

Экология, 2022, № 4, стр. 289-298
Динамика проективного покрытия и видовой насыщенности травяных сообществ в ходе восстановительной сукцессии на залежах и отвалах
Т. В. Жуйкова a, *, Э. В. Мелинг a, В. С. Безель b
a Нижнетагильский государственный социально-педагогический институт, филиал ФГАОУ ВО
“Российский государственный профессионально-педагогический университет”
622031 Нижний Тагил, ул. Красногвардейская, 57, Россия
b Институт экологии растений и животных УрО РАН
620144 Екатеринбург, ул. 8 Марта, 202, Россия
* E-mail: hbfnt@rambler.ru
Поступила в редакцию 23.01.2022
После доработки 15.02.2022
Принята к публикации 16.02.2022
- EDN: TSIWJG
- DOI: 10.31857/S0367059722040084
Аннотация
Изучена временна́я динамика видовой насыщенности и проективного покрытия видов травяных сообществ, формирующихся на залежах и отвалах. В период 2006–2019 гг. на территории Притагильской зоны Среднего Урала проведены ежегодные исследования данных показателей фитоценозов четырех групп, отражающих последовательные сукцессионные этапы: злаковые с сохранением статуса в период исследования, злаковые с последующим переходом на луговую стадию, ранние луговые и сформированные луговые. Направление сукцессионного развития сообществ зависит от эдафических факторов. В условиях низкого загрязнения почв тяжелыми металлами на агроземах и техноземах развиваются фитоценозы с проективным покрытием и видовой насыщенностью, характерными для луговых сообществ фоновых территорий. В условиях высокого загрязнения луговая стадия непродолжительна и характеризуется низкими значениями исследуемых показателей. Возможны и другие траектории сукцессии: длительная задержка на злаковой стадии или пропуск луговой стадии и переход от злаковой к древесно-кустарниковой. Из двух анализируемых признаков более консервативна видовая насыщенность сообществ по сравнению с суммарным проективным покрытием видов, скорость развития которого в большей степени определяется сукцессионным возрастом сообщества, чем типом субстрата.
В последнее время интерес геоботаников связан с изучением частных закономерностей разных типов сукцессий [1]. Восстановительные сукцессии на залежных землях в различных регионах могут идти разными путями, но приводят к формированию терминальных зональных сообществ [2, 3]. Особенности сукцессионных траекторий обусловлены экологическим контекстом, региональным пулом видов, а также наследием прежнего землепользования, которое зависит от длительности использования пашни, площади и способа обработки залежных участков [4–7].
Обширная литература посвящена исследованиям сукцессий на разных типах отвалов в различных регионах [8–10 и др.], в том числе на Среднем Урале [11–14 и др.]. Показано [14–16 и др.], что при общей зональной направленности формирования растительности на отвалах наблюдается своеобразие растительных сообществ, определяемое конкретными экологическими условиями. Восстановление растительного покрова на отвалах Нижнего Тагила – города с трехсотлетней деятельностью горно-добывающей и металлургической промышленности, изучено недостаточно [17–19]. Сукцессионные процессы на залежах этой территории находятся на начальной стадии исследования [20–23].
В настоящей работе рассмотрена временна́я динамика видовой насыщенности (ВН – число видов на фиксированных площадках размером 1 м2), суммарного проективного покрытия видов в сообществах (СПП), формирующихся на залежах и отвалах Притагильской зоны Среднего Урала. Обсуждается следующая гипотеза: на динамику проективного покрытия и видовой насыщенности фитоценозов в ходе восстановительной сукцессии влияют сукцессионный возраст сообщества и тип субстрата – агрозем или технозем и степень их загрязнения тяжелыми металлами. На техноземах возможны различные траектории сукцессионного развития, в основе которых лежит разнообразие экологических условий.
МАТЕРИАЛ И МЕТОДЫ
Район исследования. Работы выполнены на территории Притагильской зоны Среднего Урала (г. Нижний Тагил Свердловской области, 58° с.ш., 60° в.д.). Изучены травяные сообщества разных сукцессионных стадий, формирующиеся на двух различных почвенных субстратах: на агроземах (А) и техноземах (Т) с разным уровнем почвенного загрязнения тяжелыми металлами (ТМ). Суммарный показатель токсической нагрузки (Z, отн. ед.) вычисляли на основании содержания в почве (мкг/г) Cd2+, Pb2+, Cu2+, Zn2+, Fe3+. Учитывали элементы, превышающие региональный фоновый уровень (табл. 1).
Таблица 1.
Характеристика видовой насыщенности и суммарного проективного покрытия фитоценозов (М ± m)
| Группа сообществ | Участки/период наблюдений | Количество лет наблюдений* | Z, отн. ед. | Видовая насыщенность, видов/м2 | Суммарное проективное покрытие, % |
|---|---|---|---|---|---|
| Злаковые | Т1 2010–2019 |
10 | 16.23 | 10.28 ± 1.79 | 80.46 ± 20.46 |
| Т2 2007–2019 |
10 | 5.17 | 9.40 ± 1.48 | 80.22 ± 6.69 | |
| Переходные злаковые | А1 2010–2015 |
6 | 1.53 | 13.43 ± 3.54 | 88.40 ± 34.12 |
| Т3 2006–2013 |
7 | 26.69 | 14.34 ± 2.79 | 86.97 ± 44.02 | |
| Ранние луговые | А1 2016–2019 |
4 | 1.53 | 14.75 ± 2.22 | 123.40 ± 30.71 |
| Т3 2014–2019 |
6 | 26.69 | 11.48 ± 4.24 | 75.01 ± 17.08 | |
| Луговые | А2 2011–2019 |
9 | 1.44 | 21.04 ± 1.73 | 145.50 ± 21.45 |
| А3 2009–2019 |
11 | 3.22 | 17.49 ± 1.69 | 134.15 ± 12.98 | |
| А4 2009–2019 |
11 | 3.33 | 19.64 ± 3.19 | 166.48 ± 15.05 | |
| А5 2006–2019 |
13 | 1.88 | 15.84 ± 1.44 | 134.19 ± 19.61 | |
| Т4 2007–2019 |
12 | 6.14 | 18.26 ± 1.35 | 145.29 ± 20.78 |
Участки исследования. Участки агроземов представлены залежными землями, выведенными из-под пашни в разное время (10–30 лет назад). Порядковый номер участка отражает увеличение возраста залежи и соответствующих фитоценозов – А1 → A2 → А3 → А4 → А5.
Участки техноземов расположены на промышленных отвалах, возраст которых более 50 лет. В этих условиях формируются молодые почвы по буроземному и литоземному типам на техногенной почвообразующей породе, богатой обменными основаниями и элементами питания растений [20]. Порядковый номер участков присвоен в соответствии с увеличением времени с момента отсыпки террас отвалов: Т1 → Т2 → Т3 → Т4. Подробная характеристика участков и периоды наблюдения за фитоценозами этих территорий представлены ранее [23].
Описание фитоценозов. Видовой состав сообществ и проективное покрытие (ПП) видов изучали ежегодно в период максимального развития травостоя (июль) на типичных по составу и структуре участках. Оценка видовой насыщенности (ВН) проведена на 5 учетных площадках (1.0 м2), расположенных методом конверта в пределах исследуемых сообществ на постоянной площади 100 м2, ПП – на 20 (по 0.25 м2). На основе ПП каждого вида рассчитано суммарное проективное покрытие (СПП). Исследования выполнены стандартными методами [24]. В работе представлены обобщенные данные за 14 лет (2006–2019 гг.).
Статистический анализ результатов выполнен с учетом среднего арифметического (М) и его ошибки (m). Сравнение СПП и ВН в пределах сукцессионных групп проведено с помощью критерия Крускаля-Уоллиса (H-критерий). О направлении и скорости развития анализируемых показателей судили по наклону линии регрессии, коэффициенту детерминации (R2). Изменение ВН и СПП (у) в исследуемый период (t) аппроксимируется уравнением линейной регрессии: у = = а + bt, где коэффициент а отвечает за сдвиг линии по вертикали, b – за наклон линии регрессии. Различия динамики анализируемых показателей в разных вариантах за время наблюдения характеризовали также с помощью средней геометрической (${{{{\overline х }}_{g}}}$), вычисленной на основании абсолютных значений изменений по годам СПП и ВН [25].
Групповые сравнения углов наклона линий регрессии проведены методом линейных контрастов (S-методом Шеффе) [26]. Сравниваются любые комбинации углов наклона (b ± SE). Например, для сравнения скорости развития СПП злаковых (Т1 и Т2) и луговых (А3–А5, Т4) сообществ используется контраст ψ = [(Т1 + Т2)/2] – [(А3 + + А4 + А5 + Т4)/4], в котором сравниваются углы наклона линий регрессии СПП групп злаковых и луговых фитоценозов. До обсуждения значимости любого оцениваемого контраста необходимо оценить дисперсию ψ(σψ). Оценка дисперсии задается выражением $\sigma _{\psi }^{2} = M{{S}_{\varpi }}\left( {\frac{{c_{1}^{2}}}{{{{n}_{1}}}} + \frac{{c_{2}^{2}}}{{{{n}_{2}}}} + ... + \frac{{c_{j}^{2}}}{{{{n}_{j}}}}} \right)$, где MSω – “средний квадрат внутри”; сj – константа, на которую умножается j-е среднее, nj – число наблюдений в j-й группе (среднее количество лет наблюдений в сообществах). Далее составляется отношение ψ к σψ. Число степеней свободы df, связанных с оценкой дисперсии, определяется величиной df для MSω, N – J, где N – общее количество лет наблюдений во всех сообществах (N = = n1 + n2 + …+ nj – в нашем случае 57 лет), J – количество сообществ, участвующих в анализе (в нашем случае 6). Фактическое значение F = $\Psi {\text{/}}{{\sigma }_{\psi }}$ сравнивали со стандартным значением F-распределения со степенями свободы J – 1 и N – J.
Статистическую обработку и построение графических иллюстраций выполняли с помощью стандартных пакетов программ Microsoft Excel 2007 и Statistica v. 10.0 (StatSoft, Inc., 2012).
РЕЗУЛЬТАТЫ
В данной работе рассмотрены описанные ранее четыре группы фитоценозов, отражающие последовательные сукцессионные этапы [23]:
1) злаковые с сохранением сукцессионного статуса в течение наблюдаемого периода (далее злаковые);
2) переходные злаковые – стадия, предшествующая непосредственному переходу на луговую;
3) ранние луговые (рудерализированные луга), формирующиеся на основе злаковой стадии, включают виды предыдущей стадии с высоким ПП;
4) стабильно луговые – характеризуются большим видовым разнообразием злаков, являющихся эдификаторами луговых сообществ (далее луговые). В пределах этой группы различали молодое луговое сообщество (участок А2) и зрелые (участки А3–А5, Т4) (см. табл. 1).
При определении стадий сукцессии опирались на представление о рудеральных сообществах с преобладанием многолетних злаков Bromopsis inermis, Elytrigia repens, Calamagrostis epigeios как предшествующей лугам стадии восстановительной сукцессии [27] и оценивали эту стадию как злаковую. Сообщество с доминированием рыхлокустовых злаков рассматривали как луговую стадию в соответствии с представлениями В. Р. Вильямса о зрелой стадии развития луга [28].
Характеристика ВН и СПП. Исследуемые территории отличаются по характеру протекающих сукцессионных процессов в травянистых сообществах. В большинстве случаев встречаются разновозрастные сообщества, находящиеся на одной сукцессионной стадии: злаковой (Т1, Т2) или луговой (А2–А5 и Т4). Только на двух участках (А1 и Т3) представлена серия, включающая две стадии: злаковую и луговую. Это позволяет сопоставить динамические процессы, протекающие в относительно стабильный и переходный периоды. Показатели видовой насыщенности и суммарного проективного покрытия исследуемых фитоценозов приведены в табл. 1, а значимость различий СПП и ВН фитоценозов в пределах сукцессионных групп в табл. 2.
Таблица 2.
Различия видовой насыщенности и суммарного проективного покрытия между разными сообществами в пределах сукцессионных групп (H-критерий Крускаля-Уоллиса)
| Группа сообществ/участок | df | Видовая насыщенность, видов/м2 | Суммарное проективное покрытие, % | ||
|---|---|---|---|---|---|
| Н | p | Н | p | ||
| Злаковые/Т1, Т2 | 1, N = 100 | 2.65 | 0.113 | 0.59 | 0.441 |
| Переходные злаковые/А1, Т3 | 1, N = 65 | 1.01 | 0.314 | 0.10 | 0.764 |
| Ранние луговые/А1, Т3 | 1, N = 40 | 0.20 | 0.652 | 17.59 | 0.001 |
| Луговые/А2, А3, А4, А5, Т4 | 4, N = 280 | 58.45 | 0.001 | 55.58 | ⪡0.001 |
Злаковые фитоценозы, будучи хронически сериальной стадией сукцессии на техногенных отвалах, при отсутствии оптимальных эдафических условий, вероятно, связанных с несформированностью почв, имеют низкий уровень ВН и СПП по сравнению с остальными фитоценозами. Различия между сообществами этой группы статистически незначимы по исследуемым показателям (см. табл. 2).
Злаковые с последующим переходом на луговую стадию фитоценозы характеризуются более высокими значениями ВН и СПП по сравнению со стабильно злаковыми, но меньшими, чем у луговых. При этом обнаружено сходство ВН и СПП независимо от типа субстрата, на котором они развиваются (см. табл. 2). В отличие от них ранние луговые сообщества агроземов и техноземов статистически значимо различаются между собой по показателю СПП (см. табл. 2). При этом в сообществе участка А1 показатели СПП и ВН выше, а в Т3 ниже предшествующей переходной злаковой сукцессионной стадии (см. табл. 1).
Таким образом, влияние типа субстрата на развитие травяного покрова проявляется на более поздних сукцессионных стадиях. Если на переходной злаковой стадии зависимости СПП от типа субстрата (фоновые условия агроземов/высокий уровень почвенного загрязнения техноземов) не обнаружено, то на ранней луговой развитие проективного покрытия сообщества зависит от типа субстрата. Это свидетельствует об устойчивости переходной злаковой стадии к техногенной трансформации почв.
Отметим, что токсическая нагрузка на участках А1 и Т3 различалась в 17 раз и не снижалась за время наблюдений. Меньшая нагрузка на почвы А1 более благоприятна для реабилитации эдафических условий этого участка, чем можно объяснить более быстрый переход к следующему этапу сукцессии. Развитие травяного покрова на стадии раннего лугового сообщества участка Т3, видимо, лимитирует высокая техногенная нагрузка (см. табл. 1, значение Z). Это ставит вопрос о возможности его дальнейшего развития как лугового сообщества.
Луговые фитоценозы характеризуются максимально высоким уровнем ВН и СПП (см. табл. 1). Они формируются как на агроземах при региональном фоновом уровне загрязнения почв, так и на техноземах при слабом уровне почвенного загрязнения. Луговые сообщества участков А2–А5 и Т4 статистически значимо различаются между собой по рассматриваемым показателям (см. табл. 2), что связано с их различным сукцессионным возрастом и видовым составом, определяемым экологическим контекстом.
Сравнение выделенных групп фитоценозов (переходные злаковые, ранние луговые и луговые) на агроземах показало статистически значимые различия между ними по уровню ВН (H (2, N = 270) = 43.73; p ⪡ 0.001) и СПП (H (2, N = 270) = = 62.83; p ⪡ 0.001). Аналогичные различия между четырьмя группами фитоценозов, отражающими последовательные сукцессионные этапы, показаны для техноземов – ВН: H (3, N = 215) = 131.16; p ⪡ 0.001; СПП: H (3, N = 215) = 103.24; p ⪡ 0.001.
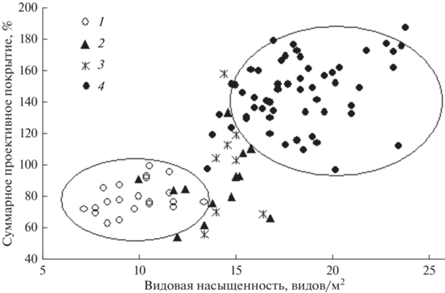
Полученные результаты являются доказательством правомерности выделения рассматриваемых групп серийных сообществ [23]. Увеличение ВН и СПП в сообществах агроземов в ходе сукцессии отражает общую закономерность сукцессионных процессов на залежах. На техноземах в условиях низкого загрязнения выдерживается общая закономерность восстановительной сукцессии – повышение ВН и СПП травяного покрова. При высокой токсической нагрузке рост ВН не сопровождается увеличением СПП. Выявленные закономерности демонстрирует рис. 1, отражающий связь СПП и ВН в ходе сукцессии.
Рис. 1.
Взаимосвязь суммарного проективного покрытия и видовой насыщенности исследуемых травяных сообществ: 1 – злаковые, 2 – переходные злаковые, 3 – ранние луговые, 4 – луговые.
Показатели, отражающие направление и скорость изменения СПП и ВН отдельных серийных сообществ в пределах сукцессионных групп приведены в табл. 3. Анализ был проведен отдельно для каждого серийного сообщества.
Таблица 3.
Значения коэффициентов детерминации, уравнения линейной регрессии у = а + bt и средней геометрической (${{\bar {x}}_{g}}$) для исследуемых показателей серийных сообществ
| Участок/группа сообществ | R2 | df | p | a ± SE | b ± SE | ${{\bar {x}}_{g}}$ |
|---|---|---|---|---|---|---|
| Видовая насыщенность | ||||||
| Т1 (злаковые) | 0.20 | 1; 8 | 0.199 | 8.81 ± 1.16 | 0.27 ± 0.18 | 0.31 |
| Т2 (злаковые) | 0.10 | 1; 9 | 0.374 | 8.71 ± 0.90 | 0.13 ± 0.13 | 0.28 |
| А1 (переходные злаковые) | 0.01 | 1; 4 | 0.875 | 13.17 ± 1.57 | 0.07 ± 0.40 | 0.52 |
| Т3 (переходные злаковые) | 0.07 | 1; 5 | 0.593 | 13.35 ± 2.02 | 0.22 ± 0.45 | 0.33 |
| А1 (ранние луговые) | 0.13 | 1; 2 | 0.225 | 14.50 ± 0.28 | 0.10 ± 0.16 | 0.16 |
| Т3 (ранние луговые) | 0.84 | 1; 4 | 0.009 | 21.95 ± 0.68 | –0.71 ± 0.17 | –0.8 |
| А2 (луговые) | 0.02 | 1; 7 | 0.695 | 21.53 ± 1.33 | –0.10 ± 0.24 | 0.15 |
| А3 (луговые) | 0.003 | 1; 9 | 0.884 | 17.34 ± 0.33 | 0.03 ± 0.17 | 0.22 |
| А4 (луговые) | 0.23 | 1; 9 | 0.258 | 17.00 ± 1.97 | 0.47 ± 0.29 | 0.60 |
| А5 (луговые) | 0.001 | 1; 11 | 0.933 | 15.90 ± 0.88 | –0.01 ± 0.11 | 0.17 |
| Т4 (луговые) | 0.06 | 1; 10 | 0.441 | 18.82 ± 0.85 | –0.09 ± 0.12 | 0.02 |
| Суммарное проективное покрытие | ||||||
| Т1 (злаковые) | 0.04 | 1; 8 | 0.828 | 71.29 ± 6.96 | –0.58 ± 1.09 | 0.89 |
| Т2 (злаковые) | 0.07 | 1; 9 | 0.462 | 77.86 ± 5.45 | 0.67 ± 0.77 | 3.0 |
| А1 (переходные злаковые) | 0.69 | 1; 4 | 0.041 | 48.13 ± 15.04 | 11.51 ± 3.86 | 15.95 |
| Т3 (переходные злаковые) | 0.36 | 1; 5 | 0.153 | 64.66 ± 13.85 | 5.57 ± 3.09 | 5.73 |
| А1 (ранние луговые) | 0.13 | 1; 2 | 0.638 | 106.25 ± 33.89 | 6.86 ± 12.37 | 3.28 |
| Т3 (ранние луговые) | 0.002 | 1; 4 | 0.933 | 74.89 ± 17.45 | 0.43 ± 4.48 | 0.67 |
| А2 (луговые) | 0.58 | 1; 7 | 0.023 | 113.48 ± 12.11 | 6.65 ± 2.15 | 6.03 |
| А3 (луговые) | 0.20 | 1; 9 | 0.163 | 123.54 ± 0.29 | 1.77 ± 1.16 | 1.71 |
| А4 (луговые) | 0.22 | 1; 9 | 0.142 | 149.27 ± 7.68 | 3.22 ± 1.13 | 1.96 |
| А5 (луговые) | 0.22 | 1; 11 | 0.085 | 117.74 ± 10.66 | 2.35 ± 1.21 | 2.71 |
| Т4 (луговые) | 0.43 | 1; 10 | 0.021 | 122.53 ± 10.66 | 3.51 ± 1.29 | 5.73 |
По динамике СПП установлена близость между злаковыми сообществами участков Т1 и Т2 – углы наклона линий регрессии статистически значимо не различаются (F = 2.81; df = 1; 16; p > 0.05), а также между зрелыми луговыми участков А3, А4, А5 и Т4 (F = 2.33; df = 3; 36; p > 0.05). Это позволяет при построении регрессионной зависимости СПП от временно́го ряда объединить Т1 и Т2, а также А3, А4, А5 и Т4 в пределах соответствующих сукцессионных стадий. Молодое луговое сообщество (участок А2) по динамике СПП, выраженной через угол наклона линии регрессии, значимо отличается от зрелых луговых фитоценозов участков А3, А4, А5, Т4 (F = 7.57; df = 4; 42; p < < 0.001) и при анализе динамических процессов рассматривается отдельно.
Переходные злаковые фитоценозы участков А1 и Т3 по динамике СПП не различаются (F = = 2.68; df = 1; 8; p > 0.05). Значительные различия (в 65 раз) в коэффициентах детерминации СПП установлены для ранних луговых сообществ участков А1 и Т3, однако эти различия статистически незначимы (табл. 4). Это обусловлено существенными межгодовыми колебаниями СПП, которые указывают на неустойчивость данной сукцессионной стадии. Принимая во внимание различия в коэффициентах детерминации, линии регрессии для каждого серийного сообщества построены отдельно.
Таблица 4.
Результаты сравнения углов наклона линии регрессии (b ± SE) видовой насыщенности и суммарного проективного покрытия групп сообществ (контрастов) методом линейных контрастов
| Контрасты | df | F | p |
|---|---|---|---|
| Видовая насыщенность | |||
| Т1 – Т2 | 1; 16 | 2.00 | >0.050 |
| [(А3 + А4 + А5)/3] – Т4 | 3; 36 | 5.00 | <0.010 |
| А2 – [(А3 + А4 + А5 + Т4)/4] | 4; 42 | 2.53 | >0.050 |
| [(Т1 + Т2)/2] – Т3 (переходные злаковые) | 2; 20 | 0.40 | >0.050 |
| Т3 (переходные злаковые) – Т3 (ранние луговые) | 1; 8 | 6.20 | <0.050 |
| Т3 (ранние луговые) – Т4 | 1; 14 | 12.4 | <0.010 |
| А1 (переходные злаковые) – А1 (ранние луговые) | 1; 5 | 0.18 | >0.050 |
| [(А2 + А3 + А4 + А5)/4] – А1 (ранние луговые) | 4; 36 | 0.03 | >0.050 |
| А1 (переходные злаковые) – Т3 (переходные злаковые) | 1; 8 | 0.26 | >0.050 |
| А1 (ранние луговые) – Т3 (ранние луговые) | 1; 8 | 1.48 | >0.050 |
| [(Т1 + Т2)/2] – [(А3 + А4 + А5 + Т4)/4] | 5; 51 | 2.00 | >0.050 |
| Суммарное проективное покрытие | |||
| Т1 – Т2 | 1; 16 | 2.81 | >0.050 |
| [(А3 + А4 + А5)/3] – Т4 | 3; 36 | 2.33 | >0.050 |
| А2 – [(А3 + А4 + А5 + Т4)/4] | 4; 42 | 7.57 | <0.001 |
| [(Т1 + Т2)/2] – Т3 (переходные злаковые) | 2; 20 | 6.16 | <0.010 |
| Т3 (переходные злаковые) – Т3 (ранние луговые) | 1; 8 | 2.96 | >0.050 |
| Т3 (ранние луговые) – Т4 | 1; 14 | 2.60 | >0.050 |
| А1 (переходные злаковые) – А1 (ранние луговые) | 1; 8 | 0.88 | >0.050 |
| [(А2 + А3 + А4 + А5)/4] – А1 (ранние луговые) | 4; 36 | 1.00 | >0.050 |
| А1 (переходные злаковые) – Т3 (переходные злаковые) | 1; 8 | 2.68 | >0.050 |
| А1 (ранние луговые) – Т3 (ранние луговые) | 1; 5 | 1.19 | >0.050 |
| [(Т1 + Т2)/2] – [(А3 + А4 + А5 + Т4))/4] | 5; 51 | 8.07 | <0.001 |
Для большинства рассматриваемых сообществ ВН в рассматриваемом временно́м периоде относительно стабильна независимо от сукцессионной стадии (рис. 2а, 3а). Статистически значимых различий по динамике ВН сообществ разных сукцессионных стадий не установлено (см. табл. 4). Исключением является раннее луговое сообщество технозема Т3 (2014–2019), поскольку характеризуется существенным снижением ВН (см. табл. 3) и значимо отличается от сообществ предшествующей переходной злаковой этого участка (2006–2013) и луговой стадии (участок Т4) при аналогичном субстрате, но низком загрязнении ТМ (см. рис. 3а, табл. 4). Полученные результаты свидетельствуют о регрессивном развитии травяного яруса раннего лугового сообщества участка Т3. Учитывая прогрессивное развитие древесно-кустарниковых растений на этом участке, описанное нами ранее [23], можно говорить о развитии со временем на данной территории лесного сообщества, минуя зрелую луговую стадию.
Рис. 2.
Динамика видовой насыщенности (а) и суммарного проективного покрытия (б) зрелых луговых (1) и злаковых (2) фитоценозов.
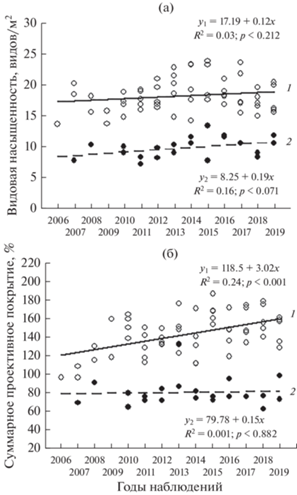
Рис. 3.
Динамика видовой насыщенности (а) и суммарного проективного покрытия (б) переходных злаковых и ранних луговых фитоценозов: 1 – переходное злаковое А1, 2 – переходное злаковое Т3, 3 – раннее луговое А1, 4 – раннее луговое Т3.
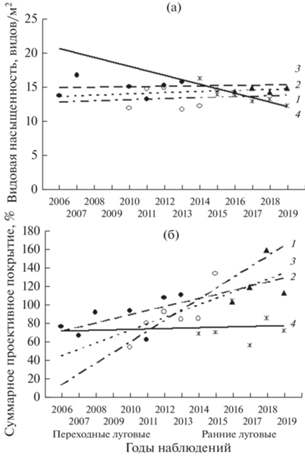
Сравнение сукцессионных групп между собой по скорости изменения СПП показало, что злаковые сообщества характеризуются слабовыраженной положительной динамикой в отличие от зрелых более динамичных луговых (F = 8.07; df = 5; 51; p < < 0.001; рис. 2б, табл. 4). Для раннего лугового сообщества участка Т3 (2014–2019) по показателю ВН характерны низкие значения R2 и ${{\bar {x}}_{g}}$, указывающие на слабо выраженные динамические процессы. Это сообщество по скорости развития СПП не отличается от злаковой стадии (F = 1.9; df = 2; 19; p > 0.05; табл. 3, рис. 3б). В зрелых луговых сообществах развитие СПП происходит более динамично (см. рис. 2б, 3, 4).
Рассматриваемые сообщества различаются и по скорости развития СПП, оцененной по значениям ${{\bar {x}}_{g}}$ (см. табл. 3). Наибольшая скорость наблюдается у сообществ агроземов – переходного злакового участка А1 (2010–2015) и молодого лугового (А2) (см. табл. 3, рис. 3б, 4). Средний годовой прирост СПП на них составляет 15.95 и 6.03% соответственно. На переходном злаковом Т3 (2006–2013) и луговом (Т4) СПП в среднем увеличивается на 5.73% в год. Меньше скорость развития СПП у раннего лугового участка А1 (2016–2019) и зрелых луговых (А3–А5) сообществ агроземов (см. табл. 3, рис. 4). Не наблюдается положительной динамики этого показателя и у раннего лугового фитоценоза техноземов Т3 (2014–2019).
Рис. 4.
Динамика суммарного проективного покрытия разновозрастных луговых сообществ: 1 – молодое (А2); 2 – зрелые (А3, А4, А5, Т4).
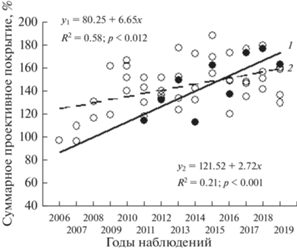
Таким образом, на более поздних стадиях развития луговых сообществ сохраняется характерный для ранних стадий рост СПП при неизменной ВН, при этом ежегодный прирост СПП замедляется. Это согласуется с общей закономерностью восстановительной сукцессии: увеличение обилия до оптимального уровня, соответствующего ограничивающей емкости окружающей среды. Далее наступает стабилизация этого показателя [29].
Влияние типа субстрата на скорость развития ВН и СПП проанализировано в серийных сообществах, находящихся на одной сукцессионной стадии, встречающейся на агроземах и техноземах. На переходной злаковой, ранней луговой и луговой стадиях динамика ВН, выраженная через ${{\bar {x}}_{g}}$, на агроземах выше, чем на техноземах. Однако статистически значимо различается ВН только на стадии луговых сообществ (участки А3–А5 и Т4: F = 5.0; df = 3; 36; p < 0.01). Влияние группы почв на скорость развития СПП проявляется на уровне тенденции на переходной злаковой и ранней луговой стадиях: показатель ${{\bar {x}}_{g}}$ выше на агроземе, чем на техноземе (см. табл. 3, 4). Зрелое луговое сообщество техноземов участка Т4 по скорости развития СПП близко к молодому луговому на агроземах (А2), что подтверждает высказанное ранее положение о задержке развития сообщества в условиях техноземов [20].
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Из двух анализируемых нами признаков более консервативна ВН сообществ, поскольку СПП развивается более динамично. При сохранении относительного постоянства ВН в луговых, переходных злаковых, а также раннем луговом сообществах на агроземах отмечено закономерное возрастание СПП. Это, вероятно, связано с увеличением количества особей и изменением их онтогенетического состояния, что ведет к формированию полночленной возрастной структуры составляющих сообщество ценопопуляций видов. Наибольший вклад в увеличение СПП в этих сообществах вносят виды семейства Poaceae. Так, в луговом фитоценозе А2 СПП пяти видов злаков (Festuca pratensis, F. rubra, Poa angustifolia, P. palustris, Deschampsia cespitosa) в 2011 г. составляло 39.0%, а в 2019 г. уже 50.5%. В фитоценозе на переходной злаковой стадии (участок А1) состав и количество видов злаков относительно стабильны, но наблюдается увеличение их проективного покрытия с 19.6 до 28.2% в период с 2010 г. по 2015 г. Все это отражает тип развивающегося лугового фитоценоза.
Показатели злаковых сообществ характеризуются отсутствием выраженной положительной динамики. Длительное (более 10 лет) сохранение сообществ на этой стадии характерно для техноземов с доминированием Calamagrostis epigeios. Доля его участия в СПП сообществ на участках Т1 составляет 26–44%, а Т2 – 35–41%. СПП дерновинных злаков меняется незначительно: на Т1 – от 1.0 до 2.55% в период с 2010 г. по 2019 г., на Т2 – от 0.4 до 8.7% с 2007 г. по 2019 г. Следствием является простая архитектоника травяного покрова сообществ, не усложненная участием ценопопуляций других видов. Внедрение луговых видов ограничивается неразвитостью почвенных горизонтов, загрязнением почв ТМ и доминированием C. epigeios. Почва этих участков молодая, представлена слабовыраженной подстилкой, состоящей в основном из ветоши трав, и слабозадернованным гумусовым горизонтом АY мощностью 4.5–5.0 см [20]. Известно, что C. epigeios имеет стратегию демутационного эксплерента [30], энергично захватывая свободные территории [31], и доминирует на ранних стадиях сукцессий в течение нескольких лет или даже десятилетий [32]. Установлена также устойчивость этого вида к ТМ в фазе прорастания семян [33], он сдерживает развитие древесных растений и переход сообщества на древесно-кустарниковую стадию [34]. Ослабление позиций вейника наблюдается лишь при разрастании лесных фитоценозов на прилегающей территории.
В связи с сохранением обсуждаемых неблагоприятных эдафических (слабовыраженный гумусовый горизонт, высокий уровень загрязнения почв) и биотических (устойчивость вейника на нарушенных территориях) условий сообщества Т1 и Т2 задерживаются на злаковой стадии. Аналогичная задержка сукцессии при сохранении неблагоприятных эдафических условий показана на восстановленных площадках нефтяных и газовых скважин [35].
Раннее луговое сообщество участка Т3 (2014–2019) отличается по динамике исследуемых показателей. Отмечено снижение ВН и отсутствие положительной динамики СПП, что сопровождается возобновлением древесных растений (СПП от 2.9 до 10.0%). Однако такое значение не является пороговым для перехода на кустарниковую или лесную стадии. Кустарниковая стадия сукцессии характеризуется преобладанием древесных фанерофитов высотой от 0.5 до 5 м и покрытием >30% [36]. Для лесного сообщества в качестве порогового указывают покрытие полога 40% [37].
В нашем случае отмеченные изменения фитоценоза участка Т3 (2014–2019) свидетельствуют о регрессивном развитии травяного яруса раннего лугового сообщества. При высокой техногенной трансформации почв возможность перехода на зрелую луговую стадию с типичными для нашей зоны показателями ВН и СПП отсутствует. При низких концентрациях ТМ (участок Т4) развивается стабильное луговое сообщество, сохраняющее сукцессионный статус в течение 12 лет при ВН 18 видов/м2 и СПП 145.3%, что характерно для типичных региональных луговых фитоценозов агроземов (см. табл. 1, А2–А5) и послелесных луговых сообществ нашей зоны (по исследованиям авторов данной статьи ВН – 17 видов/м2, СПП – 151.8%).
Луговые сообщества, не изменяя сукцессионного статуса, имеют положительную динамику СПП. Это, вероятно, связано, с одной стороны, с повторными нарушениями в форме нерегулярного кошения, с другой – с густым травяным покровом (СПП от 134.2 до 166.5%), препятствующим развитию проростков древесных растений.
Таким образом, траектория сукцессионного развития травяных сообществ на исследуемой территории соответствует восстановительной сукцессии, при которой происходят увеличение видового богатства, повышение продуктивности сообществ и биоразнообразия. В нашем исследовании продемонстрировано увеличение ВН и СПП при переходе от злаковой к луговой сукцессионной стадии. Выявлены различные траектории сукцессии в зависимости от эдафических условий: 1) формирование типичного лугового сообщества (агрозем и технозем с низким почвенным загрязнением ТМ); 2) развитие раннего лугового сообщества с низкими показателями ВН и СПП (технозем с высоким загрязнением); 3) задержка развития сообщества на злаковой стадии (технозем). Возможен пропуск луговой стадии и переход от злаковой к древесно-кустарниковой (технозем с высоким загрязнением).
В ходе проведенных исследований показано влияние на формирование величины и динамики ВН и СПП следующих факторов: эдафических условий (в том числе уровня почвенного загрязнения); затенения при разрастании лесных фитоценозов на прилегающих территориях. Роль экологических условий в траектории сукцессии показана во многих работах [4–7, 15]. Характер наблюдаемых нами изменений соответствует модели пороговой динамики [37].
ВЫВОДЫ
1. Исследованный сукцессионный ряд сообществ: переходных злаковые, переходные злаковые, ранние луговые и луговые, демонстрирует увеличение видовой насыщенности и проективного покрытия, что свидетельствует об активно идущей восстановительной сукцессии на рассматриваемой территории Притагильской зоны Среднего Урала.
2. При неизменной ВН в пределах луговых, переходных злаковых и ранних луговых фитоценозов на агроземах СПП возрастает, что обусловлено увеличением количества особей и формированием полночленной возрастной структуры ценопопуляций, в том числе дерновинных злаков. Злаковые сообщества характеризуются отсутствием выраженной положительной динамики всех исследуемых показателей, что говорит о задержке этой стадии при неблагоприятных эдафических условиях.
3. При низком загрязнении почв ТМ на агроземах и техноземах наблюдается развитие фитоценозов с типичными признаками лугового сообщества регионального фона, при высоком луговая стадия непродолжительна, характеризуется низкими значениями ВН и СПП. Возможны пропуск луговой стадии и переход от злаковой к древесно-кустарниковой, а также задержка сукцессии на злаковой стадии.
4. Ведущим фактором, определяющим скорость развития суммарного проективного покрытия, является сукцессионный возраст сообщества. В градиенте сукцессиионного возраста наблюдается повышение скорости развития суммарного проективного покрытия при переходе от злаковой к переходной злаковой стадии с последующим снижением этого показателя. Проявление данной закономерности модифицируют эдафические условия. На переходных стадиях ежегодный прирост СПП на агроземах выше, чем на техноземах, на сформированной луговой – выше на техноземах.
Работа выполнена в рамках госзадания Нижнетагильского государственного социально-педагогического института ФГАОУ ВО “Российский государственный профессионально-педагогический университет” и Института экологии растений и животных УрО РАН.
Авторы декларируют отсутствие конфликта интересов.
Список литературы
Миркин Б.М., Наумова Л.Г. Основные тенденции развития теории синтаксономии и динамики растительных сообществ // Журнал общ. биол. 2014. Т. 75. № 2. С. 83–94.
Дикарева Т.В., Опарин М.Л. Растительность северной части сухих степей Заволжья и ее антропогенные производные на залежах и пастбищах // Поволжский экологич. журн. 2002. № 3. С. 199–216.
Титлянова А.А., Самбуу А.Д. Детерминированность и синхронность залежной сукцессии в степях Тувы // Изв. РАН. Серия биологич. 2014. № 6. С. 621–630.
Бембеева О.Г., Джапова P.P. Восстановительная сукцессия залежных земель в пустынной зоне Калмыкии // Изв. Самарского научного центра РАН. 2012. Т. 14. № 1 (5). С. 1195–1997.
Battaglia L.L., Minchin P.R., Pritchett D.W. Sixteen years of old-field succession and reestablishment of A bottomland hardwood forest in the Lower Mississippi alluvial valley // Wetlands. 2002. V. 22. № 1. P. 1–17.https://doi.org/10.1672/0277-5212(2002)022[0001: SYOOFS]2.0.CO;2
Knapp S., Stadler J., Harpke A., Klotz S. Dispersal traits as indicators of vegetation dynamics in long-termold-field succession // Ecological Indicators. 2016. V. 65. P. 44–54. https://doi.org/10.1016/j.ecolind.2015.10.003
Stadler J., Klotz S., Brandl R., Knapp S. Species richness and phylogenetic structure in plant communities: 20 years of succession // Web Ecol. 2017. V. 17. P. 37–46.
Конысбаева Д.Т. Влияние субстрата промышленных отвалов на формирование растительности // Вестник науки КГУ им. Байтурсынова. Костанай, 2002. С. 152–158.
Сибирина Л.А., Полохин О.В., Жабыко Е.В. Начальные этапы формирования растительного покрова на техногенных экотопах Приморского края // Изв. Самарского научного центра РАН. 2012. Т. 14. № 1 (6). С. 1539–1542.
Лиханова И.А., Лаптева Е.М., Ковалева В.А. Продуктивность растительных сообществ и формирование органогенных горизонтов почв в ходе самовосстановительной сукцессии на техногенных субстратах // Изв. Самарского научного центра РАН. 2014. Т.16. № 5. С. 182–189.
Лукина Н.В. Формирование фитоценозов на золоотвалах Южноуральской ГРЭС // Аридные экосистемы. 2010. Т. 16. № 4 (44). С. 62–69.
Лукина Н.В., Глазырина М.А., Филимонова Е.И. и др. Формирование растительности на отвалах Баженовского месторождения хризотил-асбеста // Изв. Самарского научного центра РАН. 2017. Т. 19. № 2 (2). С. 294–299.
Веселкин Д.В., Лукина Н.В., Чибрик Т.С. Соотношение микоризных и немикоризных видов растений в первичных техногенных сукцессиях // Экология. 2015. № 5. С. 417–424. [Veselkin D.V., Lukina N.V., Chibrik T.S. The ratio of mycorrhizal and nonmycorrhizal plant species in primary technogenic successions // Russ. J. of Ecology. 2015. V. 46. № 5. P. 345–353. https://doi.org/10.1134/S1067413615050203]
Чибрик Т.С., Лукина Н.В., Филимонова Е.И. и др. Формирование фитоценозов на золоотвалах Нижнетуринской ГРЭС // Проблемы региональной экологии. 2018. № 6. С. 27–29.
Манаков Ю.А., Куприянов А.Н. Критерии для диагностики первичных стадий сукцессии на отвалах Кузбасса // Горный информационно-аналитический бюлл. 2009. Т. 7. № 12. С. 186–193.
Наумова Л.Г. Экологическая ботаника. Ч. II. Фитоценология: Учеб. пособие-экстерн для магистрантов биологич. и экологич. направлений. Уфа: Вагант, 2012. 42 с.
Лукина Н.В., Филимонова Е.И., Глазырина М.А., Чибрик Т.С. Структурная организация лесных фитоценозов на промышленных отвалах Урала / Изв. Самарского научного центра РАН. 2015. Т. 17. № 6. С. 220–224.
Безель В.С., Жуйкова Т.В., Гордеева В.А. и др. Надземная биомасса и скорость деструкции растительных остатков в травянистых сообществах при загрязнении почвы тяжелыми металлами // Экология. 2016. № 4. С. 264–269. [Bezel’ V.S., Zhuikova T.V., Gor-deeva V.A. et al. Aboveground phytomass and rate of plant debris decomposition in herbaceous communities exposed to soil pollution with heavy metals // Russ. J. of Ecology. 2016. V. 47. №. 4. P. 343–348. https://doi.org/10.1134/S1067413616040081]
Мелинг Э.В., Жуйкова Т.В., Голоушкина Е.В., Гордеева В.А. Динамика травяных сообществ на отвалах горно-металлургической промышленности Нижнего Тагила // Биологические системы: устойчивость, принципы и механизмы функционирования : Сб. статей V Всерос. научн.-практ. конф. (c междунар. участ.). Нижний Тагил: Нижнетагильский гос. соц.-пед. ин-т, 2017. С. 237–246.
Жуйкова Т.В., Мелинг Э.В., Кайгородова С.Ю. и др. Особенности почв и травянистых растительных сообществ в условиях техногенеза на Среднем Урале // Экология. 2015. № 3. С. 163–172. [Zhuikova T.V., Meling E.V., Kaigorodova S.Yu. et al. Specific features of soils and herbaceous plant communities in industrially polluted areas of the Middle Urals // Russ. J. of Ecology. 2015. V. 46. №. 3. P. 213–221. https://doi.org/10.1134/S1067413615030133]
Жуйкова Т.В., Безель В.С., Бергман И.Е. и др. Зависимость фитомассы травяных сообществ, произрастающих на антропогенно трансформированных территориях, от погодных факторов // Сибирский экологич. журн. 2018. № 4. С. 489–501. [Zhuikova T.V., Bezel’ V.S., Bergman I.E. et al. Dependence of phytomass of herbaceous cenoses on weather factors in anthropogenically impacted areas // Contemporary Problems of Ecology. 2018. V. 11. № 4. P. 428–437. https://doi.org/10.1134/S1995425518040108]
Безель В.С., Жуйкова Т.В., Гордеева В.А. и др. Надземная биомасса и скорость деструкции растительных остатков в травянистых сообществах при загрязнении почвы тяжелыми металлами // Экология. 2016. № 4. С. 264–269 [Bezel’ V.S., Zhuikova T.V., Gordeeva V.A. et al. Aboveground phytomass and rate of plant debris decomposition in herbaceous communities exposed to soil pollution with heavy metals // Russ. J. of Ecology. 2016. V. 47. № 4. P. 343–348. https://doi.org/10.1134/S1067413614020039]
Жуйкова Т.В., Мелинг Э.В., Безель В.С. Динамика альфа-разнообразия в ходе восстановительной сукцессии травяных сообществ залежей и отвалов // Экология. 2022. № 3. С. 178–188.
Миркин Б.М., Розенберг Г.С. Фитоценология: принципы и методы. М.: Наука, 1978. 211 с.
Лакин Г.Ф. Биометрия. М.: Высш. школа, 1990. 352 с.
Гласс Дж., Стенли Дж. Статистические методы в педагогике и психологии. М.: Прогресс, 1976. 495 с.
Миркин Б.М., Наумова Л.Г., Соломещ А.И. Современная наука о растительности: Учеб. для вузов. М.: Логос, 2000. 264 с.
Работнов Т.А. Луговедение: Учеб. для ун-тов. 2-е изд. М.: Изд-во МГУ, 1984. 320 с.
Бигон М., Харпер Дж., Таунсенд К. Экология. Особи, популяции и сообщества: в 2-х т. М.: Мир, 1989. Т. 2. 477 с.
Уланова Н.Г. Структура и динамика популяций вейника наземного в различных экологических условиях // Роль заповедников России в сохранении и изучении природы : Mат-лы юбилейной научно-практич. конф. Сер. “Труды Окского государственного природного биосферного заповедника”. Рязань: Изд-во: НП “Голос губернии”, 2015. С. 201–207.
Prach K., Pyšek P. How do species dominating in succession differ from others? // Journal of Vegetation Science. 1999. V. 10. P. 383–392.
Работнов Т.А. Фитоценология. 2-е изд. М.: Изд-во МГУ, 1983. 296 с.
Маджугина Ю.Г., Кузнецов Вл.В., Шевякова Н.И. Растения полигонов захоронения бытовых отходов мегаполисов как перспективные виды для фиторемедиации // Физиология растений. 2008. Т. 55. № 3. С. 453–463.
Уланова Н.Г. Вейник наземный // Биологическая флора Московской области. М.: Изд-во МГУ, 1995. Вып. 10. С. 4–19.
Lupardus R.C., McIntosh A.C.S., Janz A., Farr D. Succession after reclamation: identifying and assessing ecological indicators of forest recovery on reclaimed oil and natural gas well pads // Ecological Indicators. 2019. V. 106. P. 105515. https://doi.org/10.1016/j.ecolind.2019.105515
Prévosto B., Kuiters L., Bernhardt-Römermann M. et al. Impacts of land abandonment on vegetation: Successional pathways in european habitats // Folia Geobotanica, Springer Verlag. 2011. V. 46 (4). P. 303–325. https://doi.org/10.1007/s12224-010-9096-z
Bourgeois B., Vanasse A., Gonzalez E. et al. Threshold dynamics in plant succession after tree planting in agricultural riparian zones // Journal of Applied Ecology. 2016. V. 53(6). P. 1704–1713. https://doi.org/10.1111/1365-2664.12675
Дополнительные материалы отсутствуют.