

Экология, 2022, № 3, стр. 202-210
Временны́е масштабы динамики сообществ грызунов и их соотношение с уровнями пространственных различий фаун
Н. Г. Смирнов a, *, Ю. Э. Кропачева a
a Институт экологии растений и животных УрО РАН
620144 Екатеринбург, ул. 8 Марта, 202, Россия
* E-mail: nsmirnov@ipae.uran.ru
Поступила в редакцию 30.09.2021
После доработки 11.11.2021
Принята к публикации 06.12.2021
- EDN: MDFXEL
- DOI: 10.31857/S0367059722030106
Аннотация
Работа выполнена на материалах из современных и древних (голоценовых и позднеплейстоценовых) местонахождений орнитогенных скоплений остатков грызунов Урала. Разработаны и апробированы критерии отнесения временно́й динамики сообществ грызунов к тем или иным масштабам на основе их сопоставления с уровнями пространственных различий современных фаун грызунов из местонахождений, находящихся на территориях разного ботанико-географического ранга – округов, подзон и зон. На основе распределения долей остатков грызунов (%) в разновременных местонахождениях Среднего Урала рассчитаны парные коэффициенты экологического и таксономического сходства. Показана аналогия пространственных различий и масштабов временно́й динамики (зональные различия – эволюционный масштаб, подзональные различия – исторический масштаб, внутриокружные различия – актуальный масштаб).
Цель данной работы – разработка и апробация приемов оценки временны́х масштабов динамики состава и структуры сообществ грызунов в позднем плейстоцене и голоцене на основе сопоставления современных референтных и тестируемых фаун разного возраста.
При изучении динамики фаун грызунов основным материалом служат разновозрастные скопления субфоссильных и ископаемых костных остатков, накопленные в результате кормодобывающей деятельности сов-миофагов. Такие скопления на Урале чаще всего обнаруживаются в гротах и нишах на скалах, где поколение за поколением оставляют свои погадки филины. Хорошо известно [1], что в кормовой спектр филинов входят практически все виды мелких млекопитающих, обитающие в пределах их охотничьих территорий. При обильных сборах костных остатков жертв это обеспечивает полную характеристику видового состава региональной фауны грызунов. Структура населения в отличие от состава фауны отражается в добыче с определенным искажением. Жертвы подразделяются на основные, альтернативные и сопутствующие в соответствии с их долей в добыче хищника [1, 2]. Наибольшая доля основных жертв обусловлена не только большой численностью в природе, широким распространением соответствующих местообитаний, но и их доступностью. Эти характеристики приходится учитывать при реконструкции структуры локальных фаун на основе изучения костных остатков, извлекаемых из орнитогенных местонахождений. В основе такой работы лежит список видов жертв с указанием количества остатков идентифицированных элементов скелета. Согласно этим данным вычисляются количество и доля особей соответствующих видов.
Попытки классификации временны́х преобразований фаун путем отнесения их к одному из трех масштабов – актуальному, историческому или эволюционному – предпринимались и ранее, но на ограниченном материале и без четко сформулированных методических приемов [3]. Настоящая работа частично заполняет эти пробелы и позволяет продвинуться к поставленной цели. Нами сохранены самые общие критерии разделения динамики на три масштаба, в основе которых находятся степень обратимости и глубина преобразования фаун, максимальной из которых является смена зонального типа, относимая к эволюционному масштабу. Преобразования сообществ эволюционного масштаба, как правило, сопровождаются трансформациями морфологических характеристик отдельных видов, вымиранием или появлением новых. Минимальная динамика фиксируется по обратимым колебаниям структуры фаун в пределах актуального масштаба. Промежуточный масштаб именуется историческим и фиксируется по частично обратимым сдвигам структуры и состава фаун, который не достигает уровня смены зонального типа. Исследование масштабов временно́й динамики в данной работе самым тесным образом сопрягается с пространственными различиями современных фаун из местонахождений, относящихся к различным ботанико-географическим подразделениям.
МАТЕРИАЛ И МЕТОДЫ
Пространственные различия современных фаун оценены по материалам из местонахождений, относящихся к различным ботанико-географическим подразделениям Урала (округа, подзоны, зоны) [4]. Расположение использованных местонахождений и их принадлежность к ботанико-географическому подразделению представлены на рис. 1. В лесостепной зоне Зауралья и южной оконечности Уральских гор расположены местонахождения Сухарыш [5] и Верблюжка [6]. Остальные местонахождения относятся к таежной зоне. В подзоне южной тайги Среднего Урала расположены местонахождения Филин, Старик [7], Ёква [8] (Чусовской округ), в подзоне подтаежных лесов на границе с Красноуфимской лесостепью находятся гроты Сухореченский [9] и Нижнеиргинский, слой 2 [10]. Фауны из районов средней (Пихтовка, Шежим) и северной (Кыбла) тайги охарактеризованы по данным из местонахождений в верховьях рек Печора и Илыч [11, 12].
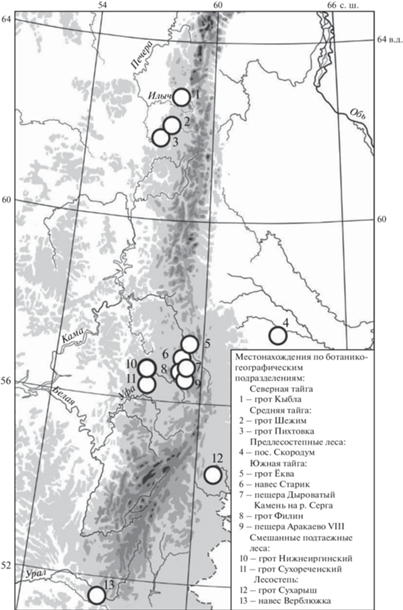
Отнесение данных местонахождений к современности основано на том, что они происходят из погадок ныне жилых гнезд филинов или заселялись в ХХ в., как об этом свидетельствуют очевидцы (гроты Филин, Старик, Ёква, Пихтовка, Шежим, Кыбла, Верблюжка). Материалы из второго слоя Нижнеиргинского грота имеют радиоуглеродную дату 705 ± 20 лет (СПБ № 571) [10] и также рассматриваются в ряду современных местонахождений.
Материалы по временно́й динамике фаун грызунов происходят из позднеплейстоценовых и голоценовых местонахождений Среднего Урала, имеющих радиоуглеродные датировки, а также материалы из современных локалитетов, которым был присвоен статус референтных. Приводятся калиброванные значения возраста древних местонахождений:
Подзона южной тайги: пещера Аракаево VIII – 19144 ± 695 лет (ИМЭЖ-230) [9]; грот Дыроватый Камень на р. Серга, слой 11 – 10 577 ± 237 лет (ИЭМЭЖ-1072) [9]; грот Дыроватый Камень на р. Серга, сл. 8 – 7367 ± 44 (ИЭМЭЖ-1365) [9] (см. рис. 1).
Подзона смешанных подтаежных лесов: грот Нижнеиргинский, сл. 2 – 705 ± 20 лет (СПБ-571); грот Нижнеиргинский, сл. 3, гор. 5 – 3104 ± 116 лет (СПБ-809) [10].
Для иллюстрации наименьшего масштаба временно́й динамики фаун из орнитогенных сборов костных остатков грызунов использованы межгодовые различия в питании бородатой неясыти. Погадки птиц собраны на компактной территории в период гнездования птиц в 2017–2020 гг. в окр. пос. Скородум (предлесостепные леса южной тайги) [13].
Процедуре описания временно́й динамики состава и структуры фаун предшествует этап извлечения костных остатков из породы и определение их принадлежности к той или иной части скелета и таксону. На основе этих данных для каждого вскрытого горизонта и слоя формируются таблицы, где для обнаруженных таксонов указывается количество остатков и особей. Эти сведения служат основой для характеристики состава фауны и соотношения долей разных компонентов.
При описании и анализе динамики в данной работе был использован метод парных сравнений. Для начала цепочки сравнений тестируемых древних фаун, характеризующих разные временны́е фазы региональной динамики, выбирается ближайшее по территориальному расположению местонахождение остатков современных грызунов, фауне из которого можно присвоить ранг референтной. Изучаемые фауны упорядочиваются в хронологическом порядке и проводится процедура парных сравнений. В этом ряду референтная фауна служит эталоном современного зонального состояния. Тестируемые древние фауны в первую очередь сравниваются с ней, а также между собой.
Основное качество референтной фауны – полноценное отображение видового состава обитающих в регионе грызунов. Ранее было показано [14], что в орнитогенных остеологичеких сборах обнаруживаются остатки даже самых редких грызунов, если количество особей превосходит 150, а число щёчных зубов порядка одной тысячи. Еще одно требование к сравниваемым местонахождениям – принадлежность к одному и тому же типу по агенту накопления костных остатков. Этот показатель оценивается по комплексу признаков: расположению местонахождений, степени и типу сохранности остатков. В некоторых случаях важнейшей характеристикой служит состав жертв (птицы, мелкие млекопитающие, рыбы или другие группы животных) [15]. Нами использованы местонахождения в карстовых полостях, где агентом накопления был филин. Единственным исключением служат сборы погадок у гнезд бородатой неясыти у пос. Скородум.
Сравнение древних и референтных фаун дает количественную оценку различий в хронологическом аспекте. Чтобы отнести эти различия к определенному масштабу динамики, они сопоставлены с пространственными различиями фаун из современных местонахождений, расположенных в разных ботанико-географических подразделениях Урала.
В качестве критерия сходства сравниваемых фаун использован показатель сходства Л.А. Животовского [16]:$~$
где pi и qi – частоты i-го по номеру вида или группы видов в сравниваемых выборках, m – число общих видов.Значимость отличия R от 1 определяется критерием идентичности:
где N – объемы 1-й и 2-й выборок. Если величина I больше табличного значения χ2 с m – 1 степенями свободы, то на выбранном уровне значимости выборки отличаются друг от друга.Показатель R принимает значения от 0 при полном несходстве до 1 при полном сходстве. Когда в основу расчетов берутся доли особей таксонов в сравниваемых фаунах, то он является показателем таксономического сходства и обозначается как Rtax. Если виды объединяются по принадлежности к экологическим группам, то показатель сравниваемых пар фаун именуется экологическим (Reco). Группы, по которым распределяются виды для расчета Reco, формируются в соответствии с распространением в современных биомах и занимаемых там экологических нишах.
Поскольку основной задачей описания эволюционного и исторического масштабов динамики фаун является оценка глубины их преобразования, то в этой процедуре определяющим элементом служит наземная (плакорная) часть населения, так как именно по ней оцениваются зональные характеристики [17]. Остальные элементы – виды из околоводной, агрополевой, синантропной групп – могут образовывать основу пищевого спектра хищника, но для поставленной задачи бесполезны и должны исключаться из анализа. Деление фаун на экологические категории начинается с подразделения на естественную и синантропную части. К последней относятся серая крыса и домовая мышь. В отдельную агрополевую категорию включаются обыкновенная полевка и ее виды-двойники. Виды околоводных местообитаний (ондатра, водяная полевка, полевка-экономка) невозможно отнести ни к одной зональной группе, и они вместе с вышеперечисленными синантропными и агрополевыми видами исключаются из анализа зональной характеристики фаун.
При расчетах экологических показателей сходства Reco зональные виды группируются в следующие категории: 1) таежные (белка, бурундук, летяга, лесные полевки, лесной лемминг, лесная мышовка); 2) лесолуговые (темная полевка, малая лесная мышь); 3) луговые (обыкновенный хомяк и полевая мышь; 4) лугово-степной вид (узкочерепная полевка); 5) степные (слепушонка, степная пеструшка, серый и эверсманов хомячки, большой тушканчик, степная пищуха); 6) тундровые (копытный, сибирский лемминги, полевка Миддендорфа). Частоты встречаемости особей соответствующих видов суммируются в пределах каждой категории и по этим значениям производятся вычисления. Показатели таксономического сходства Rtax высчитываются на основе частот встречаемости особей каждого вида, общего для сравниваемых фаун, из тех, что вошли в список зональных видов.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Характеристики временно́й динамики сопоставлены с пространственными различиями фаун из районов, находящихся в разных ботанико-географических подразделениях (окружного, подзонального, зонального уровней).
Минимальное пространственное сходство наблюдалось при межзональном сравнении (рис. 2). Так, 16 пар сравнений местонахождений, расположенных в таежной зоне Северного и Среднего Урала, имеют показатели экологического сходства с фаунами из лесостепной зоны Зауралья и южной оконечности Урала – от 0.28 до 0.75, в среднем 0.52 (табл. 1).
Рис. 2.
Средние значения показателя R при разных типах пространственных сравнений современных сообществ грызунов: 1 – экологического сходства (Reco); 2 – таксономического сходства (Rtax).

Таблица 1.
Показатели экологического (Reco, левая половина таблицы) и таксономического (Rtax, правая половина таблицы) сходства фаун грызунов из местонахождений, находящихся на территориях разного ботанико-географического ранга (I; df)
| Зона | Тайга | Лесостепь | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Подзона | Северная тайга | Средняя тайга | Смешанные подтаежные леса | Южная тайга | |||||||||
| Округ | Саранинский | Чусовской | |||||||||||
| Местона- хождение |
Кыбла | Шежим | Пихтовка | Нижне- иргинский, слой 2 |
Сухоре- ченский |
Филин | Старик | Ёква | Верблюжка | Сухарыш | |||
| Тайга | Северная тайга |
Кыбла | 0.91 (27.09; 4) | 0.84 (39.11; 5) | 0.53 (216.88; 5) | 0.47 (374.92; 4) | 0.8 (59.1; 7) | 0.78 (51.42; 4) | 0.91 (63.07; 6) | 0.3 (196.08; 4) | 0.45 (171.90; 4) | ||
| Средняя тайга |
Шежим | 0.97 (9.99; 1) | 0.94 (12.06; 4) | 0.42 (174.74; 3) | 0.36 (253.88; 3) | 0.59 (89.2; 4) | 0.56 (82.54; 3) | 0.83 (63.72; 4) | 0.28 (164.48; 1) | 0.44 (140.34; 4) | |||
| Пихтовка | 0.91 (23.72; 1) | 0.98 (3.04; 1) | 0.39 (157.02; 4) | 0.34 (211.81; 5) | 0.52 (93.16; 5) | 0.46 (92.26; 3) | 0.83 (53.41; 1) | 0.3 (425.40; 4) | 0.22 (632.62; 4) | ||||
| Смешанные подтаежные леса |
Сара- нинский |
Нижне- иргинский, сл. 2 |
0.73 (125.36; 1) | 0.73 (81.46; 1) | 0.71 (75.94; 1) | 0.86 (109.72; 7) | 0.88 (36.23; 7) | 0.85 (37.33; 6) | 0.74 (194.13; 9) | 0.53 (198.69; 5) | 0.46 (279.64; 5) | ||
| Сухоре- ченский |
0.66 (238.98; 1) | 0.58 (164.96; 1) | 0.51 (159.16; 1) | 0.94 (48.6; 2) | 0.72 (109.06; 6) | 0.7 (90.0; 4) | 0.6 (668.13; 7) | 0.54 (290.58; 3) | 0.35 (551.27; 3) | ||||
| Южная тайга |
Чусов- ской |
Филин | 0.89 (30.9; 1) | 0.87 (28.51; 1) | 0.82 (35.05; 1) | 0.95 (14.29; 2) | 0.91 (37.18; 2) | 0.98 (4.14; 6) | 0.89 (41.91; 8) | 0.14 (239.47; 1) | 0.06 (294.87; 0) | ||
| Старик | 0.91 (21.26; 1) | 0.88 (23.24; 1) | 0.82 (30.68; 1) | 0.94 (15.32; 2) | 0.9 (30.03; 2) | 0.99 (0.28; 2) | 0.84 (47.09; 6) | 0.11 (214.96; 1) | 0.03 (257.38; 0) | ||||
| Ёква | 0.95 (33.28; 1) | 0.97 (11.87; 1) | 0.94 (17.63; 1) | 0.87 (92.95; 2) | 0.76 (409.17; 2) | 0.96 (14.42; 2) | 0.96 (11.12; 2) | 0.18 (330.75; 2) | 0.1 (436.27; 0) | ||||
| Лесостепь | Верблюжка | 0.50 (139.1; 2) | 0.49 (117.85; 2) | 0.41 (358.53; 2) | 0.65 (150.72; 3) | 0.75 (270.57; 2) | 0.29 (198.61; 1) | 0.28 (174.15; 1) | 0.29 (288.14; 1) | 0.58 (188.88; 6) | |||
| Сухарыш | 0.62 (118.2; 2) | 0.63 (94.64; 2) | 0.61 (313.14; 2) | 0.62 (196.7; 3) | 0.55 (386.03; 2) | 0.57 (135.83; 1) | 0.53 (124.21; 1) | 0.58 (167.99; 1) | 0,76 (108.32; 4) | ||||
Показатели таксономического сходства при том же сравнении оказались еще меньше (см. рис. 2) – от 0.03 до 0.54 при среднем 0.28, что отражает кардинальное различие в видовом составе. Это дает основание при сравнениях разновременных фаун, получив аналогичные величины показателей сходства, считать такие фауны принадлежавшими к разным зональным типам. Именно такой уровень сходства (табл. 2) дает сравнение современной референтной таежной фауны из грота Ёква с позднеплейстоценовой фауной из пещеры Аракаево VIII (их различие в возрасте составляет около 19 тыс. лет). Масштаб их динамики демонстрируют данные по соотношению долей остатков грызунов из разных экологических групп (табл. 3). Так, доля лугово-степных, степных и тундровых видов в Аракаево VIII составляет 94%, тогда как в Ёкве виды этих групп отсутствуют. Не столь радикальную динамику, но вполне сопоставимую с межзональными различиями демонстрирует сравнение таежной фауны из Ёквы с фауной из слоя 11 грота Дыроватый Камень на р. Серга (разница в возрасте около 9 тыс. лет). Отсутствующие в таежной фауне Ёквы виды из лугово-степной, степной и тундровой групп в слое 11 Дыроватого Камня составляют по доле остатков 68%. Эти примеры, безусловно, убеждают, что аналогия межзональных пространственных различий эволюционному масштабу временно́й динамики вполне правомочна. Графическое представление показателей сходства пар фаун разного временно́го масштаба на основе табл. 2 представлено на рис. 3.
Таблица 2.
Парные коэффициенты экологического и таксономического сходства фаун, демонстрирующие аналогию пространственных различий и масштабы временно́й динамики
| Сравниваемые местонахождения | Reco (I; df) | Rtax (I; df) | Разница в возрасте местонахождений, лет |
|---|---|---|---|
| Аналог межзонального различия (эволюционный масштаб временнóй динамики) | |||
| Ёква – Аракаево VIII | 0.24 (2032.51; 2) | 0.18 (2213.32; 4) | 19 144 |
| Ёква – Дыроватый Камень, сл. 11 | 0.5 (1496.21; 2) | 0.41 (1752.16; 6) | 10 577 |
| Аналог подзонального различия (исторический масштаб временнóй динамики) | |||
| Ёква – Дыроватый Камень, сл. 8 | 0.85 (174.3; 2) | 0.59 (481.16; 4) | 7367 |
| Дыроватый Камень, сл. 11 – Аракаево VIII | 0.82 (3442.59; 5) | 0.77 (4390.72; 10) | 8567 |
| Дыроватый Камень, сл. 8 – сл. 11 | 0.79 (393.81; 5) | 0.77 (442.16; 8) | 3210 |
| Аналог различий внутри одного округа (актуальный масштаб временно́й динамики) | |||
| Нижнеиргинский, сл. 2 – сл. 3 | 0.99 (6.82; 2) | 0.94 (32.27; 9) | 2399 |
| Скородум, 2017–2020 | 0.96 (19.97; 2) | 0.95 (27.13; 3) | 3 |
| Скородум, 2019–2020 | 0.97 (9.47; 2) | 0.88 (32.88; 3) | 1 |
| Скородум, 2017–2019 | 0.99 (0.56; 2) | 0.93 (21.43; 4) | 2 |
Таблица 3.
Распределение долей остатков грызунов по экологическим группам в разновременных (от современности до позднего плейстоцена) местонахождениях Среднего Урала, %
| Местонахождение и его возраст | Экологические группы видов | ||||||
|---|---|---|---|---|---|---|---|
| таежные | лесолуговые | луговые | лугово-степные | степные | тундровые | ||
| Грот Ёква (ХХ в.) | 76 | 18 | 6 | 0 | 0 | 0 | |
| Грот Нижнеиргинский | сл. 3, гор. 5 (3104 ± 116 лет) | 46 | 5 | 48 | 0 | 0 | 0 |
| сл. 2 (705 ± 20 лет) | 42 | 12 | 45 | 0 | 1 | 0 | |
| Пос. Скородум (сборы свежих погадок) | 2017 г. | 11 | 48 | 0 | 41 | 0 | 0 |
| 2019 г. | 13 | 44 | 0 | 42 | 0 | 0 | |
| 2020 г. | 22 | 58 | 0 | 20 | 0 | 0 | |
| Пещера Дыроватый Камень | сл. 8 (7367 ± 44) | 64 | 8 | 2 | 22 | <1 | 4 |
| сл. 11 (10577 ± 237 лет) | 13 | 19 | <1 | 60 | 8 | <1 | |
| Аракаево VIII (19144 ± 695 лет) | 5 | 1 | <1 | 46 | 30 | 18 | |
Рис. 3.
Показатели сходства (R) пар фаун разного масштаба временно́й динамики: а – экологического сходства (Reco); б – таксономического сходства (Rtax).

Временна́я динамика эволюционного масштаба, как и межзональное пространственное сопоставление, нуждаются в подразделении на градации по степени выраженности. Широкий диапазон варьирования (от близкого к 0 до 0.75) имеет смысл делить по крайней мере на три части (0–0.24; 0.25–0.49; 0.50–0.75). Соответствующие масштабы следует именовать эволюционным максимального, среднего и минимального уровней. Эти уровни отражают не столько временно́й диапазон, сколько глубину преобразования структуры фаун [3].
Больший уровень сходства по сравнению с межзональным демонстрируют фауны, которые расположены в пределах одной зоны, но в разных подзонах. Так, при сравнении 23 пар современных фаун, находящихся в северной, средней и южной тайге, экологическое сходство варьировало от 0.51 до 0.97 при среднем 0.83, а таксономическое было равно от 0.34 до 0.91 при среднем 0.65. Такое пространственное сходство фаун на подзональном уровне позволяет искать в подобном интервале соответствующие аналоги среди сравнений фаун разного возраста. Именно в эти интервалы значений попадает сходство следующих разновременных фаун: Ёква – Дыроватый Камень на р. Серга, сл. 8; Дыроватый Камень на р. Серга, сл. 8 – Дыроватый Камень на р. Серга, сл. 11; Дыроватый Камень на р. Серга, сл. 11 – Аракаево VIII (см. табл. 2). Эта временна́я динамика относится к историческому масштабу. Состав фаун при такой динамике меняется не кардинально, а за счет редких и очень редких видов при постоянстве доминантов [3].
Третий, самый тесный, уровень пространственного сходства демонстрируют фауны, находящиеся внутри одних ботанико-географических округов. Для сравнений удалось подобрать 5 пар фаун. Показатели экологического сходства варьировали от 0.94 до 0.99 (среднее 0.97), а таксономического – от 0.84 до 0.98 (среднее 0.90). Временны́м аналогом такой пространственной динамики может служить сходство фаун с показателями, близкими к единице, которые демонстрируют динамику в актуальном масштабе. Примером могут служить фауны горизонтов 2 и 3 грота Нижнеиргинский, возраст которых различается всего на 1780 лет, а показатели экологического и таксономического сходства соответственно равны 0.99 и 0.94, т.е. близки к единице (см. табл. 2). Такие значения отражают обратимые сдвиги относительной численности некоторых видов при постоянстве основных зональных характеристик [3]. Не менее убедительным примером динамики в актуальном масштабе служит сходство фаун, выявленное при сравнении состава погадок бородатых неясытей, собранных на компактной территории гнездования в Ирбитском районе Свердловской области в 2017–2020 гг.: показатели экологического сходства варьировали от 0.96 до 0.99 при среднем 0.97, а таксономического – от 0.88 до 0.95 при среднем 0.92 (см. табл. 2).
ЗАКЛЮЧЕНИЕ
Полноценным материалом для изучения многолетней динамики фауны грызунов являются субфоссильные и ископаемые остатки мелких млекопитающих из их массовых скоплений в скальных гротах и навесах, служивших для гнезд и присад филинов. Послойные раскопки и датирование этих скоплений позволяют сравнивать видовой состав и структуру жертв хищников в последовательные временны́е отрезки. Количественной мерой такого сравнения в данной работе послужили парные показатели экологического и таксономического сходства (Reco и Rtax). Однако в описаниях и сравнениях использованы не все обнаруженные виды, а только те, которые указывают на зональную характеристику фауны. Для оценки экологического сходства последние группируются в следующие категории: таежные виды, лесолуговые, луговые, лугово-степные, степные, тундровые. Не использованы в анализе виды околоводных и агрополевых местообитаний, а также синантропные.
Для отнесения временно́й динамики к тому или иному масштабу основным критерием служит аналог степени различия (сходства) фаун в пространственном градиенте.
Динамика наименьшего временно́го масштаба, именуемого актуальным, аналогична степени сходства пар сравниваемых фаун, находящиеся внутри одних ботанико-географических округов. Такое сходство настолько велико, что сравнение долей экологических категорий близко или равно единице, как и таксономическое сходство.
Эволюционный масштаб временно́й динамики сопоставляется с пространственным сравнением фаун, находящихся в разных ботанико-географических зонах. Такая динамика, как и межзональное сопоставление, подразделяются на три градации по степени выраженности.
Исторический масштаб временно́й динамики фаун занимает промежуточное положение между актуальным и эволюционным. Его аналогом в пространственной дифференцировке служит сходство фаун из разных ботанико-географических подзон.
Работа выполнена в рамках государственного задания Института экологии растений и животных УрО РАН при частичной поддержке РФФИ (проекты № 19-04-01008 и 19-04-00507).
Авторы выражают благодарность Е.П. Изварину за содействие в выполнении работы и рецензенту за конструктивную критику и советы по улучшению статьи.
Настоящая работа не содержит исследований с участием людей или животных в качестве объектов исследования.
Авторы подтверждают отсутствие конфликта интересов.
Список литературы
Penteriani V., del Mar Delgado M. The eagle owl. London: Bloomsbury Publishing, 2019. 384 p.
Смирнов Н.Г., Кропачева Ю.Э. Основные и сопутствующие жертвы филина (Bubo bubo) в задачах исторической экологии // Экология. 2019. № 5. С. 387–391.
Смирнов Н.Г. Динамика видов и их комплексов как предмет исследований исторической экологии // Экология. 2006. № 6. С. 452–457.
Эндемичные растения Урала во флоре Свердловской области / П.В. Куликов, Н.В. Золотарева, Е.Н. Подгаевская; науч. ред. Мухин В.А. Екатеринбург: “Гощицкий”, 2013. 612 с.
Смирнов Н.Г. Проблемы исторической экологии млекопитающих Северной Евразии // Вековая динамика биогеоценозов: Чтения памяти академика В.Н. Сукачева. М.: Наука, 1992. С. 17–35.
Kuzmina E.A., Smirnov N.G., Ulitko A.I. New data on Late Pleistocene–Holocene small mammal communities from the Ural–Sakmara interfluve, Southern Urals // Quaternary international. 2016. V. 420. P. 56–64.
Садыкова Н.О. Сообщество мелких млекопитающих долины реки Серги в позднем голоцене // Экология в меняющемся мире: Мат-лы конф. молодых ученых. Екатеринбург, 2006. С. 208–210.
Кропачева Ю.Э., Улитко А.И., Шершнев М.Ю. и др. Характеристика питания филина по материалам из грота Ёква в природном парке “Река Чусовая” // Научные исследования на ООПТ Урала: Мат-лы межрегион. конф. Екатеринбург, 2021. С. 67–70.
Смирнов Н.Г. Мелкие млекопитающие Среднего Урала в позднем плейстоцене и голоцене. Екатеринбург: Наука, 1993. 64 с.
Izvarin E.P., Ulitko A.I., Nekrasov A.E. Palaeontological description of Nizhneirginsky Grotto Upper Holocene sediments (Ufa Plateau, Fore-Urals) with taphonomic and palaeoenvironmental remarks based on bird and small-mammal assemblages // Quaternary International. 2020. V. 546. P. 160–169.
Садыкова Н.О., Смирнов Н.Г. Формирование локальных и элементарных фаун в зоогенных отложениях Печоро-Илычского заповедника / Труды Печоро-Илычского государственного заповедника. 2005. Вып. 14. С. 152–158.
Садыкова Н.О. Подходы к изучению вековой и многолетней динамики сообществ мелких млекопитающих на палеонтологическом материале (на примере Северного Урала) // Экология от Арктики до Антарктики: Мат-лы конф. молодых ученых. Екатеринбург, 2007. С. 277–282.
Кропачева Ю.Э., Смирнов Н.Г., Зыков С.В. и др. Питание бородатой неясыти (Strix nebulosa) при разной численности жертв в период гнездования // Экология. 2019. № 1. С. 40–46.
Смирнов Н.Г., Маркова А.К. Методические вопросы оценки таксономического разнообразия млекопитающих на основе остеологических сборов // Материалы и исследования по истории современной фауны Урала. Екатеринбург, 1996. С. 3–16.
Andrews P. Owls, caves and fossils: predation, preservation and accumulation of small mammal bones in caves, with an analysis of the Pleistocene cave faunas from Westbury-sub-Mendip, Somerset, UK. Chicago: University of Chicago Press, 1990. 231 p.
Животовский Л.А. Показатель сходства популяций по полиморфным признакам // Журн. общ. биол. 1979. Т. 40. № 4. С. 587–601.
Чернов Ю.И. Природная зональность и животный мир суши. М.: Мысль, 1975. 222 с.
Дополнительные материалы отсутствуют.