.В. и др. Экологические шкалы и методы анализа экологического разнообразия растений. Йошкар-Ола: Мар. гос. ун-т, 2010. 368 с.
Зубкова Е.В. О некоторых особенностях диапазонных экологических шкал растений Д.Н. Цыганова // Изв. Самарского научного центра РАН. 2011. Т. 13. № 5. С. 48–53.
Золотова Е.С., Иванова Н.С. Использование шкал Д.Н. Цыганова для анализа экологического пространства типов леса Среднего Урала // Фундаментальные исследования. 2015. № 2. С. 5114–5119.
Горчаковский П.Л. Растительный мир высокогорного Урала. М.: Наука, 1975. 284 с.
Физико-географическое районирование Башкирской АССР / Под ред. Кадильникова И.П. и др. Уфа: Башкирский гос. ун-т, 1964. 210 с.
Справочник по климату СССР. Вып. 9. Ч. II–IV. Л.: Гидрометеоиздат, 1965. 362 с.
Ямалов С.М. Продромус растительных сообществ Республики Башкортостан. Уфа: Гилем, 2012. 100 с.
Mucina L., Bültmann H., Dierßen K. et al. Vegetation of Europe: hierarchical floristic classification system of vascular plant, bryophyte, lichen, and algal communities // Appl. Veg. Sci. 2016. V.19. P. 3–264. https://doi.org/10.1111/avsc.12257
Ишбирдин А.Р., Муллагулов Р.Ю., Янтурин С.И. Растительность горного массива Иремель: синтаксономия и вопросы охраны. Уфа, 1996. 109 с.
Флора и растительность Южно-Уральского государственного природного заповедника / Под ред. Миркина Б.М. Уфа: Гилем, 2008. 528 с.
Заугольнова Л.Б., Мартыненко В.Б. Определитель типов леса eвропейской части России. 2012. [Электронный ресурс] URL: http://mfd.cepl.rssi.ru/forest/index.htm
Martynenko V., Shirokikh P., Solomeshch A., Muldashev A. Vegetation database forest of Southern Ural // Biodiversity & Ecology. 2012. V. 4. P. 289.
Braun-Blanquet J. Pflanzensoziologie. Grundzüge der Vegetationskunde. 3 Aufl. Wien; New York. 1964. 865 p. https://doi.org/10.1007/978-3-7091-8110-2
Hennekens S.M. TURBO(VEG). Software package for input, processing and presentation of phytosociological data. Lancaster: Wageningen et University of Lancaster, 1995. 70 p.
Tichý L., Holt J., Nejezchlebová M. JUICE. Program for management, analysis and classification of ecological data. 2nd edition. Brno: Masaryk University, 2011. 61 p.
Широких П.С., Зверев А.А. Некоторые результаты коррекции экологических шкал в системе Д.Н. Цыганова для Южно-Уральского региона // Актуальные проблемы геоботаники. Уфа: Издательский центр “МедиаПринт”, 2012. С. 551–556.
Зверев А.А. Информационные технологии в исследованиях растительного покрова: Учеб. пособ. Томск: Изд-во “ТМЛ-Пресс”, 2007. 304 с.
Зверев А.А. Методические аспекты применения фитоиндикационного анализа в изучении биоразнообразия // Сибирский экологич. журн. 2020. Т. 27. № 4. С. 401–415.
Дорогова Ю.А., Жукова Л.А. Экологическая характеристика ценопопуляций липы сердцевидной в подзоне хвойно-широколиственных лесов // Вестн. Казанского гос. аграрного ун-та. Казань, 2009. № 2(12). С. 155–160.
Жукова Л.А., Турмухаметова Н.В., Акшенцев Е.В. Экологическая характеристика некоторых видов растений // Онтогенетический атлас растений. Йошкар-Ола, 2007. Т. 5. С. 318–331.
Ter Braak C.J.F., Šmilauer P. CANOCO Reference manual and CanoDraw for Windows User’s guide: Software for Canonical Commuhity Ordination (version 4.5). Microcomputer Power. Ithaca, NY, USA, 2002. P. 500.
Hutchinson G.E. A treatise on limnology. Vol. I. Geography, Physics and Chemistry. New York: John Willey and Sons, 1957. 1015 p.
Whittaker R.H. Communities and ecosystems. New York: Macmillan, 1975. 385 p.
Пианка Э. Эволюционная экология. М.: Мир, 1981. 400 с.
Онипченко В.Г. Функциональная фитоценология: Синэкология растений. Учеб. пособ. 2014. 576 с.
Щеголева Н.В., Зверев А.А. Особенности экологического ареала лютиков (Ranunculus) Cибири // Итоги и перспективы геоботанических исследований в Сибири: Мат-лы Всеросс. науч.-практ. конф. Новосибирск, 2019. С. 124–125.
Щеголева Н.В., Зверев А.А. Особенности экологического ареала лютиков (Ranunculus) Cибири // Проблемы промышленной ботаники индустриально развитых регионов: Мат-лы V Междунар. конф. Кемерово, 2018. С. 132–135.
Юсупова О.В., Абрамова Л.M., Юсупов И.Р. Особенности организации популяций высокогорного эндемичного вида Anemonastmm biarmiense (Juz.) Holub s.1. на Южном Урале // Вестн. Удмуртского ун-та. Биология. Науки о земле. 2017. Т. 27. Вып. 2. С. 171–179.
Юсупова О.В., Абрамова Л.M., Юсупов И.Р. Динамика популяций Anemonastmm biarmiense (Ranunculaceae) в Южно-Уральском заповеднике (Республика Башкортостан) // Ботанич. журн. 2020. Т. 105. № 9. С. 29–45. https://doi.org/10.31857/S0006813620090100
Ленская О.Ю., Ботова М.Г. Особенности текущих климатических изменений в регионе Южного Урала // Вестн. Челябинского гос. ун-та. 2011. № 5(220). С. 44–49.
Оценочный доклад об изменениях климата и их последствиях на территории Российской Федерации: техн. резюме [Электронный ресурс] / Под ред. Мелешко В.П., Семенова С.М. М.: Росгидромет, 2008. 89 с. URL: www.meteoinfo.ru/media/climate/tehnicheskoe_rezume_rus.pdf
Шиятов С.Г., Мазепа В.С. Использование повторных ландшафтных снимков для оценки динамики лесотундровых сообществ на Полярном Урале // Леса России и хозяйство в них. 2015. № 3(54). С. 20–28.
Моисеев П.А., Шиятов С.Г., Григорьев А.А. Климатогенная динамика древесной растительности на верхнем пределе ее распространения на хребте Большой Таганай за последнее столетие. Екатеринбург, 2016. 136 с.
Моисеев П.А., Бубнов М.Ю., Дэви Н.М., Нагимов З.Я. Изменение структуры и фитомассы древостоев на верхнем пределе их произрастания на Южном Урале // Экология. 2016. № 3. С. 219–227. https://doi.org/10.1134/S1067413616030085
Шиятов С.Г., Терентьев М.М., Фомин В.В., Циммерманн Н.Е. Вертикальный и горизонтальный сдвиги верхней границы редколесий и сомкнутых лесов в XX столетии на Полярном Урале // Экология. 2007. № 4. С. 243–248.https://doi.org/10.1134/S1067413607040017
Дополнительные материалы отсутствуют.