

Экология, 2023, № 1, стр. 46-57
Средовые предпочтения рыжей полевки (Clethrionomys glareolus Schreber, 1780) в отличающихся биотопических условиях на охраняемой территории Среднего Урала
Л. Е. Лукьянова *
ФГБУН Институт экологии растений и животных УрО РАН
620144 Екатеринбург, ул. 8 Марта, 202, Россия
* E-mail: lukyanova@ipae.uran.ru
Поступила в редакцию 30.11.2021
После доработки 20.05.2022
Принята к публикации 24.05.2022
- EDN: DBKULM
- DOI: 10.31857/S0367059722060099
Аннотация
Изучали средовые предпочтения рыжей полевки на охраняемой территории Среднего Урала в отличающихся биотопических условиях, сформированных в результате последствий природных катастрофических явлений. По значениям обилия населения оценивали “успешность” существования вида в четырех биотопах с разной степенью нарушенности ветровально-пирогенным воздействием. Выявлен статистически высокозначимый уровень отличий сравниваемых биотопов по шести микросредовым переменным, оценивающим кормозащитные условия местообитаний животных. В целом биотопическая изменчивость оказалась выше хронографической (межгодовой) изменчивости. За многолетний период исследования рыжая полевка преобладала по численности в ненарушенном биотопе, который в ряду исследуемых биотопов выделялся высокими значениями микросредовых показателей, оценивающих преимущественно кормовые ресурсы местообитаний животных – покрытие участков кустарником и травянистой растительностью. Условия обитания в ненарушенном природными катастрофическими факторами биотопе в наибольшей степени отвечают средовым предпочтениям рыжей полевки.
Знание особенностей выбора среды разными видами позволяет понять причины распределения организмов в определенном пространстве и времени, а также прогнозировать состояние популяций в измененных условиях, вызванных различными нарушениями [1]. Степень благополучия существования вида в природной среде, его биологический успех и приспособительную пластичность отражают такие популяционные показатели, как обилие и пространственная структура [2, 3]. В связи с этим выявление средовых предпочтений видов на основе изучения их популяционной динамики в отличающихся биотопических условиях является важной экологической проблемой.
Видовые предпочтения проявляются на макро- и микросредовом уровнях [1, 4–7]. В выборе условий обитания мелкими млекопитающими решающую роль играет микросредовая структура биотопов, поскольку жизнедеятельность многих видов в конкретные периоды их жизненного цикла протекает на участках, несоизмеримых с территорией всей популяции [8]. На выбор среды влияет комплекс факторов, включая доступность пищи, пригодность убежищ, наличие конкурентов, риск хищничества, паразитизма и болезней [9–19]. Средовые предпочтения грызунов в нарушенных природными или антропогенными воздействиями местообитаниях могут меняться, так как в дестабилизированной среде реализуются видовые особенности, не проявляющиеся в стабильных условиях [20]. Мозаичность среды, вызванная последствиями природных катастрофических явлений, отражается на характере использования местообитаний мелкими млекопитающими, что приводит к пространственному перераспределению обилия видов [20–27]. Выявлено, что при выраженной мозаичности среды в короткие временны́е интервалы важнейшим элементом адаптивной стратегии подвижных организмов становится дифференцированное использование жизненного пространства [28]. Это имеет большой биологический смысл, поскольку с одной стороны оно определяет наиболее эффективное использование кормовых, защитных, микроклиматических и других ресурсов среды, а с другой – служит основой устойчивого поддержания необходимого уровня внутривидовых (внутрипопуляционных) контактов между особями [29].
Рыжая полевка (Clethrionomys glareolus Schreber, 1780) – представитель рода лесных полевок (Clethrionomys, в современной систематике чаще используют название Myodes), является фоновым видом лесов умеренного климата. В таежной зоне наиболее высокой численности она достигает в ягодных ельниках и граничащих с ними вырубках, обычна в пойменных лесах, повсеместно избегает лесов с сомкнутым древостоем, населяя светлые вторичные леса [30]. Предпочтительными для рыжей полевки являются местообитания с богатыми и разнообразными кормовыми ресурсами (высокий урожай семян и ягод, обильная вегетация трав) и благоприятными защитными условиями, обеспечивающими зверьков широкими возможностями устройства жилищ. На распределение численности вида оказывают влияние такие характеристики среды, как состояние подлеска (развитый подрост, обилие кустарников), состав древостоя, степень захламленности участков (веточный опад, упавшие мертвые деревья) [31–37].
Ранее мы показали [38], что выбор среды рыжей полевкой в экологически контрастных биотопических условиях на двух охраняемых территориях Урала имеет отличительные особенности: в биотопах нарушенной ветровалом и пожаром территории Среднего Урала видовые предпочтения связаны со стадиями посткатастрофических восстановительных сукцессий, а в стабильных условиях местообитаний на ненарушенной территории Северного Урала выбор среды в большей степени обусловлен ландшафтной структурой биотопов.
Цель настоящего исследования – выявить средовые предпочтения рыжей полевки в отличающихся по степени нарушенности природными катастрофическими факторами биотопических условиях. Основные решаемые задачи: 1) изучить многолетнюю динамику обилия рыжей полевки в биотопах с разной степенью природных нарушений; 2) сравнить микросредовую структуру по семи основным характеристикам, отражающим кормозащитные условия местообитаний грызунов; 3) оценить межгодовую изменчивость микросредовых характеристик; 4) выявить связь обилия рыжей полевки с параметрами микросреды в отличающихся биотопических условиях.
МАТЕРИАЛ И МЕТОДЫ
В исследовании использованы рыжие полевки, отловленные в период 2013–2021 гг. на территории Висимского государственного природного биосферного заповедника (Свердловская область, Средний Урал, 57°19′–57°31′ с.ш. и 59°20′–59°50′ в.д.). В широком спектре местообитаний Clethrionomys glareolus на Среднем Урале отмечены разнообразные биотопы, включая послелесные растительные формации на вырубках и гарях, а также целый ряд антропогенных местообитаний [39]. Динамику численности населения рыжей полевки и ее средовые предпочтения изучали в отличающихся биотопических условиях, сформированных вследствие природных катастрофических явлений: ветровала в 1995 г. и последующего пожара, возникшего в 1998 г. от молнии во время “сухой грозы”, охватившего лишь часть исследуемой ветровальной территории. В 2010 г. в заповеднике по той же причине случился второй пожар, от которого первично пострадала не нарушенная предыдущим пирогенным воздействием часть ветровальной территории, а также горевшая ранее, которая в итоге оказалась дважды пирогенно нарушенной ветровальной территорией.
Для исследования были отобраны четыре биотопа с разной степенью нарушенности природными катастрофическими факторами: ветровалом и двумя пожарами (биотоп I), ветровалом и одним пожаром (биотоп II), частично (по периферии участка) ветровалом и одним пожаром (биотоп III), и местообитание, не нарушенное внешними неблагоприятными воздействиями (биотоп IV). До природных катастрофических явлений биотопы I и II входили в состав пихто-ельников липнякового коренного и условно-коренного, а также мелкотравно-вейникового условно-коренного (с примесью березы и осины). В настоящее время эти биотопы являются ветровально-гаревыми лесными сообществами на разных стадиях восстановительных сукцессий: биотоп I представляет собой послепожарное вейниково-кипрейное сообщество под разреженным древесным пологом, биотоп II − послепожарное березово-пихтово-еловое редколесье. Биотоп III входит в состав пихтово-елово-березового большехвостоосоково-липнякового леса, биотоп IV расположен в коренном высокотравно-папоротниковом пихтово-еловом лесном сообществе. Все изучаемые биотопы имеют ландшафтное сходство, расположены в привершинной части невысоких гор: биотопы I–III занимают пологий склон г. Липовый Сутук (495 м над ур. м.), а биотоп IV находится на склоне г. Малый Сутук (560 м над ур. м.).
Состояние населения рыжей полевки оценивали по уровню ее обилия (численности). Для отлова животных использовали стандартный метод ловушко-линий, показатель обилия рассчитывали по числу отловленных особей за первые 5 сут в пересчете на 100 ловушко-суток (ос/100 лов.-сут). В каждом биотопе в конце летнего сезона одновременно размещали по 50 проволочных капканчиков в линии длиной 500 м, время экспозиции составляло 5 сут. Проверку осуществляли ежедневно в утренние часы. Каждая ловушка на протяжении всего многолетнего периода наблюдений имела неизменное местоположение, была снабжена постоянным номером, что важно для учета поимок животных и анализа их пространственного распределения. Общее число использованных в работе рыжих полевок составило 532 особи.
Вокруг каждой ловушки, расположенной в центре квадрата со сторонами 3.33 м (площадь – 10 м2), дважды за исследуемый период (в 2013 и 2017 гг.) в августе проводили количественное описание характеристик микросреды. Оценивали параметры, характеризующие кормозащитные условия обитания рыжей полевки на основе методики, предложенной О.А. Лукьяновым и Г. Буяльской [8], с незначительными изменениями. Измеряли площадь покрытия микроучастков: 1) мхом (MC), 2) травянистой растительностью (HC), 3) кустарником (CS), 4) основаниями стволов живых деревьев (TC), 5) основаниями стволов стоящих сухих деревьев и пней (SC), 6) стволами лежащих деревьев (LC) и 7) подростом древесных пород (AU) высотой не более 1 м (табл. 1).
Таблица 1.
Характеристики, используемые для анализа микросреды местообитаний рыжей полевки
| Характеристика | Условное обозначение |
|---|---|
| Площадь участка (м2), покрытая: | |
| 1) мхом 2) травянистой растительностью 3) кустарником 4) стволами живых деревьев 5) стволами стоящих сухих деревьев и пней 6) лежащими стволами деревьев 7) подростом древесных пород, экз/м2 |
MC HC CS TC SC LC AU |
Выбранная нами периодичность проведения количественных описаний объясняется тем, что подавляющее число данных характеристик микросреды подвержено незначительным межгодовым флуктуациям, и они отражают ее статичные свойства [8]. При отборе микросредовых переменных учитывали их “физические” свойства для рыжей полевки. Исходя из “функциональности” отобранных семи переменных микросреды, все они в целом характеризуют кормозащитные условия местообитаний исследуемого вида. Отмечено, что микросредовые переменные, оценивающие травянистый и кустарниковый покровы, отражают в большей степени кормовые характеристики местообитаний. Наряду с этим признаки, связанные с покрытием участков мхом, живыми деревьями и древесным подростом, характеризуют кормозащитные свойства, но с акцентом на защитный аспект, а в “чистом” виде защитные условия для рыжей полевки обеспечивают характеристики, оценивающие покрытие участков пнями и сухими деревьями [8].
Травянистый ярус в исследуемых нами четырех биотопах в целом однотипен, в его состав входят вейники Лангсдорфа (Calamagrostis langsdorffii) и тупочешуйный (Calamagrostis obtusata), иван-чай узколистный (Chamaenerion angustifolium), которые доминируют в послепожарных сообществах (биотопы I и II). В составе травостоя биотопа III широко представлена осока большехвостая (Carex macroura), встречаются сныть обыкновенная (Aegopodium podagraria), а также хвощ лесной (Equisetum sylvaticum). В травостое ненарушенного коренного высокотравно-папоротникового пихто-ельника (биотоп IV) доминируют папоротники, среди которых преобладает щитовник (Dryopteris assimilis). Кустарниковый подлесок изучаемых биотопов сформирован в основном малиной обыкновенной (Rubus idaeus) и сахалинской (Rubus matsumuranus), а также шиповником иглистым (Rosa acicularis), что обеспечивает благоприятные кормовые условия для рыжей полевки. Малина и шиповник являются доминирующими кустарниками в нарушенных ветровалом и пожарами биотопах I и II; в составе кустарниковой растительности частично нарушенного биотопа III и ненарушенного биотопа IV встречается также жимолость обыкновенная (Lonicera xylosteum).
Состояние древостоя в исследуемых биотопах отличается: в нарушенных сообществах он существенно разрежен. Его состав представлен в основном березой пушистой (Betula pubescens) с примесью березы повислой (Betula pendula), а также липой (Tilia cordata). Древесный полог биотопа III менее разреженный по сравнению с биотопами I и II – наряду с березой и липой в составе древостоя присутствуют ель сибирская (Picea obovata) и пихта сибирская (Abies sibirica). Подрост этого биотопа представлен в основном березой пушистой, липой, елью и пихтой. Древостой ненарушенного биотопа IV несомкнутый – в его составе ель, пихта и береза пушистая, а также сосна сибирская (Pinus sibirica). В первом и втором ярусах древесного полога преобладает пихта сибирская.
Известно, что способности рыжей полевки к рытью нор весьма ограничены. Предпочтительными естественными убежищами для этого вида наряду с гнилыми, омшелыми пнями, кучами валежника и вывороченными корнями бурелома являются пустоты под корнями старых живых деревьев, а также прикорневые дупла, где зверьки устраивают несложные жилища [31]. В послепожарных лесных сообществах (биотопы I и II) практически отсутствуют живые взрослые деревья, древостой представлен преимущественно молодыми породами, он существенно разрежен, поэтому его защитная роль для рыжей полевки значительно снижена. Основными укрытиями для грызунов в нарушенных местообитаниях служат пни и лежащие на земле стволы мертвых деревьев (валеж). Кора молодых деревьев может использоваться рыжей полевкой в питании, поэтому древостой в исследуемых нами нарушенных биотопах в большей степени характеризует кормовые условия местообитаний вида. Площадь, занятую живыми, а также сухими деревьями и пнями, оценивали по диаметру ствола, измеряемому у основания на расстоянии не выше 0.5 м от земли. По вычисленным значениям рассчитывали площадь основания каждого ствола и пня в пределах участка, полученные результаты суммировали для оценки общей площади. Площадь покрытия микроучастков стволами упавших мертвых деревьев (валежа) рассчитывали умножением длины и ширины каждого ствола, полученные значения суммировали. В сумме проведено 400 количественных описаний.
Для изучения связи обилия рыжей полевки с характеристиками микросреды (предикторами) использовали множественный регрессионный анализ со стандартизованными частными коэффициентами регрессии β, показывающими, на сколько стандартных отклонений отличается численность вида при изменении средовых переменных на одно стандартное отклонение. Применяли коэффициент детерминации (R2), оценивающий долю объясненной дисперсии. Достоверность уравнений регрессии оценивали по F-критерию. Анализировали распределение остатков модели множественной регрессии, проверяли предикторы на “нормальность”. При отклонении их от нормального распределения проводили процедуру логарифмирования. Попарные сравнения значений обилия рыжей полевки в сравниваемых биотопах проводили непараметрическим тестом Манна-Уитни (Mann-Whitney), достоверность полученных отличий оценивали по Z-критерию.
Биотопическую и хронографическую (межгодовую) изменчивости численности вида и переменных микросреды местообитаний изучали с помощью двухфакторного дисперсионного анализа, где фиксированным фактором являлся “биотоп”, в качестве случайного фактора использовали “год”. В целях сокращения числа микросредовых переменных для характеристики сравниваемых биотопов применяли факторный анализ. Учитывали значения нагрузок по первым двум факторам – F1 и F2, суммарная доля дисперсии которых, вносимая в общую изменчивость микросредовой структуры местообитаний, была наибольшей. Все используемые в работе методы многомерного статистического анализа выполнены в программе “Statistica 6.0” (модули “Основные статистики”, “Множественная регрессия”, “Непараметрическая статистика и распределения”, “ANOVA”, “Факторный анализ”).
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Дисперсионный анализ микросредовых характеристик четырех сравниваемых биотопов, проведенный по результатам количественных описаний в 2013 и 2017 гг., выявил статистически значимые отличия между всеми переменными, за исключением одной, оценивающей площадь покрытия микроучастков валежом. В целом уровень биотопической изменчивости микросреды местообитаний рыжей полевки был выше хронографической (межгодовой) (табл. 2). Согласно данным исследователей, изучавшим микросредовую структуру ненарушенных местообитаний островной популяции рыжей полевки [8], наибольшей вариабельностью в межгодовом масштабе отличаются переменные, характеризующие травянистый, моховый и кустарниковый покровы, а также покрытие микроучастков пнями и валежом. Наименее изменчива переменная, оценивающая покрытие живыми деревьями, промежуточное значение в этом ряду занимает характеристика, связанная с численностью подроста [8].
Таблица 2.
Двухфакторный дисперсионный анализ биотопической и хронографической изменчивости микросредовых характеристик местообитаний рыжей полевки на территории Висимского заповедника
| Обозначение характеристик | Источник изменчивости |
df | Сумма квадратов | Средний квадрат | F-критерий | Уровень значимости, p |
|---|---|---|---|---|---|---|
| MC | Биотоп | 3 | 288.51 | 96.17 | 41.37 | <0.01 |
| Год | 1 | 144.48 | 144.48 | 62.16 | <0.01 | |
| Биотоп × год | 3 | 41.62 | 13.87 | 5.97 | <0.01 | |
| Внутригрупповая | 392 | 911.17 | 2.32 | |||
| Общая | 399 | 1385.78 | ||||
| HC | Биотоп | 3 | 183.06 | 61.02 | 13.83 | <0.01 |
| Год | 1 | 64.88 | 64.88 | 14.71 | <0.01 | |
| Биотоп × год | 3 | 225.17 | 75.06 | 17.01 | <0.01 | |
| Внутригрупповая | 392 | 1729.51 | 4.41 | |||
| Общая | 399 | 2202.62 | ||||
| CS | Биотоп | 3 | 77.46 | 25.82 | 32.30 | <0.01 |
| Год | 1 | 0.03 | 0.03 | 0.04 | ns | |
| Биотоп × год | 3 | 1.86 | 0.62 | 0.77 | ns | |
| Внутригрупповая | 392 | 313.40 | 0.80 | |||
| Общая | 399 | 392.75 | ||||
| TC | Биотоп | 3 | 0.45 | 0.15 | 56.11 | <0.01 |
| Год | 1 | 0.004 | 0.004 | 1.43 | ns | |
| Биотоп × год | 3 | 0.007 | 0.002 | 0.93 | ns | |
| Внутригрупповая | 392 | 1.04 | 0.003 | |||
| Общая | 399 | 1.50 | ||||
| SC | Биотоп | 3 | 0.12 | 0.04 | 17.32 | <0.01 |
| Год | 1 | 0.0009 | 0.0009 | 0.43 | ns | |
| Биотоп × год | 3 | 0.005 | 0.002 | 0.77 | ns | |
| Внутригрупповая | 392 | 0.87 | 0.002 | |||
| Общая | 399 | 0.99 | ||||
| LC | Биотоп | 3 | 1.13 | 0.38 | 1.04 | ns |
| Год | 1 | 0.08 | 0.08 | 0.23 | ns | |
| Биотоп × год | 3 | 3.21 | 1.07 | 2.97 | <0.05 | |
| Внутригрупповая | 392 | 141.03 | 0.36 | |||
| Общая | 399 | 145.45 | ||||
| AU | Биотоп | 3 | 763.09 | 254.36 | 8.80 | <0.01 |
| Год | 1 | 1759.80 | 1759.80 | 60.84 | <0.01 | |
| Биотоп × год | 3 | 2002.53 | 667.51 | 23.08 | <0.01 | |
| Внутригрупповая | 392 | 11339.02 | 28.93 | |||
| Общая | 399 | 15864.44 | ||||
| Обилие | Биотоп | 3 | 31.98 | 10.66 | 21.91 | <0.01 |
| Год | 6 | 49.32 | 8.22 | 16.90 | <0.01 | |
| Биотоп × год | 18 | 18.76 | 1.04 | 2.14 | <0.01 | |
| Внутригрупповая | 1372 | 667.54 | 0.49 | |||
| Общая | 1399 | 767.60 |
При анализе межгодовой изменчивости микросредовых характеристик в нарушенных биотопах (I–III) и ненарушенном местообитании (биотоп IV) нами обнаружено, что наиболее вариабельными оказались переменные, оценивающие покрытие участков мхом (MC) и травянистой растительностью (HC) (табл. 3). Площадь покрытия кустарником (CS) – признак, отражающий в большей степени кормовые условия местообитаний рыжей полевки, в биотопах II–IV не имел межгодовых отличий, статистически значимо он различался по годам в наиболее нарушенном биотопе I. Наименьшей изменчивостью во всех биотопах характеризовались переменные, оценивающие кормозащитные условия микроместообитаний полевок: покрытие стволами живых (TC), сухих деревьев и пнями (SC). Площадь валежа во всех местообитаниях, за исключением биотопа III, также оказалась наименее изменчивой характеристикой. Численность подроста (AU) лишь в ненарушенном биотопе IV имела стабильные межгодовые значения (см. табл. 3).
Таблица 3.
Межгодовая динамика микросредовых характеристик местообитаний рыжей полевки в биотопах (I–IV) Висимского заповедника
| Обозначение характеристик | 2013 г. | 2017 г. | Уровень значимости, р | ||
|---|---|---|---|---|---|
| $\overline X $ ± SE | S | $\overline X $ ± SE | S | ||
| Биотоп I | |||||
| MC | 0.89 ± 0.19 | 1.31 | 0.02 ± 0.01 | 0.08 | *** |
| HC | 5.47 ± 0.32 | 2.27 | 6.68 ± 0.35 | 2.45 | ** |
| CS | 0.21 ± 0.06 | 0.39 | 0.43 ± 0.07 | 0.51 | ** |
| TC | 0.003 ± 0.001 | 0.01 | 0.007 ± 0.001 | 0.01 | ns |
| SC | 0.004 ± 0.002 | 0.02 | 0.01 ± 0.005 | 0.04 | ns |
| LC | 0.77 ± 0.09 | 0.65 | 0.75 ± 0.09 | 0.66 | ns |
| AU | 0.11 ± 0.03 | 0.24 | 1.28 ± 0.13 | 0.90 | *** |
| Биотоп II | |||||
| MC | 2.52 ± 0.31 | 2.22 | 0.02 ± 0.01 | 0.099 | *** |
| HC | 6.49 ± 0.24 | 1.72 | 5.49 ± 0.25 | 1.77 | *** |
| CS | 0.66 ± 0.17 | 1.23 | 0.75 ± 0.14 | 0.99 | ns |
| TC | 0.004 ± 0.001 | 0.009 | 0.005 ± 0.002 | 0.01 | ns |
| SC | 0.007 ± 0.003 | 0.018 | 0.009 ± 0.003 | 0.02 | ns |
| LC | 0.78 ± 0.10 | 0.73 | 0.55 ± 0.06 | 0.45 | ns |
| AU | 0.16 ± 0.04 | 0.29 | 0.49 ± 0.095 | 0.67 | *** |
| Биотоп III | |||||
| MC | 1.26 ± 0.24 | 1.69 | 0.71 ± 0.13 | 0.92 | ** |
| HC | 5.71 ± 0.33 | 2.35 | 3.22 ± 0.27 | 1.92 | *** |
| CS | 0.24 ± 0.06 | 0.43 | 0.13 ± 0.04 | 0.26 | ns |
| TC | 0.08 ± 0.02 | 0.07 | 0.06 ± 0.006 | 0.04 | ns |
| SC | 0.02 ± 0.006 | 0.04 | 0.03 ± 0.006 | 0.05 | ns |
| LC | 0.69 ± 0.07 | 0.47 | 0.94 ± 0.09 | 0.67 | ** |
| AU | 0.49 ± 0.05 | 0.38 | 0.67 ± 0.07 | 0.46 | ** |
| Биотоп IV | |||||
| MC | 3.29 ± 0.31 | 2.17 | 2.17 ± 0.25 | 1.79 | *** |
| HC | 7.15 ± 0.26 | 1.85 | 5.16 ± 0.29 | 2.11 | *** |
| CS | 1.33 ± 0.17 | 1.24 | 1.27 ± 0.19 | 1.32 | ns |
| TC | 0.081 ± 0.01 | 0.084 | 0.07 ± 0.01 | 0.08 | ns |
| SC | 0.05 ± 0.01 | 0.09 | 0.05 ± 0.008 | 0.06 | ns |
| LC | 0.66 ± 0.07 | 0.47 | 0.78 ± 0.09 | 0.62 | ns |
| AU | 0.47 ± 0.08 | 0.54 | 0.47 ± 0.07 | 0.51 | ns |
Дисперсионный анализ изменчивости показателей обилия рыжей полевки выявил высокозначимый уровень биотопических и межгодовых отличий (см. табл. 2). В целом за весь исследуемый период численность населения вида была выше в ненарушенном биотопе, за исключением 2015 г. (рис. 1). Попарное сравнение показателей обилия по критерию Манна-Уитни выявило достоверные различия между наиболее нарушенным и ненарушенным местообитаниями (биотопы I и IV соответственно) за весь исследуемый период. Исключение составили 2013 и 2019 гг., отличающиеся низким уровнем населения рыжей полевки во всех сравниваемых биотопах (табл. 4). Между биотопами I, II и II, III численность вида статистически значимо различалась лишь в 2015 г., а в 2017 г. – в наиболее (биотоп I) и частично (биотоп III) нарушенных местообитаниях (см. табл. 4). Во всех сравниваемых биотопах в 2018 и 2021 гг. численность вида характеризовалась нулевыми значениями, а в биотопе III и в 2015 г. (см. рис. 1). В многолетней популяционной динамике населения рыжей полевки 2017 г. выделяется по характеру распределения численности в отличающихся биотопических условиях: показатели расположены по возрастанию от наиболее нарушенного (биотоп I) к ненарушенному (биотоп IV) (см. рис. 1). Ранее нами при анализе действия средовых факторов на население бурозубок в этом же районе Висимского заповедника было показано [40], что по значениям средней температуры и суммы осадков весенне-летнего сезона 2017 г. характеризовался как относительно благоприятный. Выявленные статистически значимые отличия уровня обилия рыжей полевки в сравниваемых местообитаниях в сходных макросредовых (погодных) условиях этого года, очевидно, могут быть объяснены биотопическими отличиями микросредовой структуры.
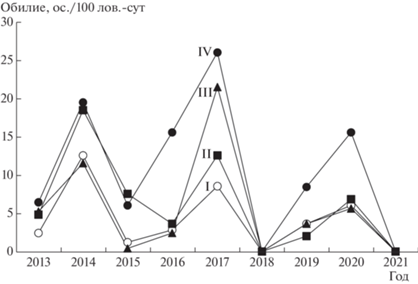
Таблица 4.
Результаты попарных сравнений по тесту Манна-Уитни обилия рыжей полевки в биотопах (I–IV) Висимского заповедника
| Биотоп | Год | ||||||
|---|---|---|---|---|---|---|---|
| 2013 | 2014 | 2015 | 2016 | 2017 | 2019 | 2020 | |
| Z (p) | |||||||
| I, II | –0.73 (ns) | –1.48 (ns) | –1.73* | –0.34 (ns) | –0.50 (ns) | 0.66 (ns) | –0.63 (ns) |
| I, III | –0.41 (ns) | –0.39 (ns) | 0.34 (ns) | 0.17 (ns) | –2.19** | –0.14 (ns) | –0.28 (ns) |
| I, IV | –0.62 (ns) | –2.12** | –1.58* | –3.24*** | –3.16*** | –0.08 (ns) | –2.63*** |
| II, III | 0.28 (ns) | 1.13 (ns) | 2.07** | 0.51 (ns) | –1.59 (ns) | –0.82 (ns) | 0.37 (ns) |
| II, IV | 0.06 (ns) | –0.65 (ns) | 0.09 (ns) | –2.97*** | –2.46** | –1.70* | –2.12** |
| III, IV | –0.20 (ns) | –1.83* | –1.91* | –3.38*** | –0.87 (ns) | –0.96 (ns) | –2.47** |
Факторный анализ структуры микросреды четырех биотопов показал, что наибольший вклад в ее изменчивость вносят два первых фактора (F1 и F2) – суммарно он составил 42.5–54.0% (табл. 5). В биотопах I−III доля объясненной дисперсии по этим двум выделенным факторам принадлежит переменным, оценивающим кормозащитные условия микроместообитаний, а в биотопе IV − переменным, характеризующим преимущественно кормовые условия.
Таблица 5.
Факторные нагрузки переменных микросреды местообитаний рыжей полевки в биотопах (I–IV) Висимского заповедника
| Обозначение характеристик | Год | Биотоп | |||||||
|---|---|---|---|---|---|---|---|---|---|
| I | II | III | IV | ||||||
| F1 | F2 | F1 | F2 | F1 | F2 | F1 | F2 | ||
| MC | 2013 | –0.043 | –0.560 | 0.811 | 0.0001 | 0.860 | 0.054 | –0.205 | 0.666 |
| 2017 | –0.379 | –0.243 | 0.239 | –0.675 | 0.866 | 0.084 | 0.590 | 0.157 | |
| HC | 2013 | –0.711 | 0.198 | 0.077 | –0.891 | –0.191 | –0.803 | –0.655 | –0.078 |
| 2017 | 0.879 | –0.149 | 0.757 | –0.173 | –0.377 | –0.765 | 0.077 | 0.699 | |
| CS | 2013 | 0.199 | –0.546 | 0.207 | 0.536 | 0.533 | –0.493 | –0.795 | –0.078 |
| 2017 | –0.125 | 0.664 | 0.044 | –0.644 | 0.350 | –0.493 | 0.175 | 0.647 | |
| TC | 2013 | 0.479 | 0.382 | –0.393 | 0.288 | –0.383 | 0.225 | 0.456 | –0.436 |
| 2017 | –0.801 | –0.196 | –0.775 | 0.0785 | –0.395 | 0.251 | –0.650 | 0.005 | |
| SC | 2013 | –0.040 | –0.714 | 0.106 | 0.479 | 0.467 | 0.454 | –0.032 | 0.545 |
| 2017 | 0.078 | 0.574 | 0.173 | 0.462 | 0.415 | 0.219 | 0.286 | –0.412 | |
| LC | 2013 | –0.423 | 0.399 | 0.877 | 0.029 | 0.797 | 0.064 | 0.110 | 0.616 |
| 2017 | 0.343 | 0.467 | 0.185 | 0.471 | 0.865 | –0.060 | 0.678 | –0.114 | |
| AU | 2013 | 0.736 | 0.122 | –0.569 | 0.255 | –0.109 | 0.749 | 0.750 | –0.381 |
| 2017 | 0.009 | 0.632 | –0.683 | –0.198 | –0.322 | 0.683 | –0.710 | –0.469 | |
| σ | 2013 | 1.50 | 1.48 | 1.97 | 1.46 | 2.07 | 1.71 | 1.89 | 1.47 |
| 2017 | 1.70 | 1.51 | 1.76 | 1.38 | 2.19 | 1.42 | 1.85 | 1.33 | |
| ∆σ, % | 2013 | 21.41 | 21.18 | 28.07 | 20.87 | 29.60 | 24.45 | 26.95 | 20.96 |
| 2017 | 24.23 | 21.53 | 25.20 | 19.71 | 31.36 | 20.24 | 26.49 | 19.07 | |
Наибольшим микросредовым своеобразием отличался биотоп III, в котором доля объясненной дисперсии переменных по двум первым факторам превышала 50% (54.5 и 51.6% в 2013 и 2017 гг. соответственно), что отличает данное местообитание рыжей полевки от трех других биотопов. Предположительно особенность микросредовой обстановки в биотопе III, в котором число значимых по вкладу переменных оказалось максимальным, можно объяснить более высокой гетерогенностью среды данного местообитания, частично нарушенного ветровалом и пожаром. Как было отмечено выше, в этом биотопе в 2017 г. уровень обилия населения рыжей полевки характеризовался наиболее высокими значениями за весь период наблюдений, как и в биотопе IV (см. рис. 1). Уровень изменчивости микросредовой структуры наиболее нарушенного местообитания (биотоп I) в 2013 г. (через три года после повторного пирогенного нарушения) оказался наименьшим (42.6%) (см. табл. 5), что, возможно, объясняется снижением гетерогенности микросреды, наблюдающимся на ранних стадиях восстановительных сукцессий в дважды пирогенно нарушенном лесном сообществе. Значимым вкладом в микросредовую структуру биотопа I в этот период характеризовались переменные, оценивающие площадь покрытия участков травянистой растительностью и подростом.
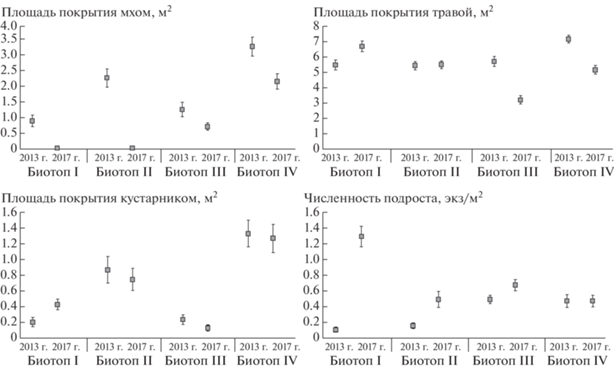
На более поздних стадиях постпирогенного восстановления, в 2017 г., существенный вклад в изменчивость структуры наиболее нарушенного биотопа вносила переменная, характеризующая площадь покрытия основаниями стволов живых деревьев (см. табл. 5). Наряду с этим значимым вкладом отличались характеристики, определяющие защитные условия местообитаний рыжей полевки: площадь покрытия валежом в биотопе II в 2013 г. и в оба года – в биотопе III. Изменчивость микросредовой структуры ненарушенного биотопа IV в 2013 г. существенно определяла переменная, связанная с площадью кустарника, а в 2013 и 2017 гг. – с численностью подроста (см. табл. 5). Площадь кустарниковой растительности в биотопе IV имела более высокие значения по сравнению с нарушенными биотопами I–III, а в биотопе I наиболее высокими показателями численности отличался древесный подрост (рис. 2), что характерно для лесных сообществ с разреженным древостоем. Известно, что подрост характеризует метаболизм экосистемы, он является показателем благополучия леса, его нормального состояния и жизнеспособности. Наличие подроста древесных пород под пологом – главный фактор непрерывного существования леса, так как жизнестойкость растительного сообщества определяется его способностью восстанавливать численность популяций, заменяя новыми погибшие экземпляры. Наиболее обилен подрост в тех участках леса, где древесный ярус разрежен [41].
Анализ связи обилия рыжей полевки с характеристиками микросреды показал, что в нарушенном биотопе II особи вида в наименьшей степени связаны с микросредовой структурой. Статистически значимая связь обнаружена лишь в 2019 г. с переменной, оценивающей площадь покрытия участков основаниями стволов живых деревьев (TC) (табл. 6, 7). В наиболее нарушенном биотопе I грызуны в 2013 г. также предпочитали участки, покрытые живыми деревьями (TC) и валежом (LC), а в 2019 гг. обилие вида было значимо связано с травянистым (HC) и моховым (MC) покрытием микроучастков. В 2016 и 2017 гг. полевки были “привязаны” к участкам, покрытым мхом, а в 2013 г. в биотопе III они предпочитали улучшенные защитные условия среды, которые обеспечивали стоящие сухие деревья и пни (SC), и микроместообитания с более благоприятными кормовыми условиями – наличием кустарниковой растительности (CS). В 2020 г. в этом биотопе обилие рыжей полевки было также связано с кустарником. Таким образом, в биотопах с разной степенью нарушенности рыжая полевка предпочитала участки с улучшенными кормозащитными условиями.
Таблица 6.
Уровни значимости отличий от нуля стандартизированных коэффициентов регрессии (βMC–βAU) обилия рыжей полевки на микросредовые переменные (MC–AU) в биотопах (I–IV) Висимского заповедника
| Биотоп | Коэффициент регрессии | ||||||
|---|---|---|---|---|---|---|---|
| βMC | βHC | βCS | βTC | βSC | βLC | βAU | |
| 2013 г. | |||||||
| I | ns | ns | ns | ** | ns | *** | ns |
| II | ns | ns | ns | ns | ns | ns | ns |
| III | ns | ns | ** | ns | *** | ns | ns |
| IV | ns | ns | *** | ns | ns | ns | ns |
| 2014 г. | |||||||
| I | ns | ns | ns | ns | ns | ns | ns |
| II | ns | ns | ns | ns | ns | ns | ns |
| III | ns | ns | ns | ns | ns | ns | ns |
| IV | ns | ns | ns | ns | ns | ns | ns |
| 2015 г. | |||||||
| I | ns | ns | ns | ns | ns | ns | ns |
| II | ns | ns | ns | ns | ns | ns | ns |
| III | ns | ns | ns | ns | ns | ns | ns |
| IV | ** | ns | ** | ns | ns | ns | ns |
| 2016 г. | |||||||
| I | ** | ns | ns | ns | ns | ns | ns |
| II | ns | ns | ns | ns | ns | ns | ns |
| III | ns | ns | ns | ns | ns | ns | ns |
| IV | ns | ns | ns | ns | ns | ns | ns |
| 2017 г. | |||||||
| I | *** | ns | ns | ns | ns | ns | ** |
| II | ns | ns | ns | ns | ns | ns | ns |
| III | ns | ns | ns | ns | ns | ns | ns |
| IV | ns | ns | ** | ns | ns | ns | ns |
| 2019 г. | |||||||
| I | ** | *** | ns | ns | ns | ns | ns |
| II | ns | ns | ns | *** | ns | ns | ns |
| III | ns | ns | ns | ns | ns | ns | ns |
| IV | ns | ns | ns | ns | ns | ns | ns |
| 2020 г. | |||||||
| I | ns | ns | ns | ns | ns | ns | ns |
| II | ns | ns | ns | ns | ns | ns | ns |
| III | ns | ns | *** | ns | ns | ns | ns |
| IV | ns | ns | ns | ns | ns | ns | ns |
Таблица 7.
Результаты множественного регрессионного анализа связи обилия рыжей полевки (зависимой переменной) с микросредовыми характеристиками (предикторами) биотопов I–IV
| Год | Уровень обилия, ос/100 лов.-сут | Уравнение регрессии | R2 | F | p |
|---|---|---|---|---|---|
| Биотоп I | |||||
| 2013 | 2.4 | Y = –0.047 + 8.090XTC + 0.177XLC | 0.463 | 6.43 | <0.01 |
| 2016 | 2.8 | Y = 0.114 + 1.42XMC | 0.290 | 4.40 | <0.05 |
| 2017 | 8.5 | Y = 0.793 – 0.101XHC + 0.172XLC | 0.494 | 7.58 | <0.01 |
| 2019 | 3.2 | Y = 0.656 + 1.928XMC – 0.069XHC | 0.514 | 5.49 | <0.05 |
| Биотоп II | |||||
| 2019 | 2.0 | Y = 0.596 + 38.93XTC | 0.369 | 7.59 | <0.01 |
| Биотоп III | |||||
| 2013 | 5.2 | Y = 0.042 + 6.632XSC + 0.448XCS | 0.485 | 7.25 | <0.01 |
| 2020 | 5.6 | Y = 0.137 + 1.08XCS | 0.522 | 17.97 | <0.01 |
| Биотоп IV | |||||
| 2013 | 6.4 | Y = –0.021 + 0.256XCS | 0.399 | 9.09 | <0.01 |
| 2015 | 6.0 | Y = 0.369 + 0.137XSC – 0.076XMC | 0.382 | 4.02 | <0.05 |
| 2017 | 26.0 | Y = 0.670 + 0.290XCS | 0.353 | 6.83 | <0.05 |
В ненарушенном местообитании численность вида оказалась тесно связанной с переменными микросреды, оценивающими преимущественно кормовые условия. В 2013, 2015 и 2017 гг. полевки предпочитали микроучастки с кустарниковой растительностью (см. табл. 6, 7). На примере островной популяции рыжей полевки было показано [8], что выбор характеристик микросреды этим видом отличается на разных фазах его популяционной динамики. Мы также выявили тесную связь вида с микросредовым окружением на разных фазах динамики численности, но лишь в условиях ненарушенной среды биотопа IV. Оказалось, что в этом местообитании рыжая полевка предпочитает участки с преобладанием кустарника как на фазе низкого уровня обилия населения в 2013 и 2015 гг., так и на пике его численности в 2017 г. (см. табл. 6, 7). Как было отмечено выше, данная микросредовая характеристика отличается низкой межгодовой изменчивостью в биотопах II–IV (см. табл. 3). Однако только в ненарушенном местообитании обнаружена связь обилия рыжей полевки с данным микросредовым параметром при разном уровне ее населения (см. табл. 7). Покрытие участков мхом – характеристика среды, не являющаяся предпочтительной для рыжей полевки, но, вероятно, в комплексе с другими микросредовыми показателями определяющая благоприятные условия для обитания вида. Данная характеристика отличалась высокими значениями в ненарушенном биотопе наряду с переменной микросреды, оценивающей площадь покрытия участков кустарниковой растительностью (см. рис. 2).
ЗАКЛЮЧЕНИЕ
Результаты проведенного исследования позволяют констатировать, что в ряду факторов, влияющих на “успешность существования” рыжей полевки в разных биотопических условиях, в первую очередь таких, как уровень численности населения вида и микросредовая структура его местообитаний, последний играет важную роль. Анализ уровня значений обилия – важной информативной характеристики, отражающей степень “экологического благополучия” вида в природной среде, показал, что наиболее благоприятные условия для успешного существования рыжей полевки на исследуемой охраняемой территории Среднего Урала складываются в ненарушенном внешними неблагоприятными воздействиями биотопе, где вид преобладает по численности.
В условиях биотопической неоднородности, вызванной последствиями природных катастрофических явлений, обилие рыжей полевки детерминировано разными микросредовыми параметрами. В нарушенных местообитаниях особи вида в большей степени привязаны к участкам с улучшенными защитными условиями, а в ненарушенной биотопической среде наблюдается “отклик” численности рыжей полевки на микросредовые переменные, характеризующие кормовые условия местообитаний. Уровень значений показателей, оценивающих пищевые ресурсы вида в отличающихся биотопах – покрытие участков травянистой и кустарниковой растительностью, в целом выше в ненарушенных местообитаниях, что, несомненно, влияет на выбор рыжей полевкой среды местообитаний.
Таким образом, на основе полученных результатов можно заключить, что наиболее предпочтительными для рыжей полевки на территории Висимского заповедника являются условия среды ненарушенного биотопа. Этот вывод обусловлен высоким уровнем обилия и его тесной связью при разном уровне численности населения с микросредовыми параметрами, оценивающими преимущественно кормовые условия местообитаний вида.
Выявленные особенности выбора среды рыжей полевкой, предпочитающей условия ненарушенного биотопа с сохранившимся древостоем условиям более осветленных местообитаний в пирогенно нарушенных разреженных лесных сообществах, не вполне согласуются с классическими литературными данными [30]. Полученные нами результаты лишь подтверждают предположение о возможном проявлении в нестабильных условиях среды неизвестных ранее в норме экологических, в том числе пищевых и поведенческих, реакций фоновых видов на изменение условий обитания [20].
Работа выполнена в рамках государственного задания Института экологии растений и животных УрО РАН № 122021000091-2.
Автор выражает искреннюю благодарность старшим научным сотрудникам Висимского государственного природного биосферного заповедника Р.З. Сибгатуллину и Н.В. Беляевой за постоянную поддержку в работе и помощь в геоботанических описаниях, а также И.Ф. Вурдовой и В.Д. Араповой за помощь в проведении полевых исследований.
Список литературы
Hodara K., Busch M. Patterns of macro and microhabitat use of two rodent species in relation to agricultural practices // Ecol. Res. 2010. V. 25. № 1. P. 113–121.
Наумов Н.П. Структура популяций и динамика численности наземных позвоночных млекопитающих // Зоол. журн. 1967. Т. 46. Вып. 10. С. 1470–1486.
Шварц С.С. Популяционная структура вида // Зоол. журн. 1967. Т. 46. Вып. 10. С. 1456–1469.
Torre I., Arrizabalaga A. Habitat preferences of the bank vole Myodes glareolus in a Mediterranean mountain range // Acta Theriol. 2008. V. 53. P. 241–250.
Андреева Т.А., Окулова Н.М. Экологические предпочтения лесных полевок // Экология. 2009. № 2. С. 149–154.
Mohammadi S. Microhabitat selection by small mammals // Advances in Biological Research. 2010. V. 4. № 5. P. 283–287.
Villanueva-Hernández A.I., Delgado-Zamora D.A., Heynes-Silerio S.A. et al. Habitat selection by rodents at the transition between the Sierra Madre Occidental and the Mexican Plateau, México // J. Mammal. 2017. V. 98. № 1. P. 293–301.
Буяльска Г., Лукьянов О.А., Мешковска Д. Детерминанты локального пространственного распределения численности островной популяции рыжей полевки // Экология. 1995. № 1. С. 35–45.
Rosenzweig M.L. A theory of habitat selection // Ecology. 1981. V. 62. № 2. P. 327–335.
Morris D.W. Test of density-dependent habitat selection in a patchy environment // Ecol. Monogr. 1987. V. 57. № 4. P. 269–281.
Falkenberg J.C., Clarke J.A. Microhabitat use of deer mice: effects of interspecific interaction risks // J. Mammal. 1998. V. 79. № 2. P. 558–568.
Drickamer L.C. Microhabitat preferences of two species of deer mice Peromyscus in a northeastern United States deciduous hard wood forest // Acta Theriol. 1990. V. 35. P. 241–252.
Johannesen E., Mauritzen M. Habitat selection of grey-sided voles and bank voles in two subalpine populations in southern Norway // Ann. Zool. Fenn. 1999. V. 36. P. 215–222.
Johannesen E., Brudevoll J., Jenstadt M. et al. Behavioural dominance of grey-sided voles over bank voles in dyadic encounters // Ann. Zool. Fenn. 2002. V. 39. P. 43–47.
Trebatická L., Sundell J., Tkadlec E. et al. Behavior and resource use of two competing vole species under shared predation risk // Oecologia. 2008. V. 157. P. 707–715.
Morris G.J., Hostetler A., Oli M.K. et al. Effects of predation, fire, and supplemental feeding on populations of two species of Peromyscus mice // J. Mammal. 2011. V. 92. P. 934–944.
Letnic M., Tischler M., Gordon C. Desert small mammal responses to wildfire and predation in the aftermath of a La Niná driven resource pulse // Austral. Ecology. 2013. V. 38. P. 841–849.
Novillo A., Cuevas M.F., Ojeda A.A. et al. Habitat selection and coexistence in small mammals of the southern Andean foothills (Argentina) // Mamm. Res. 2017. V. 62. P. 219–227.
Magnusson M., Samelius G., Hörnfeldt B. et al. Diet shift in bank voles induced by competition from grey-sided voles? // Integrative Zoology. 2019. V. 14. P. 376–382.
Шилова С.А., Неронов В.В., Касаткин М.В. и др. Пожары на современном этапе развития полупустыни юга России: влияние на растительность и население грызунов // Успехи совр. биол. 2007. Т. 127. № 4. С. 372–386.
Кулешова Л.В. Экологические и зоогеографические аспекты воздействия пожаров на лесных птиц и млекопитающих // Зоол. журн. 1981. Т. 60. Вып. 10. С. 1542–1552.
Dueser R.D., Porter J.H. Habitat use by insular small mammals relative effects of competition and habitat structure // Ecology. 1986. V. 67. № 1. P. 195–201.
Hengriques R.P.B., Bizerril M.X.A., Palma A.R.T. Changes in small mammal populations after fire in a patch of unburned cerrado in Central Brazil // Mammalia. 2000. V. 64. № 2. P. 173–185.
Истомин А.В. Влияние ветровалов на динамику сообществ мелких млекопитающих в естественных лесах южной тайги // Вестн. МГУ. Лесной вестн. 2009. № 1. С. 196–201.
Lee E.J., Rhim S.G., Son S.H. et al. Differences in small-mammal and stand structures between unburned and burned pine stands subjected to two different post-fire silvicultural management practices // Ann. Zool. Fenn. 2012. V. 49. № 3. P. 129–138.
Zwolak R., Pearson D.E., Ortega Y.K. et al. Mechanisms driving postfire abundance of a generalist mammal // Can. J. Zool. 2012. V. 90. P. 51–60.
Fordyce A., Hradsky B.A., Ritchie E.G. et al. Fire affects microhabitat selection, movement patterns, and body condition of an Australian rodent (Rattus fuscipes) // J. Mammal. 2016. V. 97. № 1. P. 102–111.
Садыков О.Ф., Большаков В.Н., Баженов А.В. Пространственная структура популяций лесных полевок // Экология. 1984. № 4. С. 58–64.
Шилов И.А. Эколого-физиологические основы популяционных отношений у животных. М.: Изд-во МГУ, 1977. 262 с.
Громов И.М., Ербаева М.А. Млекопитающие фауны России и сопредельных территорий. Зайцеобразные и грызуны. СПб.: ЗИН, 1995. 522 с.
Европейская рыжая полевка / А.А. Аристов и др. М.: Наука, 1981. 352 с.
Ивантер Э.В. Очерки популяционной экологии мелких млекопитающих на северной периферии ареала. М.: Тов-во научн. изд. КМК, 2018. 770 с.
Geuse P. Spatial microhabitat of bank voles and wood mice in a forest in central Belgium // Acta Zool. Fenn. 1985. V. 173. P. 61–64.
Mazurkiewicz M. The influence of undergrowth distribution on utilization of space by bank vole populations // Acta Theriol. 1986. V. 31. P. 55–69.
Mazurkiewicz M. Population dynamics and demography of the bank vole in different tree stands // Acta Theriol. 1991. V. 36. P. 207–227.
Mazurkiewicz M. Factors influencing the distribution of the bank vole in forest habitats // Acta Theriol. 1994. V. 39. P. 113–126.
Miklos P., Ziak D. Microhabitat selection by three small mammal species in oak-elm forest // Folia Zool. 2002. V. 51. № 4. P. 275–288.
Лукьянова Л.Е., Бобрецов А.В. Выбор рыжей полевкой (Clethrionomys glareolus Schreber, 1780) микроместообитаний в стабильных и дестабилизированных условиях среды // Вестн. Томского гос. ун-та. Биология. 2014. № 4 (28). С. 88–107.
Большаков В.Н., Бердюгин К.И., Васильева И.А., Кузнецова И.А. Млекопитающие Свердловской области: справочник-определитель. Екатеринбург, 2000. 240 с.
Лукьянова Л.Е., Ухова Н.Л., Ухова О.В., Городилова Ю.В. Население обыкновенной бурозубки (Sorex araneus, Eulipotyphla) и кормообеспеченность ее местообитаний в экологически контрастной среде // Экология. 2021. № 4. С. 298–311.
Чертовской В.Г. Таежное лесоводство. М.: Лесная пром-сть, 1974. 232 с.
Дополнительные материалы отсутствуют.