

Экология, 2022, № 3, стр. 161-170
У микоризных чужеродных растений Белорусского Полесья понижена интенсивность формирования арбускул
Д. В. Веселкин a, *, А. А. Бетехтина b, А. П. Гусев c
a Институт экологии растений и животных УрО РАН
620144 Екатеринбург, ул. 8 Марта, 202, Россия
b Уральский федеральный университет им. первого Президента России Б.Н. Ельцина
620083 Екатеринбург, просп. Ленина, 51, Россия
c Гомельский государственный университет им. Ф. Скорины
246019 Гомель, ул. Советская, 104, Республика Беларусь
* E-mail: veselkin_dv@ipae.uran.ru
Поступила в редакцию 26.01.2022
После доработки 29.01.2022
Принята к публикации 29.01.2022
- EDN: UPJALW
- DOI: 10.31857/S036705972203012X
Аннотация
Проверяли гипотезу о разной способности чужеродных и аборигенных растений к образованию арбускулярной микоризы. Исследования выполнены в Белорусском Полесье (Восточная Европа) в зоне широколиственных лесов с умеренно континентальным климатом. Микроскопически определили интенсивность формирования разных структур арбускулярной микоризы (арбускул, везикул, гиф), а также корневых волосков у 19 чужеродных и 25 аборигенных видов растений, собранных в 6 разных местообитаниях. В каждом местообитании собирали и чужеродные, и аборигенные растения. У микоризных чужеродных растений арбускулы формировались реже (28 ± 6%), чем у аборигенных (48 ± 5%). Корневые волоски у чужеродных видов закладывались чаще (43 ± 6%), чем у аборигенных (32 ± 4%). И у аборигенных, и у чужеродных растений микориза активнее формировалась в нарушенных местообитаниях, но взаимодействие факторов “степень натурализации” и “степень нарушенности местообитаний” не установлено. Таким образом, у чужеродных растений Белорусского Полесья арбускулярная микориза функционирует менее эффективно, чем у аборигенных.
К свойствам, обеспечивающим способность растений длительно удерживаться в занятых местообитаниях, наряду с обычно изучаемыми морфофизиологическими параметрами вегетативных органов [1] относятся микоризные взаимодействия. Тип и теснота микоризных связей коррелируют с такими функциональными свойствами, как скорость захвата ресурсов, способность снизить их концентрацию в почве до недоступных конкурентам уровней, быстрый рост, устойчивость к стрессам и нарушениям [1–5]. Самые распространенные и мало специализированные микоризы – арбускулярные (AM) [6, 7]. Они очень важны для определения структуры и динамики сообществ [3, 6].
Степень специализации к взаимодействию с симбионтами может иметь большое значение для расселения и проникновения чужеродных растений в новые местообитания [8, 9]. Теснота взаимодействия чужеродных растений с микоризными грибами связана с площадью ареала растений: факультативно-микоризные виды, как правило, населяют больший ареал, чем немикоризные и облигатно-микоризные пришлые растения [10]. Обычно обсуждаются два возможных паттерна тесноты микоризных связей у чужеродных растений: или меньшая, чем у местных растений, или равная с ними способность к формированию микоризы [11]. Нередко делается вывод о меньшей зависимости чужеродных растений от микоризы [12–15], но чаще констатируется, что чужеродные растения примерно в равной степени зависимы от микоризных грибов, как и местные [10, 11, 13, 14, 16, 17]. Анализ микоризности чужеродных видов из глобальной базы Global Naturalized Alien Flora (GloNAF), объединяющей сведения о 840 материковых и островных регионах, выявил более высокую вероятность натурализации у микоризных растений, чем у немикоризных [9].
Гипотеза “деградировавшего или нарушенного мутуализма” [18–20] предполагает возможность каскадного изменения сообществ вследствие особенностей взаимодействия чужеродных растений с местными микоризными грибами. Если чужеродные растения менее зависят от микоризных грибов, чем местные, это изменяет местные грибные сообщества и облегчает проникновение неместных немикоризных растений, препятствуя восстановлению местных сообществ. Поэтому важно точно и предметно, на основании специальных эмпирических проверок, представлять, как чужеродные растения взаимодействуют с местными микоризными грибами в разных регионах и экосистемах. Это тем более необходимо, что существует выраженная географическая изменчивость распространения и значимости разных типов микоризы [6]: закономерности, справедливые для одних регионов и условий, могут не воспроизводиться в другом окружении. Поэтому необходимы накопление и систематизация сведений о микоризных взаимодействиях чужеродных растений в сравнении с аборигенными.
Цель настоящей работы – проверка гипотезы о разной способности чужеродных и аборигенных растений Белорусского Полесья к образованию арбускулярной микоризы.
МАТЕРИАЛ И МЕТОДЫ
Район. Сбор растений или образцов их корневых систем выполнен вблизи г. Гомель (Восточная Европа, Республика Беларусь, 52°22′59″–52°29′28″ с.ш., 30°55′11″–30°59′43″ в.д.). Климат умеренно континентальный, средняя температура самого холодного месяца (январь) –4.4°C, самого теплого месяца (июль) +20.5°C. Среднегодовая температура +8.3°C. Годовая сумма температур выше +10°C равна 2650°C. Период со среднесуточными температурами выше 0°C длится 263 дня. Количество осадков 635 мм/год (годовой максимум осадков приходится на июнь–июль). Коэффициент увлажнения 1.1. По климатическим показателям территория относится к суббореальным гумидным ландшафтам. Зональная растительность представлена сосновыми, широколиственными, широколиственно-сосновыми, коренными мелколиственными лесами, а также пойменными лугами и болотами. Местообитания и растительные сообщества района длительно и интенсивно преобразовываются под влиянием хозяйственной деятельности человека. Большинство естественных местообитаний антропогенно трансформированы, а растительные сообщества представляют собой разные этапы естественного восстановления.
В районе исследований инвазионные процессы идут активно [21, 22]. Доли чужеродных видов составляют до 15–40% общих флористических списков в разных типах нелесных ландшафтов и до 2–30% – в лесных. Активнее всего в естественные сообщества внедряются Erigeron canadensis, Oenothera biennis и Acer negundo. Также активны Amaranthus retroflexus, Robinia pseudoacacia, Galinsoga parviflora, Oxalis stricta, Erigeron annuus, Echinocystis lobata, Solidago canadensis.
Местообитания. Сбор чужеродных и аборигенных растений выполнен в начале августа 2014 г. в 6 местообитаниях: в 2 мало нарушенных местообитаниях представлены зональные лесные сообщества, в 4 – травяные сообщества, разное время восстанавливающиеся после нарушений:
1. Лес 1; древесный ярус – Betula pendula, Populus tremula, Pinus sylvestris (возраст 30–40 лет); напочвенный покров – Calamagrostis epigejos, Melampyrum sylvaticum, Solidago virgaurea, Pteridium aquilinum; почвы дерново-слабоподзолистые супесчаные.
2. Лес 2; древесный ярус – P. sylvestris (возраст 50–60 лет); напочвенный покров – Pleurozium schreberi, Festuca ovina, M. sylvaticum, C. epigejos, Helichrysum arenarium, S. virgaurea; почвы дерново-слабоподзолистые песчаные.
3. Просека, зарастающая 10–15 лет; подрост B. pendula, P. sylvestris; напочвенный покров – C. epigejos, Artemisia alba, Berteroa incana, Echium vulgare, O. biennis, F. ovina, E. canadensis, Poa pratensis, Achillea millefolium; почвы песчаные нарушенные.
4. Залежь после пашни, зарастающая около 15 лет; подрост A. negundo, B. pendula; напочвенный покров – Tanacetum vulgare, Artemisia vulgaris, Cirsium arvense, A. millefolium, Dactylis glomerata, C. epigejos, Elymus repens, Bromus inermis; почвы дерново-палевоподзолистые суглинистые.
5. Склон карьера по добыче песка, зарастающий 12 лет; подрост древесных – P. sylvestris, Salix sp.; напочвенный покров – C. epigejos, O. biennis, E. canadensis, H. arenarium, Corynephorus canescens, Solidago canadensis; грунт песчаный, уплотненный.
6. Песчаный отвал, зарастающий около 5 лет; напочвенный покров – C. canescens, Bromus tectorum, Equisetum arvense, E. canadensis, H. arenarium; грунт песчаный, рыхлый.
По признакам силы и давности нарушений и доминирующим жизненным формам растений местообитания естественно объединяются в две группы: 1) со следами сильных нарушений и доминированием травянистых растений – местообитания № 5 и 6; 2) мало нарушенные или продолжительно восстанавливающиеся сообщества с наличием или доминированием древесных растений – местообитания № 1–4.
Сбор образцов, определение формирования микоризы, корневых волосков и темных септированных эндофитов. Собирали образцы максимального числа видов, указываемых для района как чужеродные [22]. Дополнительно использовали сведения о составе чужеродных растений географически близкой Брянской области Российской Федерации [23]. Названия видов приведены по [24]. Образцы каждого вида растений отбирали только в каком-либо одном местообитании, но в каждом местообитании собирали и чужеродные, и аборигенные растения. У каждого вида случайно отбирали и гербаризировали по 5 особей или по 5 образцов корневых систем при невозможности препарировать корневую систему полностью. В каждом образце (у каждой особи) на 15 фрагментах тонких корней последнего и предпоследнего порядков длиной по 1 см определяли микоризную колонизацию. Фрагменты отбирали из корневой системы случайно. Корни подвергали мацерации на водяной бане в KOH 30–60 мин и окрашивали анилиновым синим [25]. На давленных препаратах при увеличении ×200 регистрировали гифы, арбускулы и везикулы грибов АМ. Их присутствие определяли в 5 полях зрения микроскопа на каждом сантиметровом фрагменте корня.
Таким образом, всего было 75 полей зрения для особи и 375 для каждого вида. Характеристики развития AM – интенсивность формирования или доля полей зрения с соответствующими структурами от общего числа полей зрения: а) арбускул (Ar%); б) везикул (Ve%); в) всех структур AM (AM%). В целом результаты учета AM по методике [25] близки с результатами широко используемого метода учета развития AM [26], что специально показано в работе [27]. В тех же полях зрения, в которых определяли интенсивность микоризы, измеряли интенсивность формирования корневых волосков (RH%) и мицелия темных септированных эндофитов (DSE%).
Анализ данных. Степень натурализации растений определяли по [22, 23]: 19 чужеродных и 25 аборигенных видов. Дополнительно как возможные причины изменчивости развития AM анализировали свойства видов и местообитаний: таксономический статус, длительность онтогенеза, степень нарушенности местообитаний. Таксономический статус характеризовали принадлежностью к семейству. Из 44 проанализированных видов 21 был из сем. Asteraceae, 4 – из сем. Poaceae, 3 – из сем. Apiaceae, 12 видов – из семейств, в которых проанализировано по 1–2 вида. Длительность онтогенеза характеризовали двумя градациями: а) 14 малолетних (однолетних, одно-двулетних или двулетних); б) 30 многолетних растений. Степень нарушенности местообитаний характеризовали двумя градациями: а) 32 вида из местообитаний со слабыми и умеренными нарушениями; б) 12 видов из сильно нарушенных местообитаний.
Чтобы оценить, какие факторы сильнее влияли на формирование разных структур в корнях, использовали скорректированный информационный критерий Акаике (AICc) [28] с суммированием AICc-весов моделей (W) в отношении отдельных переменных (ΣW). Значения ΣW интерпретировали как вероятность того, что переменная обладает наибольшей объясняющей ценностью из исследованного набора. Различия признаков между группами видов оценивали с помощью ANOVA с последующим расчетом теста Тьюки для сравнения средних. Использование ANOVA было обосновано, поскольку условия его применимости в большинстве случаев выполнялись. Учетная единица в статистическом анализе – среднее значение признака у одного вида растения. При расчете AICc и в ANOVA использованы значения признаков после арксинус-преобразования, но на рисунках и в тексте использованы нетрансформированные значения. Расчеты выполнены в пакете STATISTICA 8.0 (StatSoft Inc., USA, 1984–2007). При усреднении значений через символ ± приведена стандартная ошибка (SE).
РЕЗУЛЬТАТЫ
На наличие AM исследовали особи 55 видов растений, однако далее анализируются оценки, относящиеся только к 44 видам, у которых зарегистрированы арбускулы (табл. 1). Немикоризными, т.е. не имеющими арбускул и по нашим оценкам, и не указываемые как микоризные в литературе [7, 27], были: Amaranthus retroflexus, Carex spicata, C. hirta, C. nigra, Chenopodium album, Corispermum marschallii, Equisetum arvense, Parthenocissus quinquefolia. Три чужеродных вида (Lupinus polyphyllus, Portulaca oleracea и Robinia pseudoacacia) по нашим данным имели гифы и везикулы, но не имели арбускул. Несмотря на то, что эти виды указаны в сводках как микоризные [7, 27], в работе они рассматриваются как немикоризные.
Таблица 1.
Длительность онтогенеза, нарушенность местообитаний и интенсивность формирования разных корневых структур у 44 видов растений Белорусского Полесья с ненулевой интенсивностью формирования арбускул
| № п.п. |
Вид | Длительность онтогенеза | Нарушенность местообитаний | Ar% | Ve% | AM% | RH% | DSE% |
|---|---|---|---|---|---|---|---|---|
| Аборигенные растения | ||||||||
| 1 | Agrimonia eupatoria | Мног. | Не сильная | 52 | 35 | 67 | 23 | 20 |
| 2 | Apera spica-venti | » | » | 60 | 32 | 85 | 40 | 0 |
| 3 | Artemisia absinthium | » | » | 54 | 24 | 65 | 31 | 0 |
| 4 | Artemisia campestris | » | » | 12 | 36 | 47 | 28 | 0 |
| 5 | Artemisia vulgaris | » | » | 13 | 20 | 25 | 50 | 18 |
| 6 | Ballota nigra | » | » | 58 | 13 | 71 | 70 | 18 |
| 7 | Carduus crispus | » | » | 66 | 26 | 77 | 58 | 18 |
| 8 | Centaurea jacea | » | Сильная | 78 | 38 | 87 | 18 | 6 |
| 9 | Cichorium intybus | » | Не сильная | 52 | 50 | 87 | 15 | 9 |
| 10 | Cirsium arvense | » | » | 53 | 48 | 81 | 55 | 5 |
| 11 | Convolvulus arvensis | » | » | 71 | 63 | 79 | 4 | 16 |
| 12 | Daucus carota | » | » | 52 | 24 | 76 | 29 | 6 |
| 13 | Epilobium angustifolium | » | » | 9 | 27 | 58 | 51 | 2 |
| 14 | Lolium pratense | » | » | 88 | 74 | 100 | 41 | 0 |
| 15 | Helichrysum arenarium | » | » | 40 | 28 | 79 | 23 | 0 |
| 16 | Jasione montana | » | » | 10 | 1 | 42 | 1 | 0 |
| 17 | Koeleria glauca | » | » | 20 | 24 | 57 | 44 | 0 |
| 18 | Pastinaca sativa | » | » | 13 | 0 | 13 | 0 | 2 |
| 19 | Plantago indica | Мал. | Сильная | 76 | 69 | 95 | 0 | 0 |
| 20 | Plantago lanceolata | Мног. | Не сильная | 38 | 10 | 51 | 22 | 0 |
| 21 | Solidago virgaurea | » | » | 96 | 65 | 99 | 3 | 0 |
| 22 | Tanacetum vulgare | » | » | 76 | 46 | 90 | 53 | 0 |
| 23 | Taraxacum sect. Taraxacum | » | » | 77 | 32 | 93 | 35 | 0 |
| 24 | Trifolium arvense | Мал. | » | 16 | 7 | 36 | 45 | 0 |
| 25 | Vicia cracca | Мног. | » | 22 | 31 | 52 | 60 | 15 |
| Чужеродные растения | ||||||||
| 26 | Acer negundo | Мног. | Сильная | 15 | 7 | 30 | 57 | 1 |
| 27 | Ambrosia artemisiifolia | Мал. | » | 20 | 50 | 64 | 20 | 11 |
| 28 | Bidens frondosa | » | Не сильная | 15 | 2 | 19 | 24 | 0 |
| 29 | Cyclachaena xanthiifolia | » | » | 12 | 8 | 30 | 83 | 0 |
| 30 | Echinocystis lobata | » | Сильная | 32 | 43 | 56 | 30 | 0 |
| 31 | Erigeron annuus | Мног. | Не сильная | 79 | 54 | 94 | 15 | 1 |
| 32 | Erigeron canadensis | Мал. | Сильная | 23 | 50 | 62 | 59 | 0 |
| 33 | Galinsoga parviflora | » | » | 26 | 10 | 55 | 44 | 0 |
| 34 | Heracleum sosnowskyi | Мног. | Не сильная | 7 | 8 | 58 | 1 | 0 |
| 35 | Hippophae rhamnoides | » | Сильная | 5 | 11 | 16 | 78 | 0 |
| 36 | Impatiens glandulifera | Мал. | Не сильная | 12 | 3 | 20 | 37 | 0 |
| 37 | Impatiens parviflora | » | » | 1 | 1 | 3 | 97 | 4 |
| 38 | Lactuca serriola | Мног. | Сильная | 76 | 44 | 92 | 59 | 28 |
| 39 | Matricaria discoidea | Мал. | » | 42 | 31 | 71 | 10 | 0 |
| 40 | Oenothera biennis | » | Не сильная | 4 | 20 | 43 | 15 | 9 |
| 41 | Oxalis stricta | Мног. | » | 0.4 | 1 | 10 | 75 | 0 |
| 42 | Setaria pumila | Мал. | Сильная | 60 | 67 | 92 | 15 | 0 |
| 43 | Solidago canadensis | Мног. | Не сильная | 34 | 29 | 77 | 43 | 0 |
| 44 | Xanthium orientale | Мал. | Сильная | 73 | 92 | 96 | 49 | 0 |
Факторы, влияющие на интенсивность формирования корневых структур. На основе анализа значений AICc-весов можно заключить, что для оптимального по соотношению качество/сложность объяснения изменчивости развития АМ достаточно учитывать два фактора: степень натурализации вида растения и степень нарушенности местообитаний (табл. 2). Другие факторы – таксономический статус и длительность онтогенеза – не внесли заметного вклада в объяснение изменчивости характеристик развития микоризы. Интенсивность формирования корневых волосков лучше всего объяснялась степенью натурализации вида растения. Самая большая сумма AICc-весов в отношении мицелия темных септированных эндофитов была у предиктора “длительность онтогенеза вида”. Для всех изученных признаков таксономический статус не был оптимальным предиктором.
Таблица 2.
Качество объяснения (ΣW – суммы AICc-весов) изменчивости интенсивности формирования корневых структур разными предикторами
| Признак | Предикторы | |||
|---|---|---|---|---|
| степень натурализации | семейство | длительность онтогенеза | степень нарушен-ности местообитаний | |
| Ar% | 0.911 | 0.004 | 0.122 | 0.502 |
| Ve% | 0.428 | 0.007 | 0.102 | 0.875 |
| AM% | 0.761 | 0.017 | 0.196 | 0.452 |
| RH% | 0.647 | 0.001 | 0.226 | 0.222 |
| DSE% | 0.341 | 0.000 | 0.572 | 0.180 |
Таким образом, для анализа характеристик развития AM и корневых волосков следует использовать сочетание двух легко экологически интерпретируемых предикторов – степень натурализации вида растения и степень нарушенности местообитаний.
Арбускулярная микориза. Двухфакторный дисперсионный анализ с факторами “степень натурализации” и “степень нарушенности местообитаний” (табл. 3) показал, что в зависимости от степени натурализации вида значимо изменяются значения Ar% и AM%. Характеристики развития микоризы значимо изменяются в зависимости от степени нарушенности местообитаний. Отсутствие значимого взаимодействия факторов указывает на аддитивность, т.е. на простое суммирование, но не взаимную модификацию эффектов, связанных со степенью натурализации видов и степенью нарушенности местообитаний.
Таблица 3.
Результаты двух ANOVA интенсивности формирования корневых структур (F – критерий Фишера, P – уровень значимости)
| Признак | Факторы и число степеней свободы | |||||
|---|---|---|---|---|---|---|
| степень натурализации [1] (dF = 1) | степень нарушенности местообитаний [2] (dF = 1) | [1] × [2] (dF = 1) | ||||
| F | P | F | P | F | P | |
| Двухфакторный ANOVA с факторами “степень натурализации” и “степень нарушенности местообитаний” | ||||||
| Ar% | 9.28 | 0.0041 | 5.34 | 0.0261 | 0.10 | 0.7501 |
| Ve% | 2.88 | 0.0976 | 7.11 | 0.0110 | 0.17 | 0.6819 |
| AM% | 5.81 | 0.0206 | 4.43 | 0.0417 | 0.01 | 0.9965 |
| RH% | 5.23 | 0.0275 | 2.14 | 0.1513 | 1.89 | 0.1764 |
| DSE% | 0.57 | 0.4528 | 0.01 | 0.9912 | 0.20 | 0.6597 |
| Трехфакторный ANOVA с факторами “степень натурализации”, “степень нарушенности местообитаний” и “длительность онтогенеза” | ||||||
| Ar% | 4.60 | 0.0388 | 5.61 | 0.0234 | 0.81 | 0.3748 |
| Ve% | 1.42 | 0.2406 | 7.18 | 0.0111 | 0.26 | 0.6165 |
| AM% | 3.01 | 0.0912 | 4.91 | 0.0332 | 0.67 | 0.4168 |
| RH% | 3.97 | 0.0539 | 1.59 | 0.2151 | 3.09 | 0.0871 |
| DSE% | 0.01 | 0.9520 | 0.41 | 0.5266 | 0.03 | 0.8622 |
Интенсивность колонизации структурами арбускулярной микоризы была несколько выше у растений, собранных в сильно нарушенных местообитаниях с несомкнутыми сообществами, по сравнению с мало и средне нарушенными местообитаниями (рис. 1). Средняя интенсивность формирования арбускул в мало и средне нарушенных местообитаниях составляла 38 ± 5%, в сильно нарушенных – 44 ± 8%. Соответствующие средние значения для везикул – 26 ± 4 и 43 ± 7%; всех грибных структур – 59 ± 5 и 68 ± 8%. С использованием критерия Тьюки, рассчитанного в двухфакторном ANOVA, значимость различий в зависимости от нарушенности местообитаний подтвердилась только для везикул (P = 0.0282).
Рис. 1.
Интенсивность формирования арбускул (а), везикул (б) и всех микоризных структур (в) в корнях аборигенных и чужеродных растений в мало, средне (⚪) и сильно (⚫) нарушенных местообитаниях (размахи – SE).
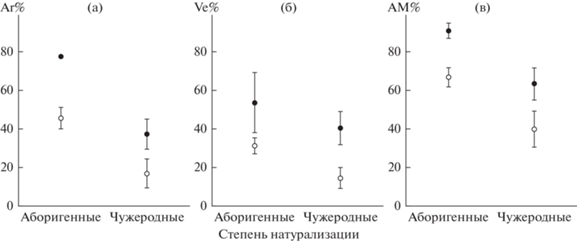
Интенсивность формирования всех структур арбускулярной микоризы у чужеродных видов была ниже, чем у аборигенных. Соответствующие средние значения для чужеродных и аборигенных видов составляли: интенсивности формирования арбускул – 28 ± 6 и 48 ± 5%; везикул – 28 ± 6 и 33 ± 4%; всех структур – 52 ± 7 и 68 ± 5%. Эти различия, хотя и небольшие, в отношении двух признаков подтверждены с помощью критерия Тьюки, рассчитанном в двухфакторном ANOVA: арбускулы – P = 0.0104; все структуры – P = 0.0423. Таким образом, интенсивность формирования арбускул сильнее, чем прочих микоризных структур, связана со степенью натурализации вида в местной флоре.
Чтобы оценить надежность заключения об особенностях формирования АМ у аборигенных и чужеродных растений, дополнительно к двухфакторному ANOVA провели анализ с учетом, во-первых, меньшего, во-вторых, большего числа факторов. В однофакторных ANOVA различия между аборигенными и чужеродными растениями проявились только по интенсивности формирования арбускул (F(1;42) = 6.64; P = 0.0135). В трехфакторном ANOVA с факторами “степень натурализации”, “степень нарушенности местообитаний” и “длительность онтогенеза” различия между аборигенными и чужеродными растениями проявились также только по интенсивности формирования арбускул (см. табл. 3). Пониженная колонизация всеми структурами АМ и повышенная интенсивность формирования корневых волосков у чужеродных растений в трехфакторных ANOVA статистически не подтвердились.
Корневые волоски. Двухфакторный дисперсионный анализ (см. табл. 3) показал, что значения RH% значимо изменяются в зависимости от натурализации вида, но не в зависимости от нарушенности местообитаний. Интенсивность формирования корневых волосков у чужеродных видов составляла 43 ± 6%, у аборигенных – 32 ± 4% (рис. 2); у малолетних трав – 38 ± 7% и многолетних растений – 36 ± 4% , и эти различия незначимы.
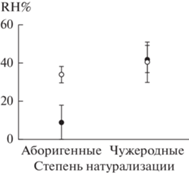
ОБСУЖДЕНИЕ
Результаты анализа представленного массива оценок позволяют наиболее надежно заключить, что у микоризных чужеродных растений Белорусского Полесья арбускулы формируются в корнях реже, чем у аборигенных растений. Мы тщательно отобрали для включения в анализ только такие виды, у которых непосредственно наблюдали арбускулы. Поэтому наши результаты характеризуют фактически обилие или количество арбускул в корнях только микоризных растений и не затрагивают такой аспект, как оценка соотношения микоризных и немикоризных видов во фракции чужеродных в регионе Белорусского Полесья. Заключение о пониженной интенсивности образования арбускул у чужеродных растений надежно в том смысле, что оно не является следствием погрешностей, связанных с различием между аборигенными и чужеродными растениями по таксономической принадлежности, типу онтогенеза или степени нарушенности местообитаний. Вывод о пониженном обилии арбускул воспроизвелся в статистических сравнениях разного дизайна.
Арбускулы – наиболее типичные или определяющие структуры арбускулярной микоризы. Их наличие свидетельствует о полноценном формировании симбиоза [29], поэтому пониженную интенсивность заложения арбускул у чужеродных растений можно обоснованно интерпретировать как свидетельство общей пониженной успешности образования и функционирования АМ, несмотря даже на то, что пониженная колонизация всеми структурами арбускулярных грибов у чужеродных растений статистически подтвердилась не очень надежно. Вывод о пониженной успешности образования AM у чужеродных растений по сравнению с аборигенными согласуется с одной частью опубликованных оценок [12–15], но не согласуется с другой [10, 11, 13, 14, 16, 17, 30, 31]. Наш результат поддерживает представление о существовании широкого диапазона способности чужеродных растений к микоризообразованию во вторичных ареалах. Наиболее вероятным механизмом установленных особенностей кажется приспособленность аборигенных и чужеродных растений к взаимодействию с разными видами грибов [11]. Это объяснение справедливо, в частности, для пары видов, включенных в массив, обсуждаемый в настоящей работе, – чужеродного Solidago canadensis и местного S. virgaurea [32].
В отличие от исследований, выполненных на единичных модельных видах [16, 30, 31], мы обнаружили большой разброс оценок общей колонизации АМ у разных видов чужеродных растений: например, диапазон значений общей колонизации – от 3 до 96%, интенсивности формирования арбускул – от 0.4 до 79%. При таком большом варьировании общие заключения могут сильно зависеть от критериев формирования списка исследуемых видов. Поэтому мы специально оценили факторы, которые могли сказаться на развитии микоризы и строении корней, помимо степени натурализации.
Интенсивность колонизации АМ в нашем массиве оценок не продемонстрировала какой-либо связи с таксономическим положением растений, хотя такая связь прослеживается на уровне крупных таксономических подразделений [7]. По-видимому, этот результат можно объяснить в том числе и тем, что в списке изученных видов только сем. Asteraceae представлено значительным числом, а представители остальных семейств фактически единичны.
В то же время интенсивность образования арбускул, везикул и всех структур АМ, вероятно, регулируется экологически, так как зависит от нарушенности местообитаний. По нашим данным, больше микоризы формируется у растений в сильно нарушенных местообитаниях по сравнению с мало нарушенными. Это, возможно, объясняется тем, что почвы нарушенных местообитаний могут быть бедны ресурсами, и инвестиции в АМ необходимы для выживания растений, как это показано для сообществ на песчаных почвах [15]. Кроме того, следует учитывать, что в нашем случае мало нарушенные местообитания – это преимущественно затененные участки с выраженным доминированием древесных растений, а сильно нарушенные – исключительно нелесные открытые участки.
Таким образом, возможно, что обилие микоризы в корнях связано с градиентом освещенности: много микоризы образуется при хорошем освещении и меньше – в условиях затенения. Важно, что микориза активнее формировалась в нарушенных местообитаниях и у аборигенных, и у чужеродных растений. Учет фактора “местообитание” позволил разделить общую дисперсию характеристик микоризообразования на компоненты. Хотя различия, сопряженные со степенью нарушенности местообитаний, были небольшие, их учет оказался необходимым для выявления эффектов, связанных со степенью натурализации видов.
Вывод о повышенной интенсивности развития корневых волосков у чужеродных растений статистически не столь надежен, как вывод о пониженной успешности микоризообразования. Однако можно предположить, что обе реакции – это части общего тренда, который феноменологически описывается так, что чужеродные растения более приспособлены к автономному осуществлению почвенного питания по сравнению с аборигенными. Это подтверждает отрицательная корреляция между интенсивностью колонизации AM и интенсивностью развития корневых волосков (рис. 3). Такая корреляция статистически значима у чужеродных растений (r = –0.51; n = 19; P = 0.0249), но не у аборигенных (r = –0.03; n = 25; P = 0.8951).
Рис. 3.
Связь интенсивностей колонизации AM (AM%) и формирования корневых волосков (RH%) у аборигенных (⚪; штриховая линия) и чужеродных (⚫; сплошная линия) растений.

Следовательно, наши данные можно интерпретировать как подтверждение альтернативного проявления двух приспособлений для осуществления почвенного питания – автономного (корневые волоски) и симбиотического (микориза). У части чужеродных растений поглощение ресурсов возможно с помощью активного образования корневых волосков, и необходимость в задействовании симбиотического приспособления минимальна. Похожий альтернативный характер формирования микоризы и корневых волосков ранее наблюдался у растений литорали Белого моря [33] и травянистых растений, росших на почве из зарослей инвазивного Acer negundo [34]. Это подтверждает возможный неслучайный характер сопряженности между обилием у чужеродных растений АМ и корневых волосков. Вместе с тем очевидно, что низкая статистическая поддержка связи между переменными и отсутствие предположений о механизмах, обеспечивающих эту связь, придает приведенным рассуждениям гипотетический характер. Для строгого обоснования сопряженности между интенсивностью образования корневых волосков и микоризы необходимы специальные исследования.
По нашим данным, у чужеродных растений не обнаружены особенности колонизации темными септированными эндофитами, хотя известно, что их развитие зависит от вида растения и содержания органического вещества в почве [35].
ЗАКЛЮЧЕНИЕ
У арбускулярно микоризных чужеродных растений Белорусского Полесья арбускулы в корнях развиты менее интенсивно, чем у аборигенных. Этот вывод не является следствием погрешностей, связанных с различием между аборигенными и чужеродными растениями по таксономической принадлежности, типу онтогенеза или степени нарушенности местообитаний. Различия между аборигенными и чужеродными растениями наиболее заметно проявились по формированию арбускул, а не везикул или гиф арбускулярных грибов в корнях. Это позволяет считать, что у чужеродных растений арбускулярная микориза функционирует менее эффективно, чем у аборигенных. Таким образом, наши результаты свидетельствуют о потенциальной меньшей зависимости чужеродных растений от взаимодействия с сообществами местных микоризных грибов.
Работа выполнена при поддержке РФФИ (проект № 14-04-90019_Бел-а – полевые исследования; проект № 20-54-00024_Бел-а – анализ данных) и в рамках темы госзадания Института экологии растений и животных УрО РАН.
Авторы выражают благодарность М.А. Конопленко, участвовавшему в сборе полевых материалов.
Авторы декларируют отсутствие конфликта интересов.
Список литературы
Grime J.P., Hodson J.G., Hunt R. Comparative plant ecology: a functional approach to common British flora. London: Unwin Hyman, 1988. 742 p.
Cornelissen J.H.C., Aerts R., Cerabolini B. et al. Carbon cycling traits of plant species are linked with mycorrhizal strategy // Oecologia. 2001. V. 129. № 4. P. 611–619.
van der Heijden M., Klironomos J., Ursic M. et al. Mycorrhizal fungal diversity determines plant biodiversity, ecosystem variability and productivity // Nature. 1998. V. 396. P. 69–72. https://doi.org/10.1038/23932
Betekhtina A.A., Veselkin D.V. Prevalence and intensity of mycorrhiza formation in herbaceous plants with different types of ecological strategies in the Middle Urals // Rus. J. Ecol. 2011. V. 42. № 3. P. 192–198.
Hempel S., Götzenberger L., Kühn I. et al. Mycorrhizas in the Central European flora: relationships with plant life history traits and ecology // Ecology. 2013. V. 94. № 6. P. 1389–1399. https://doi.org/10.1890/12-1700.1
Brundrett M.C. Mycorrhizal associations and other means of nutrition of vascular plants: understanding the global diversity of host plants by resolving conflicting information and developing reliablemeans of diagnosis // Plant Soil. 2009. V. 320. P. 37–77.
Wang B., Qiu Y.-L. Phylogenetic distribution and evolution of mycorrhizas in land plants // Mycorrhiza. 2006. V. 16. № 5. P. 299–363. https://doi.org/10.1007/s00572-005-0033-6
Pringle A., Bever J.D., Gardes M. et al. Mycorrhizal symbioses and plant invasions // Annu. Rev. Ecol. Evol. 2009. V. 40. № 1. P. 699–715.
Pyšek P., Guo W.-Y., Štajerová K. et al. Facultative mycorrhizal associations promote plant naturalization worldwide // Ecosphere. 2019. V. 10. № 11. e02937. https://doi.org/10.1002/ecs2.2937
Menzel A., Hempel S., Klotz S. et al. Mycorrhizal status helps explain invasion success of alien plant species // Ecology. 2017. V. 98. № 1. P. 92–102.
Bunn R.A., Ramsey P.W., Lekberg Y. Do native and invasive plants differ in their interactions with arbuscular mycorrhizal fungi? A meta-analysis // J. Ecol. 2015. V. 103. № 6. P. 1547–1556.
Marler M.J., Zabinski C.A., Callaway R.M. Mycorrhizae indirectly enhance competitive effects of an invasive forb on a native bunchgrass // Ecology. 1999. V. 80. № 4. P. 1180–1186.
Fitter A.H. Darkness visible. Reflections on underground ecology // J. Ecol. 2005. V. 93. P. 231–243.
Shah M.A., Reshi Z.A., Khasa D.P. Arbuscular mycorrhizas: drivers or passengers of alien plant invasion // Bot. Rev. 2009. V. 75. № 4. P. 397–417.
Endresz G., Somodi I., Kalapos T. Arbuscular mycorrhizal colonisation of roots of grass species differing in invasiveness // Community Ecol. 2013. V. 14. № 1. P. 67–76.
DeBellis T., Kembel S.W., Lessard J.P. Shared mycorrhizae but distinct communities of other root-associated microbes on co-occurring native and invasive maples // PeerJ. 2019. V. 7. e7295. https://doi.org/10.7717/peerj.7295
Moyano J., Rodriguez Cabal M.A., Nuñez M.A. Invasive trees rely more on mycorrhizas, countering the ideal weed hypothesis // Ecology. 2021. V. 102. e03330. https://doi.org/10.1002/ecy.3330
Reinhart K.O., Callaway R.M. Soil biota and invasive plants //New phytol. 2006. V. 170. № 3. P. 445–457.
Meinhardt K.A., Gehring C.A. Disrupting mycorrhizal mutualisms: a potential mechanism by which exotic tamarisk outcompetes native cottonwoods // Ecol. Appl. 2012. V. 22. P. 532–549. https://doi.org/10.1890/11-1247.1
Grove S., Haubensak K.A., Gehring C., Parker I.M. Mycorrhizae, invasions, and the temporal dynamics of mutualism disruption // J. Ecol. 2017. V. 105. P. 1496–1508. https://doi.org/10.1111/1365-2745.12853
Gusev A.P. Features of plant succession in landscapes disturbed by anthropogenic activity (by example of Southeastern Belarus) // Contemp. Probl. Ecol. 2012. V. 5. № 2. P. 174–178.
Gusev A.P. Plant invasions and landscape ecological state indication // Tyumen State University Herald. 2012. № 12. P. 168–174.
Panasenko N.N. Blacklist of flora of Bryansk oblast // Rus. J. Biol. Invasions. 2014. V. 5. № 3. P. 203–205.
The World Checklist of Vascular Plants (WCVP) [Электронный ресурс]. URL: https://wcvp.science. kew.org/ (дата обращения: 26.01.2022).
Селиванов И.А. Микосимбиотрофизм как форма консортивных связей в растительном покрове Советского Союза. М.: Наука, 1981. 232 с.
McGonigle T.P., Miller M.H., Evans D.G. et al. A new method which gives an objective-measure of colonization of roots by vesicular arbuscular mycorrhizal fungi // New Phytol. 1990. V. 115. P. 495–501.
Akhmetzhanova A.A., Soudzilovskaia N.A., Onipchenko V.G. et al. A rediscovered treasure: mycorrhizal intensity database for 3000 vascular plants species across the former Soviet Union // Ecology. 2012. V. 93. № 3. P. 689–690.
Burnham K.P., Anderson D.R. Model selection and multimodel inference: a practical information – theoretical approach. N.Y.: Springer, 2002. 488 p.
Smith S.E., Read D.J. Mycorrhizal symbiosis, London: Acad. Press, 2010. 803 p.
Kovacs G.M., Szigetvari C. Mycorrhizae and other root-associated fungal structures of the plants of a sandy grassland on the Great Hungarian Plain // Phyton. 2002. V. 42. № 2. P. 211–223.
Fumanal B., Plenchette C., Chauvel B., Bertagnolle F. Which role can arbuscular mycorrhizal fungi play in the facilitation of Ambrosia artemisiifolia L. invasion in France // Mycorrhiza. 2006. V. 17. № 1. P. 25–35.
Betekhtina A.A., Mukhacheva T.A., Kovalev S.Yu. et al. Abundance and diversity of arbuscular mycorrhizal fungi in invasive Solidago canadensis and indigenous S. virgaurea // Rus. J. Ecol. 2016. V. 47. № 6. P. 605–609. https://doi.org/10.1134/S1067413616060035
Веселкин Д.В., Марковская Е.Ф., Бетехтина А.А. и др. Микоризообразование у сосудистых растений береговой зоны западного побережья Белого моря // Учен. зап. Петрозаводского гос. ун-та. 2016. № 8 (161). С. 20–26.
Веселкин Д.В., Рафикова О.С., Екшибаров Е.Д. Почва из зарослей инвазивного Acer negundo неблагоприятна для образования микоризы у аборигенных трав // Журн. общ. биол. 2019. Т. 80. № 3. С. 214–225. https://doi.org/10.1134/S0044459619030084
Han L., Zuo Y., He X. et al. Plant identity and soil variables shift the colonisation and species composition of dark septate endophytes associated with medicinal plants in a northern farmland in China // Appl. Soil Ecol. 2021. V. 167. 104042. https://doi.org/10.1016/j.apsoil.2021.104042
Дополнительные материалы отсутствуют.