

Экология, 2023, № 1, стр. 3-12
Структура комплекса опылителей Veronica spicata L. на северной границе распространения
Н. И. Филиппов a, *, Л. В. Тетерюк a
a Институт биологии ФИЦ Коми научного центра УрО РАН
167982 Сыктывкар, ул. Коммунистическая, 28, Россия
* E-mail: filippov@ib.komisc.ru
Поступила в редакцию 06.05.2022
После доработки 05.08.2022
Принята к публикации 23.08.2022
- EDN: GZGYHF
- DOI: 10.31857/S0367059723010055
Аннотация
На Севере лимитирующим фактором для энтомофильных растений становится низкая численность насекомых-опылителей, которая усугубляется коротким вегетационным периодом и часто неблагоприятными погодными условиями для их жизнедеятельности. В работе представлены результаты изучения комплекса опылителей специализированного мелиттофильного вида Veronica spicata на северном пределе его распространения. Показано, что структура этого комплекса определяется особенностями строения цветка V. spicata, широтным положением и дифференциацией экологических ниш вида на Севере. В отличие от других частей ареала, где важную роль в опылении V. spicata играют одиночные пчелы, на Севере эту функцию выполняют представители рода Bombus Latr. (Hymenoptera, Apidae). С учетом уровня видового разнообразия и структуры населения основу комплекса опылителей V. spicata составляют короткохоботковые виды шмелей: Bombus lucorum s.l., B. jonellus, B. pratorum, B. sporadicus, а также среднехоботковый B. pascuorum. В соответствии с дифференциацией экологических ниш V. spicata на Севере выявлены различия в структуре комплекса опылителей и стратегиях фуражировки шмелей.
На Севере проблема перекрестного опыления растений связана с меньшей численностью насекомых-опылителей. Отсутствие медоносной пчелы (Apis mellifera) и низкая численность иных пчел являются следствием климатических особенностей северных территорий (части таежной, лесотундровой и тундровой зон), что приводит к перестроению комплекса насекомых-опылителей. Роль основных опылителей энтомофильных растений здесь переходит к представителям Diptera и Bombus Latr. (Hymenoptera, Apidae) [1]. В сравнении с другими Apidae Bombus более активны при низких температурах и интенсивности света, дождливой погоде, живут дольше [2, 3]. В таежной зоне Евразии эта мезофильная умеренно-теплолюбивая группа насекомых составляет 55–70%, а в тундровых биогеоценозах – 85–95% от общего числа пчелиных [1].
В последние десятилетия проблема усугубляется резким сокращением общей численности медоносных пчел и шмелей, которое приобрело глобальный характер [4, 5]. “Кризис опылителей” будет иметь негативные последствия для энтомофильных растений (критичные – для облигатных ксеногамов) [6, 7], приведет к изменениям в составе и структуре растительных сообществ. Особого внимания в этом плане требует группа редких и исчезающих растений [8–10]. Недостаток сведений о системах скрещивания и опылителях мешает эффективному сохранению энтомофильных видов, разработке видовых программ реставрации и поддержания численности популяций.
Объектом наших исследований является энтомофильный вид специализированного мелиттофильного комплекса Veronica spicata L. (сем. Plantaginaceae). Северная граница распространения этого лесостепного евразиатского вида в европейской части России проходит по линии Финский залив – устье р. Вычегда – нижнее течение р. Печора (около 66° с.ш.), по территории Республики Коми и сопредельных регионов. В наиболее северных местонахождениях V. spicata входит в состав скального реликтового комплекса [11]. Вид на северном пределе распространения редок, включен в Красные книги Республики Коми [12], Ханты-Мансийского автономного округа – Югры [13], Ненецкого автономного округа [14].
Самоподдержание популяций V. spicata осуществляется за счет вегетативного и семенного размножения. В работе G.B. Wilson [15] приводятся данные о способности V. spicata к самоопылению, R. Scalone с соавт. [16] относят его к факультативным ксеногамам, Е.И. Демьянова и А.Н. Пономарев на Среднем Урале – к облигатным ксеногамам [17]. В опылении вида участвуют представители семейств Andrenidae, Apidae, Colletidae, Halictidae, Melittidae, среди которых наибольшим числом видов представлены одиночные пчелы [18–21]. На Севере вопросы консорционных связей V. spicata с опылителями остаются малоизученными.
Цель настоящей работы – изучение структуры комплекса опылителей V. spicata в двух основных типах местообитаний на северной границе распространения вида.
МАТЕРИАЛ И МЕТОДЫ
Исследования проведены в 2005–2018 гг. в пределах Вычегодско-Мезенской равнины и Тиманского кряжа, в подзонах средней и северной тайги. Материал собран на охраняемых природных территориях Республики Коми и прилегающих участках: на боровых террасах рек Сысола (заказник “Ужгинский”) и Вычегда (“Немский”), а также на выходах известняков Среднего (“Белая Кедва”) и Южного Тимана (“Сойвинский”) (рис. 1).
Рис. 1.
Местонахождения Veronica spicata на территории Республики Коми: 1 – боровые террасы, 2 – выходы кальцийсодержащих пород в долинах рек, 3 – места отбора проб.
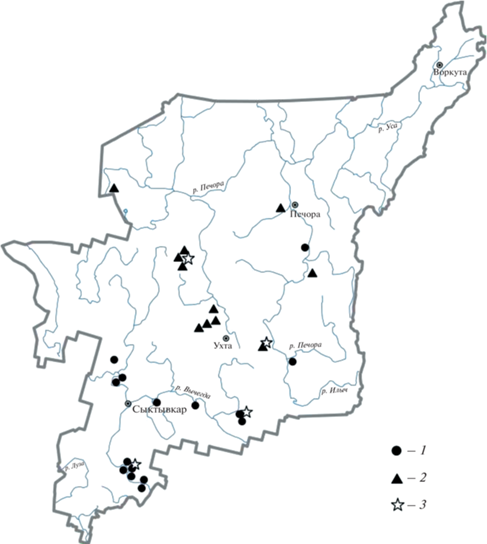
На 18 пробных площадях выполнены геоботанические описания (в сосняках размером 20 × 20 м, на выходах известняков – в естественных границах сообществ), на 8 участках (по 4 на тип экотопа) проведены популяционные исследования V. spicata [22, 23]. Названия растений приведены в соответствии с базой данных “World Flora Online”. При географическом анализе использован метод биогеографических координат [24–26].
Насекомых собирали в светлое время суток на цветущих растениях при помощи энтомологического сачка методом безвыборочного вылова, при фиксации факта питания. Выборка составила 1500 экз. насекомых. Таксоны рода Bombus Latr. приведены по P.H. Williams [27]. Для оценки видового разнообразия и меры доминирования использованы индексы Маргалефа, Шенона, Симпсона и Бергера-Паркера, коэффициент Съеренсена-Чекановского [28], для выявления детерминирующих видов – метод главных компонент (PCA). Анализ данных проведен с использованием программных пакетов ExceltoR [29] и R версии 4.1.3 (пакет “bipartite”), обработка геоботанических описаний – методом MNS-ординации с использованием фитоиндикационных экологических шкал [30, 31].
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Дифференциация экологических ниш V. spicata на Севере. На территории Республики Коми известно более 30 местонахождений V. spicata, которые связаны с интра- или азональным типами растительности. На Вычегодско-Мезенской равнине вид встречается в сосняках лишайниковых на боровых террасах (Pinetum cladinosum), севернее – в местах выхода кальцийсодержащих пород по рекам Тимана, Малокожвинской возвышенности, Предуралья (см. рис. 1).
Методами фитоиндикации и MNS-ординации показано разделение обследованных биотопов на две группы (рис. 2). Значимый (p < 0.01) вклад в дифференциацию вносит ряд климатических факторов. На боровых террасах в биотопы поступает больше тепла, менее выражен диапазон перепадов между летними и зимними температурами, зимние температуры выше, чем на скальных выходах (табл. 1, 2). Это наиболее теплообеспеченные выделы на территории Республики Коми [32], для которых характерны более сухие и кислые почвы, а также высокий уровень освещенности. В сообществах Pinetum cladinosum для травяно-кустарничкового яруса характерны низкие показатели видового разнообразия. Вместе с V. spicata встречаются 29 (от 8 до 18) таксонов высших споровых и цветковых растений. Эту группу биотопов дифференцирует ряд представителей бореальной, южнобореальной и лесостепной широтных групп, в том числе Anthoxanthum odoratum L., Calluna vulgaris (L.) Hill., Carex ericetorum Pollich, Pilosella officinarum Vaill., Anemone patens L.
Рис. 2.
Дифференциация двух типов местообитаний Veronica spicata на территории Республики Коми. Корреляция экологических факторов с осями ординации: по оси 1 – Tm, Kn, Cr, Hd, Rc, Lc (p < 0.01), по оси 2 – Om, Tr, fH (p < < 0.01), Rc (p < 0.05). Условные обозначения. Фитоиндикационных экологических шкал (здесь и далее): Tm – термоклиматическая, Kn – континентальности климата, Om – омброклиматическая аридности-гумидности, Cr – криоклиматическая, Lc – освещенности-затенения, Hd – увлажнения почв, Tr – солевой режим почв, Nt – богатства почв азотом, Rc – кислотность почв, fH – переменности увлажнения.

Таблица 1.
Экологический диапазон Veronica spicata на территории Республики Коми
| Диапазон | Экологические шкалы | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Tm | Kn | Om | Cr | Lc | Hd | Tr | Nt | Rc | fH | |
| Потенциальный | 4.0–12.0 | 3.0–13.0 | 5.0–11.0 | 3.0–11.0 | 1.0–5.0 | 3.0–13.0 | 3.0–11.0 | 1.0–5.0 | 3.0–11.0 | 5.0–8.0 |
| Фактический, на боровых террасах | 7.6–7.9 | 8.4–8.8 | 8.1–8.5 | 6.6–7.0 | 3.1–3.5 | 11.0–12.0 | 5.0–5.9 | 4.1–4.5 | 5.4–6.7 | 5.2–5.7 |
| Фактический, на скальных выходах | 6.9–7.6 | 8.5–9.5 | 7.9–8.7 | 6.0–7.2 | 3.3–4.1 | 11.4–12.8 | 4.9–6.0 | 4.3–4.8 | 6.3–7.4 | 5.1–6.7 |
Таблица 2.
Корреляция осей координат MNS со значениями экологических факторов
| Оси | Экологические шкалы | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Tm | Kn | Om | Cr | Lc | Hd | Tr | Nt | Rc | fH | |
| 1 | –0.71* | 0.76* | 0.22 | –0.63* | 0.75* | 0.75* | –0.15 | 0.46 | 0.62* | 0.20 |
| 2 | 0.12 | 0.01 | 0.79* | –0.31 | 0.08 | 0.24 | –0.59* | –0.19 | –0.53** | –0.60* |
В районе выходов кальцийсодержащих пород вдоль рек V. spicata встречается на открытых или облесенных склонах преимущественно южной экспозиции, в составе реликтового скального флористического комплекса. Вместе с V. spicata встречаются 78 (в отдельных сообществах – от 13 до 40) таксонов. Это виды, хорошо приспособленные к недостатку тепла, сезонным перепадам температур, холодным зимам, в том числе представители арктической, аркто-альпийской и гипоарктической широтных групп (Arctous alpina (L.) Nied., Euphrasia frigida Pugsley, Minuartia verna (L.) Hiern., Poa alpina L., Salix recurvigemmis A.K.Skvortsov, S. reticulata L., Vaccinium uliginosum L.), а также виды с азиатским или сибирским распространением (Adonis sibirica (Patrin ex DC.) Ledeb., Chrysanthemum zawadskii Herbich, Saussurea parviflora (Poir.) DC, Silene chamarensis Turcz. и др.). Почвенные условия (см. табл. 1) более благоприятны для растений по плодородию, кислотности и увлажнению [33], чем в первом типе экотопов. Приуроченность V. spicata к кальцийсодержащим породам на северном пределе распространения отмечена и на других территориях [34].
Показатели занимаемой площади, численности и доли цветущих растений (табл. 3, рис. 3) свидетельствуют о более благоприятных условиях для развития V. spicata на боровых террасах, чем на выходах известняков.
Таблица 3.
Характеристика экотопических популяций Veronica spicata
| Критерии | Боровые террасы | Выходы известняков | ||
|---|---|---|---|---|
| Ужгинский | Немский | Белая Кедва | Сойвинский | |
| Площадь, км2 | 7 | 17 | 0.01 | 0.5 |
| Численность, шт. | До 1000 | До 10 000 | 500 | 1000 |
| Типы обследованных ценопопуляций | Переходная, молодая | Переходная, молодая | Молодые | Молодые |
| Плотность, шт/м2 | 1.6–8.9 | 14.6–76 | 3.9–5.7 | 5.6–9.1 |
Рис. 3.
Усредненные онтогенетические спектры экотопических популяций V. spicata на выходах известняков (1) и боровых террасах (2): j – ювенильная, im – имматурная, v – виргинильная, g – генеративная, ss – субсенильная, s – сенильная, sc – отмершие.

Комплекс опылителей V. spicata включает 19 видов насекомых из 3 семейств отрядов Hymenoptera и Diptera (табл. 4). Среди опылителей из семейства Apidae преобладают виды с широкими ареалами, преимущественно палеарктические, голарктические по долготной составляющей и температные, полизональные – по широтной. Как правило, такие таксоны обладают большей экологической пластичностью, осваивают разные биотопы, что особо актуально на Севере с его нестабильными климатическими условиями в летний период. Опылители V. spicata относятся к луговой, лугово-болотной, лугово-лесной и эвритопной топическим группам (или “группировкам” в понимании А.Г. Татаринова и М.М. Долгина [35]), преобладают представители двух последних групп. Вместе с тем в этом комплексе отмечены редкие виды: B. muscorum, занесенный в основной список Красной книги Республики Коми [12], и виды дополнительного списка (бионадзор) – B. schrencki, B. sporadicus.
Таблица 4.
Распространение, показатели численности, видового состава, характеристика ротового аппарата насекомых-опылителей Veronica spicata
| Виды насекомых | Длина хоботка | Тип ареала | Боровые террасы | Выходы известняков | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Немский | Ужгинский | Белая Кедва | Сойвинский | |||||||
| доля, % | балл обилия |
доля, % | балл обилия |
доля, % | балл обилия |
доля, % | балл обилия |
|||
| Семейство Syrphidae | ||||||||||
| Melangyna barbifrons* | – | ТЕБ | – | – | – | – | – | – | 1.2 | 1 |
| Eristalis interrupta* | – | ЦАТ | – | – | – | – | 2.6 | 1 | – | – |
| Семейство Halictidae | ||||||||||
| Dufourea minuta* | – | ЕСТ | – | – | – | – | 2.6 | 1 | – | – |
| Sphecodes sp.* | – | – | – | – | – | – | 1.3 | 1 | – | – |
| Семейство Apidae | ||||||||||
| Bombus balteatus Dahl. | Д | ГААБ | – | – | – | – | – | 2 | – | – |
| B. bohemicus Seidl | К | ТПП | – | – | – | – | – | – | – | 1 |
| B. cingulatus Wahl. | С | ТЕТ | – | – | – | – | – | – | – | 3 |
| B. consobrinus Dahl. | Д | ТПП | – | – | – | – | – | 3 | – | 3 |
| B. distinguendus Mor. | С | ТЕБ | – | – | – | – | – | 3 | – | 3 |
| B. flavidus Evers.* | К | ГААБ | 4 | 2 | 10.7 | 3 | – | – | 6.2 | 2 |
| B. hortorum (L.)* | Д | ЕСТ | – | – | 5.3 | 2 | 2.6 | 1 | 3.7 | 1 |
| B. hypnorum (L.)* | К | ПОП | 9.9 | 3 | 12 | 3 | 6.6 | 2 | 6.2 | 2 |
| B. jonellus Kirby* | К | ГААБ | 14.8 | 4 | – | – | 14.5 | 4 | 7.4 | 2 |
| B. lucorum s.l.* | К | ГАОП | 25.7 | 5 | – | – | – | – | – | – |
| B. muscorum (L.)* | С | ТЕТ | 2 | 1 | – | – | – | – | – | – |
| B. norvegicus (Sp.-Schn.)* | К | ПОП | – | – | 8 | 3 | 9.2 | 3 | 7.4 | 3 |
| B. pascuorum (Scop.)* | С | ТЕТ | 18.8 | 4 | 10.7 | 3 | 9.2 | 3 | 8.6 | 3 |
| B. patagiatus Nyl.* | К | ПОП | – | – | 2.6 | 1 | – | – | – | – |
| B. pratorum (L.)* | К | ЕСТ | 4 | 2 | 6.7 | 2 | 10.5 | 3 | 13.6 | 3 |
| B. schrencki Mor.* | С | ПОП | 4.9 | 3 | 8 | 3 | 6.6 | 2 | 7.4 | 2 |
| B. semenoviellus Skor.* | К | ЕСТ | – | – | – | – | – | – | 2.5 | 1 |
| B. soroeensis (F.) | К | ТЕП | – | – | – | – | – | 3 | – | – |
| B. sporadicus Nyl.* | К | ПОП | 2 | 4 | 22.7 | 5 | 22.4 | 3 | 21 | 4 |
| B. sylvestris (Lep.)* | К | ТПП | 3 | 1 | 5.3 | 2 | 6.6 | 3 | 8.6 | 3 |
| B. veteranus (F.)* | С | ТЕТ | 10.9 | 3 | 8 | 3 | 5.3 | 2 | 6.2 | 2 |
| Общее число видов | 19 | 11 | 11 | 11 (17) | 13 (17) | |||||
| Индекс Шенона | 2.42 | 2.47 | 2.71 | 2.74 | ||||||
| Индекс Маргалефа | 2.11 | 2.32 | 3.1 | 3.41 | ||||||
| Индекс Симпсона | 1.01 | 1.03 | 1.03 | 0.99 | ||||||
| Индекс Бергера-Паркера | 0.23 | 0.23 | 0.12 | 0.14 | ||||||
Примечание: К – короткохоботковые виды, С – среднехоботковые, Д – длиннохоботковые; ЕСТ – евро-сибирская температная; ТЕБ – трансевразийская бореальная; ТЕП – трансевразийская полизональная; ТЕТ – трансевразийская температная; ПОП – палеаркто-ориентальная полизональная; ТПП – транспалеарктическая полизональная; ГААБ – голарктическая арктобореальная; ГАОП – голаркто-ориентальная полизональная; ЦАТ – циркумголарктическая температная; * – отмечены опылители; в скобках указано общее число видов шмелей на скальных обнажениях.
В отличие от других частей ареала V. spicata, где важную роль в опылении играют одиночные пчелы [18, 19], на северной границе распространения вида эту функцию осуществляют шмели (Bombus Latr.) (см. табл. 4). Их доля доходит до 100% от общего числа видов опылителей в сосновых борах и до 98.8% на скальных обнажениях Тимана, где 1.2% относятся к семействам Halictidae и Syrphidae. Это соответствует общим географическим закономерностям трансформации комплекса опылителей на Севере [1].
Структура комплекса опылителей V. spicata тесно связана с морфологией ее цветков, в частности с длиной трубки венчика, которая составляет около 2 мм. Как отмечал C. M. Kampny [36], короткие трубчатые цветки рода Veronica эволюционно приспособлены для опыления пчелами и мухами с коротким хоботком. В районе исследований среди шмелей-опылителей V. spicata 10 таксонов (67%) приходится на долю короткохоботковых видов, 4 (27%) – среднехоботковых. Несмотря на присутствие в мелиттофауне изученных участков нескольких длиннохоботковых шмелей, среди опылителей V. spicata отмечен лишь один вид (B. hortorum): число его особей составляет 2.6–5.3% от общей численности (см. табл. 4).
Влияние экотопа. Показано, что в соответствии с дифференциацией экологических ниш V. spicata состав насекомых-опылителей подразделяется на две группы: скальные обнажения (“Сойвинский”, “Белая Кедва”) и боровые террасы (“Ужгинский”, “Немский”). Уровень сходства внутри кластеров колеблется в пределах 70–75%, между ними – 64% (p < 0.05).
Высокий уровень видового разнообразия шмелей на скальных выходах (см. табл. 4) обусловлен широким спектром экологических условий и высоким ценотическим разнообразием. Из обследованных локалитетов только в этом типе экотопов выявлены семь таксонов, не являющихся опылителями V. spicata. Среди них B. balteatus, местонахождения которого на Среднем Тимане – наиболее южные в регионе, B. semenoviellus – наиболее западные в ареале, B. bohemicus (клептопаразит B. distinguendus и других таксонов) и B. consobrinus, распространение которого сопряжено с Aconitum septentrionale [37].
В скальных экотопах одновременно с V. spicata шмели фуражируют на 16 таксонах реликтового флористического комплекса (см. рис. 4а), включая специализированные мелиттофильные виды Astragalus frigidus (L.) A. Gray, Hedysarum hedysaroides subsp. arcticum (B.Fedtsch.) P.W. Ball, Thymus sp., Vicia sepium L., Vaccinium vitis-idaea L. Среди опылителей V. spicata отмечено 57–75% видов локальных фаун шмелей (см. рис. 4б). Учитывая небольшие размеры и численность популяций (см. табл. 3, рис. 3), значительное давление в конкуренции за опылителей может приводить к попаданию недостаточного количества пыльцы [38, 39] на цветки V. spicata.
Рис. 4.
Взаимосвязь шмелей и опыляемых ими растений на скальных обнажениях (а) и боровых террасах (б).

В обследованных локалитетах на боровых террасах (“Ужгинский”, “Немский”) выявлен также достаточно высокий уровень видового разнообразия шмелей. Мы связываем его с обилием кормовых ресурсов, предоставляемых опылителям цветками V. spicata. Образуя крупные популяции (см. табл. 3, рис. 3), в период массового цветения этот вид становится доминирующим источником нектара и пыльцы для 100% представителей рода Bombus Latr., выявленных в локальных фаунах боровых террас. Такая олиголектичность шмелей (см. рис. 4б) обусловлена характерными для Pinetum cladinosum низкими показателями видового разнообразия и обилия других растений.
Доминантный комплекс опылителей V. spicata на Севере образуют B. jonellus, B. lucorum s.l., B. pascuorum, B. sporadicus. Эти же таксоны являются детерминирующими при ординации выборок опылителей V. spicata с учетом количественных данных (см. табл. 4, рис. 5). В локалитетах “Ужгинский”, “Белая Кедва”, “Сойвинский” доминирует B. sporadicus – таежный вид, малочисленный по всему ареалу и нуждающийся в биологическом надзоре на территории Республики Коми [12]. Обособленность в ординационном пространстве локалитета “Немский” (самой крупной из обследованных популяций V. spicata) связана с доминированием B. lucorum s.l. По данным А.М. Бывальцева [20, 21], этот вид избирателен в фуражировке и связан с видами рода Veronica. Два вида доминантного комплекса опылителей V. spicata (B. lucorum s.l. и B. jonellus), а также не столь многочисленный B. veteranus отмечены среди активных опылителей этого вида и в других частях ареала [18, 20, 21].

ЗАКЛЮЧЕНИЕ
На Севере комплекс опылителей V. spicata, вида специализированного мелиттофильного комплекса, насчитывает 19 видов насекомых из 2 отрядов и 3 семейств. Преобладание в нем широко распространенных полилектичных видов связано с относительной молодостью и преимущественно миграционным характером энтомофауны Европейского Северо-Востока России [40].
Показано, что структура комплекса определяется особенностями строения цветка V. spicata, дифференциацией его экологических ниш на северном пределе распространения и широтным фактором. В отличие от других частей ареала, где важную роль в опылении вида играют одиночные пчелы [18], на Севере ведущая роль принадлежит представителям рода Bombus Latr. С учетом уровня видового разнообразия и структуры населения основу комплекса составляют короткохоботковые виды шмелей: B. lucorum s.l., B. jonellus, B. pratorum, B. sporadicus, а также среднехоботковый B. pascuorum.
Подтверждены различия комплекса опылителей V. spicata в двух типах местообитаний вида на Севере – в интразональных сообществах Pinetum cladinosum на боровых террасах и азональных выходах кальцийсодержащих пород с реликтовым скальным флористическим комплексом. Такая дифференциация экологических ниш V. spicata связана с историей развития растительного покрова на Европейском Северо-Востоке России [34, 41]. В сообществах Pinetum cladinosum большая площадь популяций и высокое обилие V. spicata, в сочетании с низкими показателями видового разнообразия и обилия других растений, создают предпосылки для олиголектичности шмелей. На выходах известняков реализуется иная стратегия фуражировки – шмели одновременно питаются на целом ряде растений скального комплекса. С учетом небольших размеров и численности популяций V. spicata такое значительное давление в конкуренции за опылителей может приводить к попаданию недостаточного количества пыльцы на цветки этого вида.
Работа выполнена в рамках государственного задания (№ тем 122040600026-9 и 122040600025-2). Авторы признательны А.Б. Новаковскому за консультации и помощь в обработке материалов.
Список литературы
Панфилов Д.В. Общий обзор населения пчелиных Евразии // Сборник трудов Зоологического музея Московского ун-та. 1968. Т. 11. С. 18–35.
Фэгри К., Ван дер Пейл Л. Основы экологии опыления. М.: Мир, 1982. 344 с.
Liu Y., Jin H., Naeem M., An J. Comparative transcriptome analysis reveals regulatory genes involved on cold tolerance and hypoxic adaptation of high-altitude Tibetan bumblebees // Apidologie. 2020. V. 51. P. 1166–1181. https://doi.org/10.1007/s13592-020-00795-w
Potts S.G., Imperatriz-Fonseca V., Ngo H.T. et al. Safeguarding pollinators and their values to human well-being // Nature. 2016. V. 540. P. 220–229. https://doi.org/10.1038/nature20588
Soroye P., Newbold T., Kerr J. Climate change contributes to widespread declines among bumble bees across continents // Science. 2020. V. 367. Is. 6478. P. 685–688. https://doi.org/10.1126/science.aax8591
Gagic V., Kirkland L., Kendall L.K. et al. Understanding pollinator foraging behaviour and transition rates between flowers is important to maximixe seed set in hybrid crops // Apidologie. 2021. V. 52. P. 89–100. https://doi.org/10.1007/s13592-020-00800-2
Wei N., Kaczorowski R.L., Arceo-Gómez G. et al. Pollinators contribute to the maintenance of flowering plant diversity // Nature. 2021. V. 597. P. 688–692. https://doi.org/10.1038/s41586-021-03890-9
Kwak M.M., Bekker R.M. Ecology of plant reproduction: extinction risks and restoration perspectives of rare plant species// Plant-pollinator interactions. From specialization to generalization. Eds. Waser N.M., Ollerton J. Chicago: The University of Chicago Press, 2006. 445 p. https://doi.org/10.1016/j.anbehav.2006.08.001
Zych M., Jakubiec A. Pollination of Polish Red list plants: a preliminary statistical survey // Acta agrobotanica. 2008. V. 61. Is. 1. P. 85–90.
Демьянова Е.И. О системах скрещивания у охраняемых растений Среднего Урала // Вестник Пермского ун-та. 2015. Вып. 2. С. 91–126.
Юдин Ю.П. Реликтовая флора известняков Северо-Востока европейской части СССР // Материалы по истории флоры и растительности СССР. М., Л.: Изд-во АН СССР, 1963. Вып. 4. С. 493–587.
Красная книга Республики Коми. Сыктывкар: Коми кн. изд-во, 2019. 768 с.
Красная книга Ханты-Мансийского автономного округа - Югры: животные, растения, грибы. Екатеринбург: Баско, 2013. 460 с.
Красная книга Ненецкого автономного округа. Нарьян-Мар: Администрация Ненецкого автономного округа, 2019. 455 с.
Wilson G.B., Houston L., Whittington W.J., Humphries R.N. Veronica spicata L. ssp. spicata and ssp. hybrida (L.) Gaudin (Pseudolysimachium spicatum (L.) Opiz) // J. of Ecology. 2001. V. 88. № 5. P. 890–909.
Scalone R., Kolf M., Albach D.C. Mating system variation in Veronica (Plantaginaceae): inferences from pollen/ovule ratios and other reproductive traits // Nordic J. of Botany. 2012. V. 31. Is. 3. P. 372–384. https://doi.org/10.1111/j.1756-1051.2012.01706.x
Демьянова Е.И., Пономарев А.Н. К антэкологии некоторых степных растений лесостепного Зауралья // Экология опыления. Пермь: Изд-во Пермского ун-та, 1979. С. 113–125.
Banaszak J. Pszczoły (Apoidea, Hymenoptera) siedlisk kserotermicznych rejonu dolnej Wisły // Fragmenta faunistica. 1980. V. 25. № 19. P. 335–360.
Fisogni A., Massol F., de Manincor N. et al. Network analysis highlights increased generalisation and evenness of plant-pollinator interactions after conservation measures // Acta Oecologica. 2021. V. 110. P. 1–10. https://doi.org/10.1016/j.actao.2020.103689
Бывальцев А.М. Сообщества шмелей (Hymenoptera, Apidae, Bombini) в Кулундинской равнине Западной Сибири // Евразиатский энтомол. журн. 2013. Т. 12. № 6. С. 575–586.
Бывальцев А.М., Белова К.А., Проскурякова А.А. Население и экология шмелей Прииртышской степи в раннелетний период (Hymenoptera: Apidae, Bombus) // Вестник Новосибирского гос. ун-та. Серия: Биология, клиническая медицина. 2013. Т. 11. Вып. 1. С. 40–46.
Ценопопуляции растений (Основные понятия и структура). М.: Наука, 1976. 215 с.
Серебрякова Т.И., Каргалицкая Т.Н. Большой жизненный цикл и эволюционные отношения жизненных форм некоторых видов Veronica L. секции Pseudolysimachium Koch. // Бюлл. МОИП. Отд. биол. 1972. Т. 77. № 6. С. 81–98.
Юрцев Б.А. Флора Сунтар-Хаята. Проблемы истории высокогорных ландшафтов Северо-Востока Сибири. Л.: Наука, 1968. 235 с.
Флора Северо-Востока европейской части СССР. Л.: Наука, 1974. Т. 1. 273 с.; 1976. Т. 2. 315 с.; 1976. Т. 3. 293 с.; 1977. Т. 4. 311 с.
Городков К.Б. Типы ареалов насекомых тундры и лесных зон европейской части СССР / Ареалы насекомых европейской части СССР. Л.: Наука, 1984. С. 3–20.
Williams P.H. Bumblebees of the World. Natural History Museum. URL:https://www.nhm.ac.uk/researchcuration/research/projects/bombus/subgenericlist.html (дата обращения: 28.06.2022).
Песенко Ю.А. Принципы и методы количественного анализа в фаунистических исследованиях. М.: Наука, 1982. 287 с.
Новаковский А.Б. Взаимодействие Excel и статистического пакета R для обработки данных в экологии // Вестник Института биологии Коми НЦ УрО РАН. 2016. № 3. С. 26–33. https://doi.org/10.31140/j.vestnikib.2016.3(197).4
Цыганов Д.Н. Фитоиндикация экологических режимов в подзоне хвойно-широколиственных лесов. М.: Наука, 1983. 198 с.
Экологические шкалы и методы анализа экологического разнообразия растений / Ред. Жукова Л.А. Йошкар-Ола: Изд-во Марийского гос. ун-та, 2010. 368 с.
Лесорастительное районирование Республики Коми / Козубов Г.М., Мартыненко В.А., Дегтева С.В. и др. // Леса Республики Коми. М.: Издат.-продюс. центр “Дизайн. Информация. Картография”, 1999. С. 257–288.
Тетерюк Л.В., Елсаков В.В., Лаптева Е.М. Роль рельефа в формировании терморежима и биоразнообразия реликтовых экосистем на известняках Европейского Северо-Востока России // Экология. 2012. № 6. С. 410–416. [Teteryuk L.V., Elsakov V.V., Lapteva E.M. The role of topography in forming the thermal regime and biological diversity of relict ecosystems on limestones in the Northeast of European Russia // Russ. J. of Ecology. 2012. V. 43. № 6. P. 433–439.] https://doi.org/10.1134/S1067413612060082
Еленевский Л.Г. Систематика и география вероник СССР и прилежащих стран. М.: Наука, 1978. 259 с.
Татаринов А.Г., Долгин М.М. Видовое разнообразие булавоусых чешуекрылых на Европейском Северо-Востоке России. СПб.: Наука, 2001. 244 с.
Kampny C.M. Pollination and flower diversity in Scrophulariaceae // The Botanical Review. 1995. V. 61. № 4. P. 350–366.
Teräs I. Food plants and flower visits of bumblebees (Bombus: Hymenoptera, Apidae) in southern Finland. Helsinki, 1985. 84 p.
Длусский Г.М. Значение конкуренции за опыление в формировании структуры комплекса энтомофильных растений // Журн. общ. биол. 2013. Т. 74. № 6. С. 434–449.
Pojar J. Reproductive dynamics of four plant communities of southwestern British Columbia // Canadian Journal Botany. 1974. V. 52. P. 1819–1834.https://doi.org/10.1139/b74-234
Крыжановский О.Л. Состав и распространение энтомофаун земного шара. М.: Тов-во науч. изд. КМК, 2002. 237 с.
Горчаковский П.Л. Основные проблемы исторической фитогеографии Урала // Труды Института экологии растений и животных УФАН СССР. Свердловск: Урал. филиал АН СССР, 1969. Вып. 66. 286 с.
Дополнительные материалы отсутствуют.