

Экология, 2022, № 6, стр. 430-437
О некоторых закономерностях распределения эпифитных лишайников по стволу дерева
Е. Э. Мучник a, *, **, Е. Ю. Благовещенская b
a Институт лесоведения Российской академии наук
143030 Московская обл., Одинцовский г.о., п/о Успенское, ул. Советская, 21, Россия
b Московский государственный университет имени М.В. Ломоносова
119234 Москва, Ленинские горы, 1, стр. 12, Россия
* E-mail: emuchnik@outlook.com
** E-mail: eu.muchnik@yandex.ru
Поступила в редакцию 17.03.2022
После доработки 27.04.2022
Принята к публикации 25.05.2022
- EDN: KLVWIS
- DOI: 10.31857/S0367059722060117
Аннотация
Изучено распределение эпифитных лишайников по высоте ствола для свежеповаленных берез и елей на территории природного заказника в Московской области. Показано, что положение на стволе дерева значимо влияет на число выявляемых видов лишайников, которое монотонно возрастает с увеличением высоты. Для обеих пород форофитов характерно высокое разнообразие лихенобиоты в нижней и верхней частях ствола (и особенно в кроне). От основания вверх по стволу происходит смена доминирующих видов лишайников. Отмечено, что именно в верхней части стволов сосредоточены находки охраняемых в Московской области видов родов Bryoria и Usnea.
В процессе большинства лихенологических исследований эпифитные лишайники собирают и учитывают лишь до высоты 2–2.5 м на стволе дерева. Тем самым значительная часть информации о биологическом разнообразии лишайников остается недоступной – по разным оценкам [1‒3] не учитывается от половины до двух третей от общего видового богатства лихенобиоты. Однако подобные данные могут быть очень важны как для оценки общего биоразнообразия лесных сообществ, так и для целей выявления редких видов и проведения биоиндикационных исследований.
Наличие неоднородного высотного распределения лишайников по стволу форофита отмечалось неоднократно [4‒11]. Тем не менее подробных исследований разнообразия лишайников по градиенту высоты по-прежнему немного. Например, при обследовании хвойных лесов Америки показана смена преобладающих формаций с мохового на лишайниковый покров при движении вверх по стволу [12‒14], кроме того, в старовозрастных лесах доминирующие виды родов Hypogymnia и Platismatia произрастают только в верхней части крон [12]. Также в кроне деревьев часто оказываются сосредоточены находки редких и охраняемых видов лишайников [3, 12, 13]. Показано [2], что состав лихенобиоты верхней части кроны деревьев в еловых и сосновых лесах гораздо более информативен для оценки пылевого загрязнения.
Однако детальных исследований разнообразия эпифитных лишайников на разной высоте ствола дерева проводилось крайне мало [6], что связано с объективными методическими трудностями: для обследования деревьев по всей высоте вплоть до ветвей кроны необходимы специальное дорогостоящее оборудование либо вырубка “модельных” деревьев, причем последний метод наносит ощутимый вред лесному сообществу, особенно в случае достаточной для статистической обработки выборки. Наилучшие условия для исследований распределения лишайников по высоте ствола складываются на недолгое время после ветровалов, когда довольно много свежих валежных деревьев разных пород и классов возраста оказываются доступными для сборов эпифитной лихенобиоты по всем высотам. Цель данной работы ‒ выявление закономерностей распределения эпифитных лишайников по высоте ствола на примере свежеповаленных деревьев на территории заказника “Звенигородская биологическая станция МГУ и карьер “Сима””. Рабочая гипотеза: видовое разнообразие и встречаемость эпифитных лишайников на разных высотах одного и того же форофита значимо различаются; в распределении разнообразия эпифитных лишайников по стволам форофитов разных видов предполагались некоторые общие тенденции.
МАТЕРИАЛ И МЕТОДЫ
Район исследований. Комплексный заказник регионального значения “Звенигородская биостанция МГУ и карьер “Сима”” (далее ЗБС МГУ) площадью 1116.3 га расположен на западе Московской области на правом берегу р. Москвы (рис. 1). Климат умеренно континентальный, средняя месячная температура колеблется от –10.6°С в январе до +17.2°С в июле, сумма осадков за год превышает 700 мм. В заказнике сохраняются типичные для региона естественные условия и процессы, а режим эксплуатации максимально приближен к заповедному [15]. Территория относится к подзоне хвойно-широколиственных лесов. Растительный покров очень разнообразен: хвойные сосновые и еловые древостои сочетаются с дубово-липовыми насаждениями, ольшаниками, болотами и лугами. Основная лесообразующая порода – ель европейская (Picea abies (L.) H. Karst.) – в последние годы выпадала вследствие деятельности короеда-типографа (Ips typographus L.). Кроме того, на фоне поражения короедом в 2016 и 2017 гг. леса заказника были повреждены сильным ветровалом.
Сбор и камеральная обработка материала. В течение двух полевых сезонов (2016–2017 гг.) на территории комплексного заказника ЗБС МГУ проведены сборы эпифитных лишайников на свежих валежных деревьях в субнеморальном ельнике возрастом более 100 лет. Всего обследовано 10 поваленных стволов ели (Picea abies) и 13 стволов березы (Betula pendula Roth) в нескольких кварталах заказника (см. рис. 1, врезка).
Сбор лишайников осуществляли у основания ствола, на уровне 5 м высоты и далее с шагом в 5 м. Для березы отбор лишайников проводили вплоть до начала разветвления ствола, далее отдельно учитывали лишайники кроны (как основных ветвей, так и мелких веточек); для ели отбор проводили аналогично, с поправкой на то, что ветвления стволовой части не происходит, в качестве начала кроны рассматривали первые живые ветви (мутовку или часть мутовки) [16]. Собранный материал (289 образцов) определяли с помощью общепринятых лихенологических методик [17]. Номенклатура упомянутых видов лишайников соответствует современной сводке [18].
Анализ данных. Для сравнения видового состава лихенобиот на ели и березе использовали коэффициент Съеренсена (KS) [19]. Поскольку оказалось, что состав лишайников на ели и березе заметно различается, то дальнейшие анализы проводили для этих пород раздельно. Для выявления влияния фактора положения на стволе дерева на число видов лишайников был проведен однофакторный дисперсионный анализ (ДА) для отдельных пород деревьев. На основании коэффициента KS была составлена матрица расстояний, где в качестве расстояния использовали значение 1 – KS, и проведен кластерный анализ; в качестве метода объединения использовали метод “дальнего соседа” (метод полной связи).
Обилие лишайников не учитывали, но для оценки биоразнообразия был введен дополнительный косвенный показатель обилия, вычисляемый следующим образом: наличие вида на определенной высоте конкретного дерева принимали за “находку”. Таким образом, каждому виду на конкретной породе дерева могло быть приписано некоторое число “находок” ‒ от 1 в случае однократного обнаружения до суммарного количества всех проанализированных высот по всем деревьям данной породы (75 для березы и 48 для ели). Данное число и рассматривали как показатель абсолютного обилия вида. Для вычисления относительного обилия (pi) число “находок” конкретного вида делили на сумму “находок” всех видов по данной породе дерева (181 для березы и 89 для ели). На основе полученных данных строили графики распределения обилия видов. Используя аналогичный метод для отдельных высот деревьев определенной породы, вычисляли индекс разнообразия Шеннона (H) и индекс доминирования Симпсона (D) [19] для каждой высоты.
Анализ проводили в программах Microsoft Excel 2010 и Statistica 8.0.
РЕЗУЛЬТАТЫ
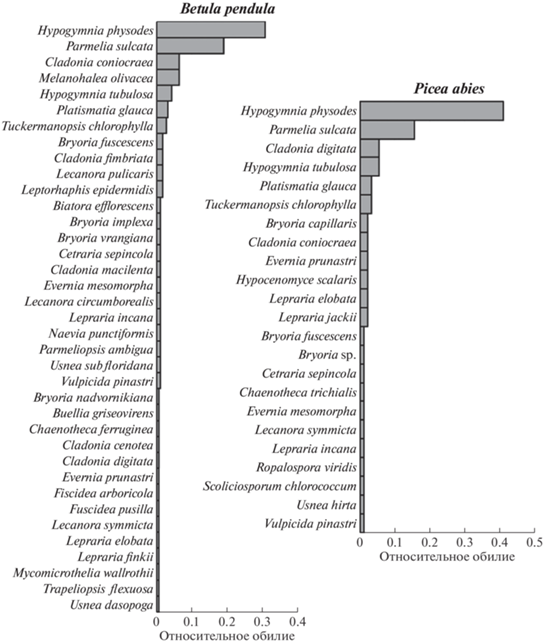
Суммарно на березе выявлено 37 видов эпифитных лишайников, на ели – 23. Коэффициент сходства между породами составил 51%. В качестве преобладающих видов отмечены: на березе ‒ Hypogymnia physodes, Parmelia sulcata, Cladonia coniocraea и Melanohalea olivacea, на ели ‒ H. physodes и P. sulcata, Cladonia digitata и H. tubulosa (рис. 2).
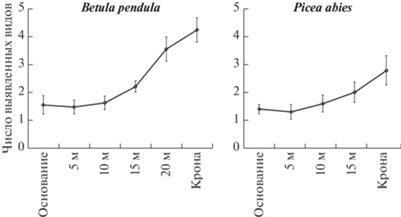
Однофакторный ДА показал значимость фактора положения на стволе дерева для числа выявляемых видов как на березе (F = 65.6; df = 6; p < < 0.01), так и на ели (F = 25.6; df = 6; p < 0.01). Следовательно, лихенобиота на разном уровне ствола характеризуется различным видовым богатством. Суммарно наибольшее число видов выявляется в кроне (16 видов для березы и 10 для ели). Аналогичные данные для стволов деревьев достаточно неоднородны, хотя можно сказать, что первые 10 м ствола дают схожие значения суммарного числа видов (8–10 для березы и 5–7 для ели). Более наглядную картину показал анализ числа видов, выявляемых в среднем на одном дереве: среднее число видов монотонно возрастает в направлении от основания к кроне, что особенно выражено у березы (рис. 3).
Рис. 3.
Среднее число выявляемых видов лишайников на Betula pendula и Picea abies (полосы погрешности указывают на ошибку среднего).
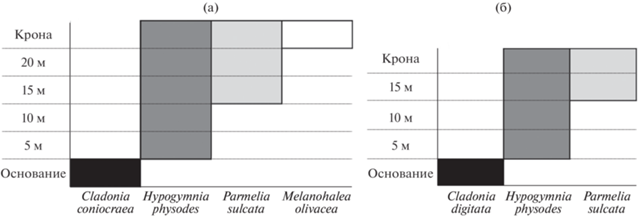
Разнообразие лишайников заметно различается для разных уровней высот на стволах и берез, и елей (табл. 1, рис. 4, 5). При движении от основания вверх по стволу происходит смена преобладающих видов. Это можно установить, если для каждого уровня высоты выделить виды, обнаруживаемые на данной высоте более чем на половине исследованных деревьев (рис. 6). Основным видом для основания ствола березы является C. coniocraea, который уже на высоте 5 м встречается в 2 раза реже и лишь один раз отмечен на высоте 10 м. По всей высоте ствола встречается Hypogymnia physodes, достигая 100% встречаемости на уровне 15 м и несколько снижая свое присутствие далее. Только с 10 м отмечен Parmelia sulcata, но далее встречается на всех высотах обследованных берез. Наконец, Melanohalea olivacea обнаружен только с высоты 20 м, но при этом преобладает в ветвях кроны. Исключительно в кроновой части на березе отмечены следующие виды: Cetraria sepincola, Evernia prunastri, Lecanora circumborealis, L. pulicaris, L. symmicta, Naevia punctiformis, Usnea dasypoga, Vulpicida pinastri. У ели в основании ствола чаще встречаются виды рода Cladonia, а в верхней части в число доминантов входит Parmelia sulcata.
Таблица 1.
Значения индекса разнообразия Шеннона (H) и индекса доминирования Симпсона (D) для эпифитной лихенобиоты на разной высоте ствола
| Индекс | Порода | Уровень высоты | |||||
|---|---|---|---|---|---|---|---|
| основание | 5 м | 10 м | 15 м | 20 м | крона | ||
| H | Береза | 2.90 | 2.25 | 2.39 | 1.44 | 3.21 | 3.33 |
| Ель | 2.50 | 1.51 | 2.09 | 1.63 | 2.87 | ||
| D | Береза | 0.18 | 0.32 | 0.31 | 0.42 | 0.15 | 0.14 |
| Ель | 0.22 | 0.50 | 0.35 | 0.38 | 0.18 | ||
Рис. 4.
Число видов лишайников, которые были выявлены на Betula pendula только на определенном уровне высоты.
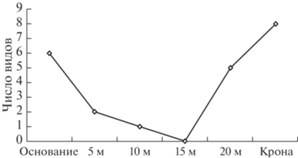
Рис. 5.
Дендрограмма сходства видовых составов лишайников на разной высоте ствола Betula pendula (а) и Picea abies (б). В качестве метода объединения использовали метод “дальнего соседа” (метод полной связи). По оси абсцисс: 1–KS.
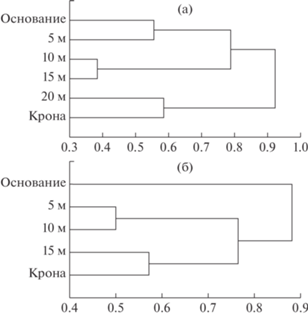
Рис. 6.
Диаграмма распределения доминантных лишайников на разной высоте стволов Betula pendula (а) и Picea abies (б); пояснения в тексте.
Для ели приведены данные по соответствию известных предпочтений лишайников к условиям освещенности и на какой высоте мы их обнаруживаем (табл. 2).
Таблица 2.
Вертикальное распределение некоторых лишайников по стволам елей (виды расположены в порядке возрастания требований к условиям освещенности ‒ приведен диапазон по 5-балльной шкале [20])
| Вид лишайника | Экологические требования по фактору освещенности местообитания | Диапазоны высот на стволах ели | |||
|---|---|---|---|---|---|
| основание | 5–10 м | 15 м | крона | ||
| Chaenotheca trichialis | 1–3 | + | |||
| Lepraria jackii | 2–3 | + | |||
| Scoliciosporum chlorococcum | 2–3 | + | |||
| Lepraria elobata | 2–4 | + | + | ||
| Lepraria incana | 2–4 | + | |||
| Hypogymnia tubulosa | 3 | + | + | ||
| Hypogymnia physodes | 3–4 | + | + | + | + |
| Cladonia coniocraea | 3–4 | + | |||
| Cladonia digitata | 3–4 | + | |||
| Ropalospora viridis | 3–4 | + | |||
| Lecanora symmicta | 3–4 | + | |||
| Tuckermanopsis chlorophylla | 3–4 | + | + | ||
| Parmelia sulcata | 3–5 | + | + | + | |
| Platismatia glauca | 3–5 | + | + | ||
| Vulpicida pinastri | 3–5 | + | |||
| Hypocenomyce scalaris | 3–5 | + | |||
| Bryoria capillaris | 3–5 | + | + | ||
| Evernia prunastri | 3–5 | + | + | ||
| Bryoria fuscescens | 3–5 | + | |||
| Usnea hirta | 4–5 | + | |||
| Cetraria sepincola | 4–5 | + | |||
| Evernia mesomorpha | 4–5 | + | |||
ОБСУЖДЕНИЕ
Выявленное различие лихенобиот березы и ели обусловлено разными свойствами коры, что соответствует данным некоторых авторов [21]. Так как оба форофита принадлежат группе деревьев с “кислой” корой (рН < 4.7) [22], в различии видового состава эпифитных лишайников предположительно бóльшую роль играют такие факторы, как структура поверхности и стабильность коры [4], а также микроклимат в кронах и подкроновой части [23].
Несмотря на то, что в нижней части дерева в среднем встречается меньше лишайников (см. рис. 3), видовой состав от дерева к дереву может меняться, что и дает сравнительно высокие значения суммарного видового богатства лихенобиоты. Таким образом, видовой состав лишайников в основании ствола варьирует сильнее, чем в кроне. Так, для высоты 15 м на березе отмечено низкое разнообразие лишайников (H = 1.44) и максимальное значение индекса доминирования (D = 0.42), т.е. лихенобиота на данном уровне представлена преимущественно самыми распространенными лишайниками. В целом разнообразие лихенобиоты выше у березы, что в значительной степени связано с бóльшим числом выявленных на данном форофите видов. Для обеих пород деревьев характерно высокое разнообразие лишайников в нижней и верхней частях ствола (и особенно в кроне) и некоторое “падение” в средней части. Особенно наглядно это иллюстрирует число видов лишайников, обнаруженных только на конкретной высоте ‒ “маркеров высоты” (см. рис. 4).
Наблюдаемая вертикальная зональность распределения лишайников по стволу дерева обусловлена различиями в микроклиматических условиях ‒ влажности, освещенности, химических и физических свойств покровных тканей дерева [4, 21‒24]. Если рассматривать стволы березы и ели, то на первое место, вероятно, выходит освещенность с поправкой на особенности микроклимата именно комлевой части стволов. В обоих случаях верхняя часть ствола кластеризуется вместе с кроной (см. рис. 5), а в нижней части стволов ели и березы мы наблюдаем некоторые различия. Для ели уровень основания ствола формирует отдельную кладу, что обусловлено произрастанием видов рода Cladonia только в основании ствола. Напротив, характер коры березы в сочетании с тем, что ствол исходно мог иметь не жестко вертикальное положение, позволяет видам Cladonia довольно высоко подниматься по стволу, обеспечивая объединение позиций основания и высоты в 5 м.
То, что Parmelia sulcata тяготеет к верхней части деревьев (см. рис. 6), отмечали и другие авторы [4, 6, 7], и в ряде случаев она действительно отмечается как доминант [4, 7]. Доминирующие позиции вида Hypogymnia physodes также широко известны [4, 7 и др.]. Безусловно, этот вид в умеренно континентальном климате хвойно-широколиственной подзоны более толерантен к фактору освещенности: его интервалы в баллах для этого фактора должны составлять не 3–4 (как в Средиземноморье ‒ по [20]), а минимум 2–4, максимум 2–5 баллов. Шире этот интервал (правда, только в сторону увеличения освещенности) и у H. tubulosa, составляя не 3 балла, а минимум 3–4, максимум 3–5. Высокие требования к освещенности у этого вида послужили причиной того, что при маршрутных лихенологических исследованиях его собирали нечасто, долгое время он считался редким в Московской области и был даже занесен в список охраняемых [25]. Тем не менее, как видно на рис. 2, данный вид входит в состав наиболее часто встречающихся, но только начиная с высоты 15 м на ели (см. табл. 2), а на березе – с высоты 10 м. В целом экологические предпочтения некоторых видов в отношении фактора освещенности, очевидно, варьируют в зависимости от природно-климатической зоны.
В основании обеих пород деревьев остаются виды рода Cladonia, относящиеся к числу сравнительно теневыносливых. Их отношение к освещенности составляет 2–4 балла [20], а если говорить о C. digitata в условиях Московской области, где он предпочитает очень затененные и затененные условия, более справедлива оценка в 1–3 балла. Характерно, что более теневыносливый вид рода Cladonia отмечен для ели, имеющей меньшую сквозистость кроны, что приводит к заметному снижению освещенности в основании ствола.
Отметим, что охраняемые в настоящее время на территории Московской области [26] виды как на березе, так и на ели отмечены, начиная с высоты 10 м (Bryoria fuscescens) и 20 м (B. capillaris, B. implexa, B. nadvornikiana, B. vrangiana, Usnea dasopoga, U. hirta, U. subfloridana). Это коррелирует с данными [3, 13, 27], показывающими, что в кроне деревьев часто оказываются сосредоточены находки редких и охраняемых видов лишайников.
ЗАКЛЮЧЕНИЕ
Обследование позволило выявить некоторые закономерности вертикального распределения эпифитных лишайников на стволах берез и елей. Несмотря на небольшую степень сходства видового состава эпифитов березы и ели, обусловленную различиями физических и химических свойств коры, в распределении видового богатства эпифитных лишайников по высоте стволов обеих пород форофитов наблюдаются некоторые общие тенденции.
Подтвердилась гипотеза о значимом различии видового разнообразия и встречаемости эпифитных лишайников на разных высотах одного и того же форофита. Для обеих пород форофитов характерно высокое разнообразие лихенобиоты в нижней и верхней частях ствола (и особенно в кроне), а в средней части наблюдается снижение этого показателя. Отметим, что в нижней части каждого конкретного ствола обычно развивается меньше видов лишайников, чем в верхней, но при этом видовой состав от дерева к дереву сильно варьирует, что и дает сравнительно высокие значения суммарного видового богатства основания ствола.
От основания вверх по стволу происходит смена доминирующих видов. На березе доминанты меняются в следующем порядке: Cladonia coniocraea (основание) ‒ Hypogymnia physodes (5‒10 м) ‒ H. physodes, Parmelia sulcata (10‒20 м) ‒ H. physodes, P. sulcata, Melanohalea olivacea (крона). Аналогичный ряд доминантов для ели выглядит несколько иначе: Cladonia digitata (основание) ‒ Hypogymnia physodes (5‒10 м) ‒ H. physodes, Parmelia sulcata (15 м ‒ крона). На смену доминантов, по-видимому, наибольшее влияние оказывает фактор освещенности: эта зависимость лучше видна в распределении по высоте лишайников ели, имеющей вечнозеленую крону. Находки H. tubulosa на стволах ели исключительно в верхней части (начиная с 15 м и в ветвях кроны) свидетельствуют о большей “фотофильности” этого вида в умеренно континентальном климате хвойно-широколиственной подзоны, чем в Средиземноморье.
Существенным для природоохранной деятельности фактом является сосредоточение в верхней части стволов и кронах находок охраняемых в регионе видов родов Bryoria и Usnea, что необходимо учитывать при определении их реальной встречаемости и численности популяций в Московской области.
Работа выполнена в рамках темы государственного задания Института лесоведения РАН АААА-А19-119053090074-7 “Структура, динамика и производительность естественных и искусственных лесных сообществ в Центре Русской равнины” и темы государственного задания МГУ № 121032300081-7. Авторы признательны к.б.н. И.Д. Инсаровой за ценные замечания и рекомендации при подготовке статьи.
Авторы заявляют об отсутствии конфликта интересов.
Настоящая статья не содержит исследований с участием людей или животных в качестве объектов исследования.
Список литературы
Boch S., Müller J., Prati D. et al. Up in the tree – the overlooked richness of bryophytes and lichens in tree crowns // PLoS ONE. 2013. V. 8. №12. P. e84913. https://doi.org/10.1371/journal.pone.0084913
Marmor L., Tõrra T., Saag L. et al. Lichens on Picea abies and Pinus sylvestris – from tree bottom to the top // Lichenologist. 2013. V. 45. №1. P. 51–63. https://doi.org/10.1017/S0024282912000564
Kiebacher T., Keller C., Scheidegger C., Bergamini A. Hidden crown jewels: the role of tree crowns for bryophyte and lichen species richness in sycamore maple wooded pastures // Biodivers. Conserv. 2016. V. 25. P. 1605–1624. https://doi.org/10.1007/s10531-016-1144-4
Hale M.E. Vertical distribution of cryptogams in a red maple swamp in Connecticut // Bryologist. 1965. V. 68. № 2. P. 193–197.
Holien H. The lichen flora on Picea abies in a suboceanic spruce forest area in Central Norway with emphasis on the relationship to site and stand parameters // Nordic J. Bot. 1997. V. 17. № 1. P. 55–76.
Caruso A., Thor G. Importance of different tree fractions for epiphytic lichen diversity on Picea abies and Populus tremula in mature managed boreonemoral Swedish forests // Scand. J. Forest Res. 2007. V. 22. № 3. P. 219–223. https://doi.org/10.1080/02827580701346031
Fritz Ö. Vertical distribution of epiphytic bryophytes and lichens emphasizes the importance of old beeches in conservation // Biodivers. Conserv. 2009. V. 18. № 2. P. 289–304. https://doi.org/10.1007/s10531-008-9483-4
Теплых A.A. Пространственная и возрастно-виталитетная структура популяции лишайника Pseudevernia furfuracea (L.) Zopf в условиях верхового болота // Научные груды Государственного природного заповедника “Большая Кокшага”. 2008. Вып. 3. С. 143–158.
Lie M.H., Arup U., Grytnes J.-A., Ohlson M. The importance of host tree age, size and growth rate as determinants of epiphytic lichen diversity in boreal spruce forests // Biodivers. Conserv. 2009. V. 18. № 13. P. 3579–3596. https://doi.org/10.1007/s10531-009-9661-z
Яцына А.П. Структура эпифитных лишайниковых сообществ Pinus sylvestris L. в Беларуси // Веснiк ВДУ. 2013. № 1. С. 45–49.
Nadyeina O., Dymytrova L., Naumovych A. et al. Distribution and dispersal ecology of Lobaria pulmonaria in the largest primeval beech forest of Europe // Biodivers. Conserv. 2014. V. 23. № 13. P. 3241–3262. https://doi.org/10.1007/s10531-014-0778-3
McCune B. Gradients in epiphyte biomass in three Pseudotsuga-Tsuga forests of different ages in Western Oregon and Washington // Bryologist. 1993. V. 96. № 3. P. 405–411.
Rosso A.L., McCune B., Rambo T.R. Ecology and conservation of a rare, old-growth associated canopy lichen in a silvicultural landscape // Bryologist. 2000. V. 103. № 1. P. 117–127.
Ellyson W.J.T., Sillet S.C. Epiphyte communities on Sitka spruce in an old-growth redwood forest // Bryologist. 2003. V. 106. № 2. P. 197–211. https://doi.org/10.1639/0007-2745(2003)106[0197: ECOSSI]2.0.CO;2
Сборник материалов X рабочего совещания комиссии по изучению макромицетов и VI микологической школы-конференции “Мицелиальный образ жизни и эколого-трофические группы грибов” / Ред. Воронина Е.Ю., Дьяков М.Ю., Попов Е.С. М., 2014. 82 с.
Анучин Н.П. Лесная таксация. Изд. 5-е, доп. М.: Лесн. пром-сть, 1982. 552 с.
Степанчикова И.С., Гагарина Л.В. Сбор, определение и хранение лихенологических коллекций // Флора лишайников России: Биология, экология, разнообразие, распространение и методы изучения лишайников. М.; СПб.: Тов-во науч. изд. КМК, 2014. С. 204–219.
Westberg M., Moberg R., Myrdal M. et al. Santesson’s checklist of fennoscandian lichen-forming and lichenicolous fungi. Uppsala University: Museum of Evolution, 2021. 933 p.
Миркин Б.М., Розенберг Г.С., Наумова Л.Г. Словарь понятий и терминов современной фитоценологии. М.: Наука, 1989. 223 с.
Nimis P.L., Martellos S. ITALIC ‒ The information system on Italian Lichens. Version 6.0. 2021. University of Trieste, Dept. of Biology. URL: http://dryades. units.it/italic, accessed on 27.12.2021.
Peciar V. Epiphytische Moosgesellschaften der Slowakei // Acta Fac. Rer. Nat. Univ. Comeniae. Botanika. 1965. V. 9. № 12. P. 371–470.
Du Rietz G.E. Om fattigbark ‒ och rikbarsamhällen // Svensk Bot. Tidskr. 1945. V. 39. № 1. P. 147‒150.
Campbell J., Coxson D.S. Canopy microclimate and arboreal lichen loading in subalpine spruce-fir forest // Can. J. Bot. 2001. V. 79. № 5. P 537–555. https://doi.org/10.1139/cjb-79-5-537
Корчиков Е.С. Лишайники Самарской Луки и Красносамарского лесного массива. Самара: “Самарский университет”, 2011. 320 с.
Красная книга Московской области. 2-е изд. / Отв. ред. Варлыгина Т.И., Зубакин В.А., Соболев Н.А. М.: Тов-во науч. изд. КМК, 2008. 828 с.
Красная книга Московской области. 3-е изд., переработ. и дополн. / Отв. ред. Варлыгина Т.И., Зубакин В.А., Никитский Н.Б., Свиридов А.В. Московская обл.: ПФ “Верховье”, 2018. 810 с.
McCune B., Rosentreter R., Ponzetti J.M., Shaw D.C. Epiphyte habitats in an old conifer forest in Western Washington, U.S.A. // Bryologist. 2000. V. 103. № 3. 417–427. https://doi.org/10.1639/0007-2745(2000)103[0417: EHIAOC]2.0.CO;2
Дополнительные материалы отсутствуют.