

Экология, 2021, № 6, стр. 460-469
Трофические взаимодействия белого гуся и черной казарки в период размножения в аспекте популяционной динамики
С. Б. Розенфельд a, И. С. Шереметьев b, *, В. В. Баранюк c
a Институт проблем экологии и эволюции им. А.Н. Северцова РАН
119071 Москва, Ленинский просп., 33, Россия
b ФНЦ биоразнообразия наземной биоты Восточной Азии ДВО РАН
690022 Владивосток, просп. 100-летия Владивостока, 159, Россия
c Всероссийский научно-исследовательский институт охраны окружающей среды (ВНИИ Экология)
117628 Москва, 36 км МКАД, влд. 1, стр. 4, Россия
* E-mail: sheremetyev@biosoil.ru
Поступила в редакцию 05.03.2021
После доработки 26.05.2021
Принята к публикации 30.05.2021
Аннотация
Изучены различия в использовании трофических ресурсов в репродуктивный период самого многочисленного в Арктике белого гуся Anser caerulescens и самой широко распространенной черной казарки Branta bernicla в условиях основной части гнездового ареала гусей (дельта р. Колвилл, п-ов Аляска). Установлено, что диеты этих видов состоят преимущественно из злаков и кустарничков, в меньшей степени – из мхов, осок, хвощей и разнотравья и почти не имеют различий по составу ресурсных наименований. Также обнаружено, что у обоих видов наибольшие затраты на кормодобывание наблюдаются в береговых местообитаниях, наименьшие – в маршевых. При отсутствии существенной дифференциации трофических ниш у белого гуся значительно меньше затраты на кормодобывание, чем у черной казарки. Предлагается вероятный механизм трансформации включающих эти виды сообществ на основе трофических взаимодействий.
Гуси и казарки (подсем. Anserinae), потребляющие колоссальный объем фитомассы в арктической экосистеме, размножаются и восстанавливают энергетические потери в условиях жестких временны́х, территориальных и в конечном счете трофических ограничений [1–4]. Поэтому общность ресурсов, неизбежная у таких систематически и экологически близких и в основном широкоареальных видов [5], связывает анализ их трофических взаимодействий с важнейшими задачами эволюционной экологии [6]. Однако значение этих взаимодействий в организации сообществ, включающих гусей и казарок, остается все еще неясным.
Первоочередную актуальность в исследованиях этих сообществ представляет объяснение различий между популяционными трендами белого гуся Anser caerulescens и черной казарки Branta bernicla, так как после глубокой депрессии в ХХ в. численность первого вида экспоненциально растет и достигла уже 15 млн, а у второго едва сохраняется на уровне 630 тыс. [5, 7]. При этом надо учитывать, что гнездовой ареал у гусей ограничен в основном Северной Америкой и Гренландией, а у казарок – циркумполярен [1, 5]. Однако очевидная смена в составе доминирующих арктических фитофагов до сих пор рассматривалась преимущественно в аспекте глобального климатического или антропогенного влияния [2, 8, 9], а взаимодействия между ними на основной части общего ареала – только в аспекте дифференциации местообитаний [10]. Анализ эволюционной сущности трансформации сообществ, включающих эти виды, пусть даже на фоне возрастающего влияния извне, вряд ли возможен посредством таких упрощений.
Основа механизма трансформации сообществ, связанной с взаимодействиями экологически сходных видов, изложена в концепции асимметричной конкуренции, исход которой на межвидовом уровне определен специфическим неравенством соперников в освоении общих ресурсов [11]. В каждом конкретном случае остается только выяснить детали этого неравенства и по соответствию популяционным трендам – их значение на уровне сообщества. Анализ неравного раздела ресурсов у гусей и казарок опирается на их морфофизиологические ограничения [5, 12, 13], место которых в динамике сообщества, включающего белого гуся и черную казарку, уже было показано на о-ве Врангеля [14]. Важнейшим новым шагом в данной работе становится характеристика асимметрии не только ресурсных долей, но и затрат на их освоение и не в нетипичных островных условиях на краю ареала белого гуся, а в его основной части, в Северной Америке.
Хорошая изученность растительности пастбищ североамериканских гусей и казарок [15, 16] впервые позволяет проанализировать и сами трофические различия, и их настоящие причины, превзойдя опосредованные характеристики конкуренции. В рассматриваемом механизме трансформации глубокий интерес представляет возможность ранжирования трофических соперников по шкале r/K-стратегий [17]. Доминирование соперника с r-стратегией освоения ресурсов сопутствует кризису сообщества, доминирование K-стратега – напротив, его долгосрочно стабильному развитию [18, 19]. Если потребителем лучших по качеству, но и требующих бóльших затрат ресурсов является черная казарка, популяция которой за много лет недалеко ушла от рекордного минимума, то перед нами всего лишь сукцессия арктических фитофагов, а сукцессия – атрибут когерентной фазы развития сообществ, характеризуемой минимальным влиянием внешних, в том числе глобальных климатических и/или антропогенных изменений [18]. Если таким r-стратегом является белый гусь, то быстрый рост его популяции – показатель масштабного, вероятно, экзогенного кризиса.
Данные об истощении ресурсов североамериканских летних пастбищ этих видов гусей и казарок [20, 21] позволяют в рабочем порядке предполагать, что исход их асимметричной конкуренции зависит, скорее, от различий в затратах на кормодобывание, чем в ширине ниш, в том числе и за счет важнейших ресурсов, т.е. в качестве питания. Значение успешного размножения, определяемого в первую очередь выживаемостью кладок и птенцов, множество которых гибнет во время и после насиживания [1, 5, 22], в приросте популяций белого гуся и черной казарки указывает на ключевую роль этой части годового цикла в анализе взаимодействий между ними [23–25]. Главная цель нашей работы – изучить особенности трофических взаимодействий белого гуся и черной казарки в период размножения и выяснить их значение в трансформации включающих эти виды сообществ.
МАТЕРИАЛ И МЕТОДЫ
Северное побережье Аляски, где динамика и размерное соотношение популяций белого гуся и черной казарки отражают положение дел не только в Западной Арктике, но и во всем мире [2, 7, 26, 27], имеет большое значение. Здесь ареной интенсивных исследований стали типичные для арктического побережья США и Канады места гнездования этих видов гусей и казарок в дельте р. Колвилл (650 км2), находящейся на побережье моря Бофорта в подзоне гипоарктических тундр [2, 28]. Можно выделить до девяти типов растительных ассоциаций, приуроченных к трем типам ландшафта (табл. 1): приморские марши с доминированием галофитов, материковые заозеренные тундры и береговые тундры [2, 15, 16].
Таблица 1.
Растительные ассоциации (1–9) типов местообитаний гусей и казарок в дельте р. Колвилл (Аляска, США)
| Типы местообитаний | № | Доминанты* |
|---|---|---|
| Берега озер | 1 | Мхи, Deschampsia |
| 2 | Carex, Eriophorum, мхи | |
| Марши | 3 | Dupontia, Arctophila, Carex, Eriophorum** |
| 4 | Carex (C. subspathacea), Eriophorum, Puccinellia | |
| 5 | Carex (C. ramenskii), Eriophorum, Stellaria | |
| 6 | Puccinellia, Carex, Eriophorum | |
| Морские берега | 7 | Злаковые, разнотравье (галечники) |
| 8 | Salix, злаковые, мхи (террасы) | |
| 9 | Злаковые, разнотравье, Artemisia (дюны) |
Кормовые пробы представлены экскрементами гусей (n = 21) и казарок (n = 15), собранными В.В. Баранюком в мае‒июле 2012 г. в местах их концентрации без непосредственного контакта. Ресурсные наименования (растения) в кормовых пробах идентифицированы по методу M. Owen [29] с использованием эталонных микрофотографий [30]. При характеристике растительности пастбищ использованы опубликованные данные R. Taylor [16], D. Derksen et al. [15] и J. Hupp et al. [2]. Растения в сети данных по типам ассоциаций и кормовым пробам объединены в основном на уровне родов (табл. 2). UPGMA-классификация проб и типов ассоциаций, в том числе в тестировании их ландшафтного распределения, выполнена на основе соотношения ресурсов по частоте встречаемости. Избирательность ресурсов определена по индексу Джэйкобса D [31]:
где ci – доля i ресурса в пробах, ai – доля i ресурса на пастбище каждого типа местообитаний. Ресурсные наименования распределены по трем категориям, соответствующим интервалам значений D [–1; 1] по В.С. Ивлеву [32]: D ∈ [0.33; 1] – элективные (предпочитаемые), D ∈ (–0.33; 0.33) – нейтральные (оппортунистические), D ∈ [–1; –0.33] – дискриминируемые (избегаемые).Таблица 2.
Вклад избирательности ресурсных наименований гусей и казарок в дельте р. Колвилл (Аляска) в дифференциацию их проб
| Ресурсы | Стандартизированные коэффициенты для первых канонических переменных | |
|---|---|---|
| F1 | F2 | |
| Alopecurus | –0.03236 | 0.39573 |
| Arctagrostis | 0.26501 | 0.42612 |
| Bryophyta | 0.31463 | 0.46878 |
| Calamagrostis | 0.52678 | 0.00691 |
| Carex/Eriophorum | –1.31355 | –0.66722 |
| Deschampsia | –1.50848 | 5.29162 |
| Dupontia/Arctophila | –0.39562 | –0.45843 |
| Equisetum | 1.42610 | 0.33209 |
| Festuca | 1.31374 | –0.47208 |
| Poa | 0.36494 | 0.06735 |
| Poaceae* | 2.75391 | 0.27526 |
| Puccinellia | –0.21995 | –0.28301 |
| Salix | 0.29144 | 0.71501 |
| Stellaria | 0.61020 | –2.03368 |
| Разнотравье* | 0.80293 | 0.73664 |
Ранжирование гусей и казарок по избирательности выполнено по (1) количественной доле элективных ресурсов в диете (% числа фрагментов эпидермиса), (2) доле элективных ресурсов в составе диеты (% числа ресурсов) и (3) отношению первого показателя ко второму. Последний интерпретирован как индекс избирательного усилия, позволяющий соотносить затраты рассматриваемых видов на кормодобывание. Вид с бóльшей количественной долей элективных ресурсов и/или меньшей их долей в составе диеты и, таким образом, с бóльшим избирательным усилием ранжирован как более избирательный. Дифференциация гусей и казарок по составу трофического спектра оценена с использованием индекса Шимкевича-Симпсона [33], по избирательности – с использованием коэффициента корреляции Спирмена между значениями D ресурсов. Все расчеты выполнены в Statistica 13 [34].
РЕЗУЛЬТАТЫ
Кормовые растения и классификация пастбищ. На переувлажненных и затопленных участках доминируют виды родов Carex, Eriophorum, Arctophila и Hippuris, на периодически затопляемых морем понижениях – C. subspathacea, к которой на поднятиях добавляются виды родов Dupontia, Poa, Puccinellia, Salix и разнотравье [2]. Ассоциации объединяются в три UPGMA-группы (типы местообитаний) близкого уровня максимальных внутрикластерных дистанций (рис. 1), которые в основном соответствуют их ландшафтной приуроченности по данным R. Taylor [16] и D. Derksen et al. [15]. Две группы – это марши и береговые тундры по R. Taylor [16], исключая ассоциации с доминированием видов рода Dupontia, которые по количественному соотношению ресурсов ближе к приозерным ассоциациям по D. Derksen et al. [15] (см. рис. 1). Береговые местообитания отличаются разнообразием злаков, доля которых везде велика, но на маршах они представлены в основном родом Puccinellia. Приозерные местообитания отличаются особенно большой долей мхов, маршевые – осок, а береговые – разнотравья (двудольные) (см. рис. 1).
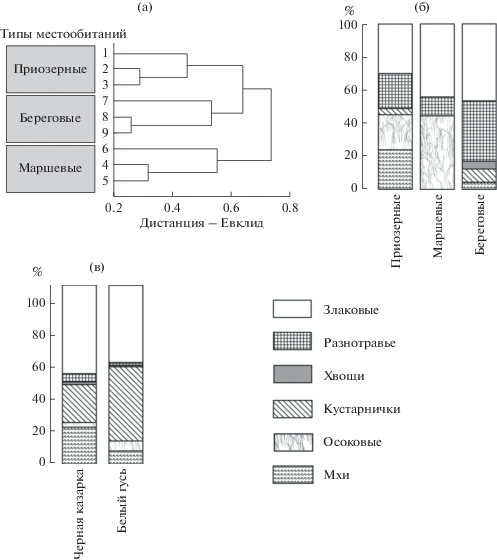
Трофические спектры гусей и казарок. В пробах гусей определены виды родов Alopecurus, Arctagrostis, Arctophila, Calamagrostis, Carex, Deschampsia, Dupontia, Equisetum, Eriophorum, Festuca, Luzula, Poa, Puccinellia, Salix, Stellaria и фрагменты, относящиеся к сем. Poaceae, Bryophyta и разнотравью. Из них у казарок не найдены только виды рода Arctagrostis. Индекс Шимкевича-Симпсона, характеризующий сходство гусей и казарок по составу ресурсов в совокупности межвидовых пар проб, равен 53%, что немного меньше внутривидовых значений (62% у казарок и 60% у гусей). У обоих видов количественно доминируют злаки, а оставшиеся более 41% у гусей это в основном кустарнички, у казарок – по 21% кустарничков и мхов (см. рис. 1). Эти различия нельзя связать с неопределенными фрагментами злаков и разнотравья, доля которых составляет единичные проценты. По суммарной количественной доле злаков и кустарничков мало различий.
Распределение ресурсов по избирательности. К элективным ресурсам наиболее часто относятся ивы, осоки, мхи и разнотравье. Отмеченные на пастбищах представители Androsace, Artemisia, Caltha, Cassiope, Koenigia, Myosotis, Papaver, Pedicularis, Ranunculus, Saxifraga, Potentilla, Juncus и Polygonum [15, 16] во всех пробах отнесены только к нейтральным или дискриминируемым ресурсам каждого из трех типов местообитаний. Только последние три отмечены в питании этих видов на арктическом побережье России, где в пробах казарок найден также Arctagrostis [23, 35].
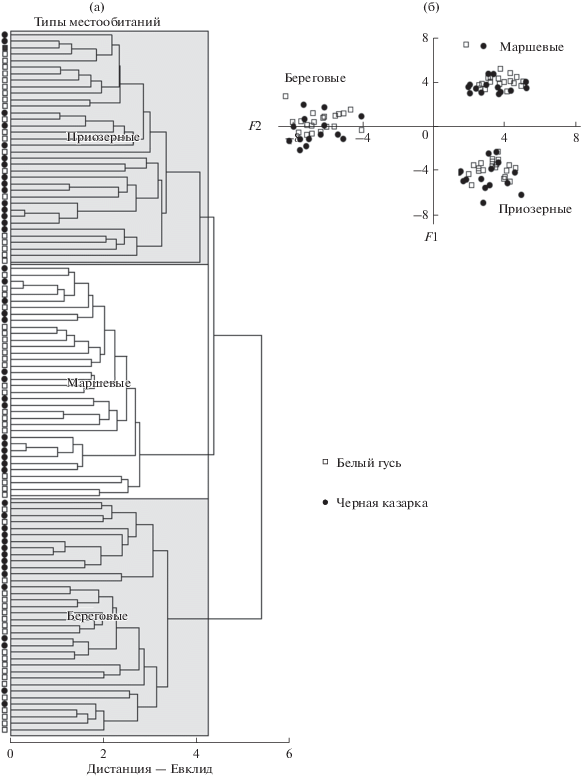
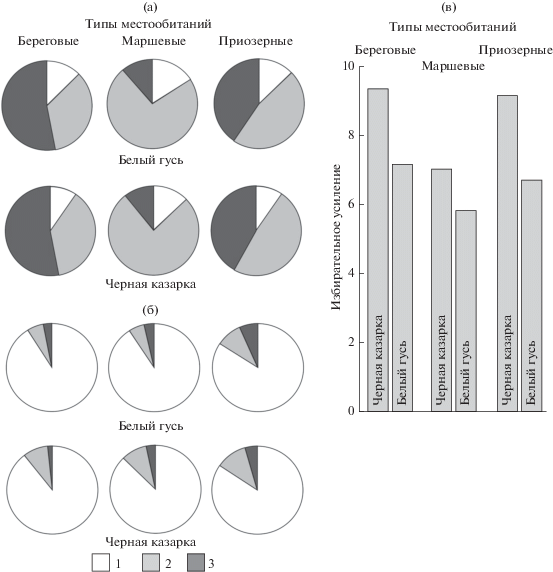
Классификация проб и различия видов по избирательности. В UPGMA-классификацию проб по избирательности включенных в них ресурсов определяющий вклад вносят различия типов местообитаний, который намного больше, чем у видовой принадлежности проб. Поэтому избирательность гусей и казарок следует оценивать для каждого типа местообитаний отдельно (рис. 2). Наибольший вклад в распределение проб в пространстве первых канонических переменных (F1 и F2) по избирательности у Salix, Carex/Eriophorum, Equisetum, нескольких злаков и разнотравья (табл. 2). Очевидна и роль межвидовых различий по избирательности ресурсов, не имеющих существенных различий по количественной доле на пастбищах. В трофическом спектре гусей и казарок во всех типах местообитаний общее число элективных ресурсов почти одинаково – 12–13 и 13–14 соответственно. Однако их доля в составе проб гусей в среднем на 26–34% больше, чем у казарок, в основном за счет нейтральных ресурсов, особенно на маршах (рис. 3). Совокупные межвидовые различия по количественной доле элективных ресурсов, составляющей 84–91%, очень небольшие. Представляют интерес межвидовые различия между количественной долей элективных ресурсов и их долей в составе диет. Последние обусловливают аналогичные различия по избирательному усилию, которое у гусей на 22–36% меньше, чем у казарок (см. рис. 3). Межвидовая корреляция проб по избирательности ресурсных списков всегда позитивна при p < 0.05 в 92% случаев для приозерных местообитаний, в 82% – для маршевых и в 97% – для береговых.
ОБСУЖДЕНИЕ
Трофические спектры белого гуся и черной казарки на исследованной территории почти не различаются по составу ресурсов даже на индивидуальном уровне, а с данными по о-ву Врангеля [23] совпадают. Близкие суммарные доли злаков и кустарничков – 85% у гусей и 71% у казарок, исключают возможность и количественной дифференциации ниш как последней основы стабильного сосуществования. Это увеличивает потенциал асимметричной конкуренции, однако наиболее простая характеристика асимметрии – по ширине ресурсных ниш [36], исключена их полным перекрыванием у сравниваемых видов. Также можно было считать преимуществом преобладание у гусей суммарной доли злаков и кустарничков и существенно меньшую долю мхов (см. рис. 1). Мхи – не лучшие корма арктических травоядных [35, 37], и их употребление казарками указывает на бóльший недостаток ресурсов, чем у гусей. Однако межвидовые различия по количественной доле элективных ресурсов, т.е. по качеству питания, в итоге незначительны (см. рис. 3). При условиях жестких трофических ограничений это позволяет заключить, что определяющее значение в конкуренции данных видов имеет неравенство затрат на освоение ресурсов.
Концепция энергетических стратегий, связывающая эффективность затрат травоядных в первую очередь с увеличением размера тела [38–40], позволяет на основании морфофизиологических ограничений предполагать наименее затратное кормодобывание у гусей. Их физическая сила и пропорции клюва также существенно расширяют возможности использовать грубые корма [35, 41], в том числе в качестве элективных ресурсов. Различия в средней массе тела – 2.6 кг у гусей и 1.54 кг у казарок [5, 35], позволяют оценить относительную приспособленность первых к менее избирательному и, таким образом, менее затратному питанию.
Почти все свое время оба сравниваемых вида проводят в поисках корма [1, 35], и даже визуальное сопоставление количественной доли элективных ресурсов с их долей в составе диеты (см. рис. 3) показывает, чего эти поиски стоят каждому из них. Злаки, которых много и в трофических спектрах, и на пастбищах всех типов (см. рис. 1), можно было считать оппортунистическим компонентом. Однако высшие кормовые качества Puccinellia среди маршевых и Arctophila с Dupontia среди приозерных злаков свидетельствуют об обратном [2, 35], а количественное сопоставление других ресурсов в пробах и на пастбищах ясно указывает на большую избирательность обоих видов. На маршах, в том числе с доминированием C. subspathacea, которые гуси и казарки используют с интенсивностью грэйзинг-газона [4, 42], характеризуемые избирательным усилием затраты (см. рис. 3) минимальны, в береговых местообитаниях – максимальны. Во всех типах местообитаний затраты гусей существенно меньше, чем у казарок.
Различия в затратах на кормодобывание соответствуют различиям в динамике численности местной популяции белого гуся, выросшей за четверть века с около 0.5 тыс. до более чем 30 тыс. [26, 27], и черной казарки, численность которой увеличивалась за это время в три раза медленнее, не достигнув и 6 тыс. [2]. Аналогичные различия в размерах и динамике западно-арктических и мировых популяций этих видов [1, 5, 7] позволяют связать трансформацию включающих их сообществ в Арктике с асимметричной трофической конкуренцией. Здесь важно подчеркнуть, что речь может идти о расселении белого гуся в Евразии за пределы о-ва Врангеля [1], а о сопутствующем вымирании или даже сокращении мировой популяции черной казарки пока не может. Однако где остановится рост численности гусей в равных с казарками условиях и после одинаково тяжелой депрессии в ХХ в. [5], мы не знаем, а демографическое и конкурентное неравенство этих видов не оставляет шансов для предлагаемого S. Hubbell [43] нейтрального сценария взаимодействий.
В детализации неравного раздела ресурсов важно также учитывать, что и до, и во время насиживания гуси вынуждены питаться интенсивнее, так как их яйцевые фолликулы формируются не за счет накоплений жира и протеинов, как у казарок, а за счет питания в местах гнездования после прилета [9]. Гнездовые участки обоих видов ассоциированы с береговыми тундрами, где раньше сходит снег [1, 9, 44] и больше растений, к питанию которыми гуси более приспособлены, чем казарки (см. рис. 1). Таким образом, сначала насиживающие гуси – основные потребители растительности береговых местообитаний, питание которой у обоих видов наиболее затратно (см. рис. 3), затем, когда выводки обоих видов уходят на марши, где кормятся до полного оперения [2, 4, 42], снятие ограничений насиживания сопутствует увеличению кормовых потребностей, которому наиболее соответствуют марши с C. subspathacea. И в это время, когда рассчитывать на какую-либо пространственную дифференциацию кормящихся в непосредственной близости гусей и казарок тоже нет оснований [2], питание гусей с более широким спектром элективных ресурсов остается менее затратным. В итоге и на маршах, и в прилегающей тундре затраты на кормодобывание у гусей меньше, их ресурсная доля больше, а казарки вынуждены использовать еще и приозерные местообитания, где их затраты на кормодобывание тоже больше. Иначе ни большую долю мхов в питании (см. рис. 1), ни отбытие потерявших птенцов казарок на время линьки за пределы изученной территории, когда такие гуси во множестве остаются [2, 10, 45], объяснить нельзя.
В реконструкции механизма трансформации сообществ, включающих эти виды, критическое значение имеют условия жестких ограничений, существенно усиливаемых конкуренцией. Увеличение затрат в связи с необходимостью в качественном питании при ресурсном ограничении возможно только за счет снижения количества потребляемой казарками фитомассы, перераспределяемой таким образом в пользу гусей. Это снижает в итоге успех размножения и годовой прирост популяции казарок в бóльшей степени, чем у гусей, и обеспечивает описанные выше различия в популяционных трендах. Увеличение популяции гусей в свою очередь снижает количество фитомассы, доступной казаркам, которые вынуждены дополнительно увеличивать затраты на кормодобывание, и так до тех пор, пока отставание роста популяции казарок не сменится сокращением их численности. Усиление трофических ограничений сопутствует, вероятно, снижению безопасности выводков, которое также негативно может отражаться на приросте популяций.
Как более избирательный конкурент за трофические ресурсы черная казарка должна быть ранжирована как r-стратег относительно белого гуся. Долгое отсутствие восстановления популяции казарок после депрессии на фоне быстрого и стабильного роста популяции гусей позволяет определить трансформацию включающих эти виды сообществ на основе асимметричной конкуренции как сукцессию, которая является надежным признаком несущественности любого внешнего влияния [18, 19]. Действительно, влияние дифференцированного истребления гусей и казарок человеком можно исключить ввиду всестороннего контроля природопользования в западном секторе Арктики [5, 7], а влияние глобального потепления может, скорее, дополнять, чем противодействовать, влияние ресурсного неравенства, улучшая условия фолликулообразования у гусей и, вероятно, ухудшая у казарок [9], хотя такое объяснение еще нуждается в дополнительных данных. Также влияние других, обычных в Арктике травоядных вряд ли можно считать существенным: северный олень Rangifer tarandus из-за гнуса уходит в период размножения гусей и казарок за пределы исследованной территории; белолобый гусь A. albifrons тяготеет к внутренним водоемам и тундре [2, 46]; лемминги (Arvicolinae), которые в годы пиков тоже осваивают большое количество фитомассы [47], на маршах могут быть исключены [2], а в тундре, скорее, сами испытывают негативное влияние гусей [14, 23 ]. В этих условиях сложно допустить, что росту популяции белого гуся в обозримом будущем сможет помешать только какой-то еще более выраженный K-стратег среди травоядных.
Сукцессионный детерминизм [48] предполагает только две основные возможности восстановления западно-арктической популяции черной казарки после депрессии ХХ в.: 1 – катастрофическое освобождение ресурсной ниши, после которого ее в первую очередь занимают r-стратеги; 2 – снижение смертности черной казарки вне сезона размножения. Примером первого является интенсивное истребление всех гусей и казарок до середины ХХ в. [5], цивилизованной заменой которого может стать дифференцированное ограничение популяции белого гуся в США и Канаде. Второе можно пока представить только в виде усилий по сохранению зимовочных местообитаний черной казарки и/или в виде ее адаптации к зимнему питанию сельскохозяйственными растениями, отмеченной пока только в Нидерландах [49].
Самостоятельный интерес представляет вопрос о значении морфофизиологических характеристик казарок в целом, сопутствующих их более избирательному питанию, чем у гусей [5]. Все дело в том, что последний аудит популяций Anserinae в Арктике приравнивает суммарную численность всех пяти видов казарок к 13.7 млн и делит их на 27 популяций, треть из которых сокращается [7], в то время как все шесть популяций только трех видов филогруппы Chen, включающей белого гуся, растут и составляют в сумме уже 17.2 млн [7]. Можно ли в связи с этим считать казарок, в целом представленных большим числом мелких популяций, в отсутствие каких-то существенных нарушений среды обитания потенциально редкими и/или уязвимыми видами по сравнению с гусями, представленными малым числом относительно крупных популяций?
ЗАКЛЮЧЕНИЕ
Из представленных выше результатов вытекает главный вывод – в равных условиях у белого гуся энергетические затраты на кормодобывание существенно меньше, чем у черной казарки, при отсутствии качественных и малых количественных различиях между их трофическими спектрами. Различия между популяционными трендами этих видов на исследованной территории так же, как в Западной Арктике и на мировом уровне, объясняются асимметричной трофической конкуренцией между ними при незначительном влиянии других обычных в Арктике фитофагов.
Соотношение r/K-стратегий освоения трофических ресурсов у белого гуся и черной казарки определяет трансформацию включающих эти виды сообществ как сукцессию, позволяя отнести эту трансформацию в настоящее время к когерентной фазе развития сообществ. Явное суммарное доминирование в Арктике всех популяций трех видов Anser, морфофизиологические характеристики которых соответствуют меньшей избирательности, над всеми популяциями всех пяти видов Branta дает основание предполагать аналогичное положение дел у Anserinae в целом. Таким образом, все или почти все виды Branta можно считать потенциально редкими или уязвимыми в стабильных условиях. Это может свидетельствовать о преимущественно естественном характере сегодняшних изменений в сообществах арктических фитофагов, а не об их экзогенной катастрофе, в том числе под влиянием глобальных климатических или антропогенных изменений.
В процессе работы животные не подвергались физическим контактам с исследователями. Были приложены специальные усилия, чтобы уменьшить беспокойство животных во время сбора образцов.
Авторы заявляют об отсутствии конфликта интересов.
Список литературы
Сыроечковский Е.В. Пути адаптации гусеобразных трибы Anserini к обитанию в Арктике. М.: КМК, 2013. 297 с.
Hupp J.W., Ward D.H., Hogrefe K.R. et al. Growth of black brant and lesser snow goose goslings in northern Alaska // J. Wildlife Manage. 2017. V. 81. P. 846–857. https://doi.org/10.1002/jwmg.21246
Lake B.C., Schmutz J.A., Lindberg M.S. et al. Body mass of prefledging emperor geese Chen canagica: large-scale effects of interspecific densities and food availability // Ibis. 2008. V. 150. P. 527–540. https://doi.org/10.1111/j.1474-919X.2008.00814.x
Person B.T., Herzog M.P., Ruess R.W. et al. Feedback dynamics of grazing lawns: coupling vegetation change with animal growth // Oecologia. 2003. V. 135. P. 583–592. https://doi.org/10.1007/s00442-003-1197-4
Розенфельд С.Б., Шереметьев И.С. Арктические гуси (Anser) и казарки (Branta) Евразии: анализ факторов динамики численности и ареалов // Журн. общ. биол. 2016. Т. 77. № 1. С. 16–37. https://doi.org/10.1134/S2079086416050078
Thompson J.N. The evolution of species interactions // Science. 1999. V. 284. P. 2116–2118. https://doi.org/10.1126/science.284.5423.2116
CAFF International Secretariat: A Global audit of the status and trends of Arctic and Northern Hemisphere goose population. 2018. Akureyri, Iceland. 32 p.
Ims R.A., Fuglei E. Trophic interaction cycles in tundra ecosystems and the impact of climate change // BioScience. 2005. V. 55. P. 311–322. https://doi.org/10.1641/0006-3568(2005)055[0311: TICITE]2.0.CO;2
Hupp J.W., Ward D.H., Soto D.X. et al. Spring temperature, migration chronology, and nutrient allocation to eggs in three species of arctic-nesting geese: Implications for resilience to climate warming // Glob. Change Biol. 2018. V. 24. № 11. P. 5056–5071. https://doi.org/10.1111/gcb.14418
Flint P.L., Mallek E.J., King R J. et al. Changes in abundance and spatial distribution of geese molting near Teshekpuk Lake, Alaska: interspecific competition or ecological change? // Polar Biol. 2008. V. 31. P. 549–556. https://doi.org/10.1007/s00300-007-0386-8
Freckleton R.P., Watkinson A.R. Asymmetric competition between plant species // Func. Ecol. 2001. V. 15. P. 615–623. https://doi.org/10.1046/j.0269-8463.2001.00558.x
Sedinger J.S. Adaptations to and consequences of an herbivorous diet in grouse and waterfowl // Condor. 1997. V. 99. P. 314–326. https://doi.org/10.2307/1369937
Розенфельд С.Б., Шереметьев И.С. Сравнительный анализ питания дикого северного оленя (Rangifer tarandus), белолобого гуся (Anser albifrons) и черной казарки (Branta bernicla) на о-вах Белый и Шокальского (ЯНАО) // Зоол. журн. 2020. Т. 99. Вып. 9. С. 1036–1046. https://doi.org/10.31857/S0044513420090147
Шереметьев И.С., Розенфельд С.Б., Груздев А.Р. Перекрывание трофических спектров жвачных, гусей и леммингов на о-ве Врангеля в летний период // Экология. 2017. № 6. С. 440–446. [Sheremetyev I.S., Rozenfeld S.B., Gruzdev A.P. Dietary overlap among ruminants, geese, and lemmings of Wrangel Island in summer // Russ. J. Ecol. 2017. V. 48. P. 532–538.] https://doi.org/10.1134/S1067413617060091
Derksen D., Eldridge W.D., Weller M.W. Habitat ecology of Pacific black brant and other geese moulting near Teshekpuk Lake, Alaska // Wildfowl. 1982. V. 33. P. 39–57.
Taylor R.J. Shoreline vegetation of the Arctic Alaska coast // Arctic. 1981. V. 34. № 1. P. 37–42.
MacArthur R.H., Wilson E.O. The theory of island biogeography. Princeton: Princeton Univ. Press, 1967. 203 p.
Раутиан А.С., Жерихин В.В. Модели филоценогенеза и уроки экологических кризисов геологического прошлого // Журн. общ. биол. 1997. Т. 58. № 4. С. 20–47.
Красилов В.А. Нерешенные проблемы теории эволюции. Владивосток: ДВНЦ АН СССР, 1986. 140 с.
Jano A.P., Jefferies R.L., Rockwell R.F. The detection of change by multitemporal analysis of LANDSAT data: the effects of goose grazing // J. Ecol. 1998. V. 86. P. 93–99.
Alisauskas R.T., Charlwood J.W., Kellett D.K. Vegetation correlates of the history and density of nesting by Ross’s geese and lesser snow geese at Karrak Lake, Nunavut // Arctic. 2006. V. 59. P. 201–210. https://doi.org/10.14430/arctic342
Розенфельд С.Б., Сыроечковский Е.Е. Питание черной казарки в тундрах России в период размножения // Казарка. 1998. № 4. С. 96–120.
Розенфельд С.Б., Шереметьев И.С., Баранюк В.В. Малый белый гусь на о. Врангеля: трофические связи со жвачными и выбор репродуктивных местообитаний // Зоол. журн. 2017. Т. 96. Вып. 3. С. 1–11. https://doi.org/10.1134/S1062359017090102
Cooch E.G., Lank D.B., Rockwell R.F. et al. Long-term decline in body size in a snow goose population: evidence of environmental degradation? // J. Anim. Ecol. 1991. V. 60. P. 483–496. https://doi.org/10.2307/5293
Richman S.E., Leafloor J.O., Karasov W.H. et al. Ecological implications of reduced forage quality on growth and survival of sympatric geese // J. Animal Ecol. 2015. V. 84. P. 284–298. https://doi.org/10.1111/1365-2656.12270
Johnson S.R., Herter D.R. The birds of the Beaufort Sea. Anchorage, Alaska: BP Exploration, 1989. 372 p.
Burgess R.M., Ritchie R.J., Person B.T. et al. Rapid growth of a nesting colony of lesser snow geese (Chen caerulescens caerulescens) on the Ikpikpuk River Delta, North Slope, Alaska, USA // Waterbirds. 2017. V. 40. P. 11–23. https://doi.org/10.1675/063.040.0103
Walker H.J. Guidebook to permafrost and related features of the Colville River Delta, Alaska. Forth International Conference on Permafrost, Fairbanks, Alaska, USA, 1983. 33 p.
Owen M. An assessment of fecal analyses technique in waterfowl feeding studies // J. Wildlife Manage. 1975. V. 39. P. 271–279.
Carriere S. Photographic key for the microhistological identification of some Arctic vascular plants //Arctic. 2002. V. 55. № 3. P. 247–268. https://doi.org/10.14430/arctic709
Jacobs J. Quantitative measurement of food selection. A modification of the forage ratio and Ivlev’s electivity index // Oecologia. 1974. V. 14. P. 413–417.
Ивлев В.С. Экспериментальная экология питания рыб. М.: Пищепромиздат, 1955. 252 с.
Песенко Ю.А. Принципы и методы количественного анализа в фаунистических исследованиях. М.: Наука, 1982. 288 с.
TIBCO Software Inc.: Statistica 13. 2017. http:// statistica.io
Розенфельд С.Б. Питание казарок и гусей в Российской Арктике. М.: КМК, 2009. 236 с.
Greenberg R. Ecological plasticity, neophobia, and resource use in birds // Stud. Avian Biol. 1990. V. 13. P. 431–437.
Soininen E.M., Zinger L., Gielly L. et al. Not only mosses: lemming winter diets as described by DNA metabarcoding // Polar Biol. 2017. V. 40. P. 2097–2103. https://doi.org/10.1007/s00300-017-2114-3
Bergman C.M., Fryxell J.M., Gates C.C. et al. Ungulate foraging strategies: energy maximizing or time minimizing? // J. Anim. Ecol. 2001.V. 70. P. 289–300. https://doi.org/10.1111/j.1365-2656.2001.00496.x
Gordon I.J., Prins H.H.T. Introduction: grazers and browsers in a changing world // The ecology of browsing and grazing. V. 195. Berlin: Springer, 2008. P. 1–20.
Clauss M., Steuer P., Muller D.W. et al. Herbivory and body size: Allometries of diet quality and gastrointestinal physiology, and implications for herbivore ecology and dinosaur gigantism // PLOS ONE. 2013. V. 8. P. e68714. https://doi.org/10.1371/journal.pone.0068714
Кондратьев А.В. Экология питания гусей в Арктике и на пути к ней // Казарка. 2002. № 8. С. 79–101.
Ruess R.W., Uliassi D.D., Mulder C.P.H. et al. Growth responses of Carex ramenskii to defoliation, salinity, and nitrogen availability: implications for goose-ecosystem dynamics in western Alaska // Ecoscience. 1997. V. 4. P. 170–178. https://doi.org/10.1080/11956860.1997.11682392
Hubbell S.P. Neutral theory in community ecology and the hypothesis of functional equivalence // Func. Ecol. 2005. V. 19. P. 166–172. https://doi.org/10.1111/j.0269-8463.2005.00965.x
Ward D.H., Helmericks J., Hupp J.W. et al. Multi-decadal trends in spring arrival of avian migrants to the central Arctic coast of Alaska: Effects of environmental and ecological factors // J. Avian Biol. 2016. V. 47. P. 197–207. https://doi.org/10.1111/jav.00774
Bollinger K.S., Derksen D.V. Demographic characteristics of molting black brant near Teshekpuk Lake, Alaska // J. Field Ornithol. 1996. V. 67. P. 141–158.
Walsh N.E., Fancy S.G., McCabe T.R. et al. Habitat used by the Porcupine Caribou Herd during predicted insect harassment // J. Wildlife Manage. 1992. V. 56. P. 465–473.
Кирющенко С.П. О трофических взаимоотношениях леммингов и растительности в тундровых экосистемах // Экология млекопитающих тундры и редколесья Северо-Восточной Сибири. Владивосток: ДВНЦ АН СССР, 1985. С. 96–108.
Разумовский С.М. Закономерности динамики биоценозов. М.: Наука, 1981. 231 с.
Koffijberg K., van Winden E., Clausen P. The Netherlands as a winter refuge for light-bellied brent geese from 1991–2011 // Wildfowl. 2013. V. 3. P. 40–57.
Дополнительные материалы отсутствуют.