

Экология, 2023, № 1, стр. 13-21
Многолетний мониторинг растительного покрова на осушенных болотах Южной Карелии (на примере мезотрофного травяно-сфагнового болота)
С. И. Грабовик a, Л. В. Канцерова a, *
a ФГБУН Институт биологии Карельского научного центра РАН
185910 Петрозаводск, ул. Пушкинская, 11, Россия
* E-mail: Kancerova.L@mail.ru
Поступила в редакцию 08.04.2022
После доработки 01.09.2022
Принята к публикации 05.09.2022
- EDN: GZJTTE
- DOI: 10.31857/S0367059723010067
Аннотация
При помощи градиентного анализа и экошкал Элленберга выявлены факторы дифференциации растительного покрова (увлажнение, освещенность, кислотность почвы, богатство почвы минеральным азотом). С помощью неметрического шкалирования была создана ординационная диаграмма распределения геоботанических описаний за 50 лет исследования (с 1971 г. по 2021 г.). Проведена корреляция осей ординации и экологических факторов. На ординационной диаграмме и по результатам кластерного анализа геоботанические описания образуют три ярко выраженные группы, соответствующие определенным временны́м периодам. Причиной смен видового состава в растительных сообществах являются изменения освещенности, увлажнения и трофности. Под влиянием осушения и сукцессий растительного покрова в течение 50 лет на месте мезотрофного кочковато-топяного травяно-сфагнового комплекса, представленного мезотрофными гидрогигрофильными болотными видами, сформировался олиготрофный кочковато-равнинный сосняк ерниково-сфагновый, образованный олиготрофными, мезогигрофильными лесо-болотными видами.
В Республике Карелия болота занимают 3.63 млн га (21%), из них на долю травяно-сфагново-гипновых (карельские кольцевые аапа болота) приходится 0.9 млн га [1]. Одним из наиболее распространенных способов трансформации болотных экосистем таежной зоны является лесоосушительная мелиорация, в результате которой происходит изменение гидрологического режима болотных массивов: на некоторых осушенных площадях проводили обработку почвы и создавали лесные культуры, ряд участков оставляли для естественного облесения. В 60–70-е годы XX в. 25% общей площади болот Карелии было осушено для целей лесного и сельского хозяйства, в отдельных районах эта цифра достигает 70% [2, 3].
Изменению растительности болот и заболоченных лесов под влиянием осушения посвящено достаточно много работ как у нас в стране [4–7], так и за рубежом [8–11]. Анализ литературы показывает, что большая часть материалов об изменении растительного покрова болот и заболоченных лесов получена на основе однократных учетов, проведенных через достаточно большой период после осушения. При этом динамика напочвенного покрова рассматривается в сравнительно немногих работах, в основном на болотах европейской части России [12–14]. В Западной Сибири [4, 15] основное внимание уделяется реакции на осушение древесного яруса или его формированию на открытых болотах.
Основная цель настоящей работы – анализ результатов 50-летних исследований постмелиоративной динамики видового состава растительного покрова, а также выявление основных экологических факторов мезотрофного травяно-сфагнового болотного массива во временно́м аспекте. В задачи исследования входило рассмотреть разнообразие растительного покрова осушенного болотного участка и провести градиентный анализ с выявлением ведущих факторов их дифференциации.
МАТЕРИАЛ И МЕТОДЫ
На территории Европейского Севера России наиболее распространены четыре типа болотных массивов [16]: сфагновый грядово-мочажинный олиготрофный, травяно-сфагновый мезотрофный, травяно-сфагново-гипновый (карельские кольцевые аапа болота) и мезотрофный сфагновый лесной (рис. 1). Значительная их часть в южной и средней Карелии была осушена в 60–70-е годы XX в.
Рис. 1.
Мезотрофный травяно-сфагновый болотный массив: a – географическое положение болота в пределах Карелии; б – схема болотных массивов стационара “Киндасово”: I – мезотрофный травяно-сфагновый, II – мезотрофный сфагновый лесной, III – южнокарельский вариант кольцевого аапа типа, IV – олиготрофный сфагновый грядово-мочажинный, V – ручьи; VI – река Шуя; VII – пробная площадь; 1–8 – болотные массивы: 1 – Неназванное, 2 – Мустусуо, 3 – Койвусуо, 4 – Ритусуо, 5 – Райское, 6 – Деловое, 7 – Близкое, 8 – Березовое; в – центральная часть болота Койвусуо: 1 – пробная площадь (0.4 га), 2 – постоянная пробная площадь (50 м2), 3 – постоянная метровая площадь (1 м2), 4 – мелиоративные каналы.
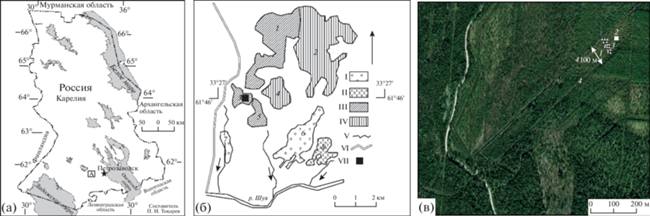
В 1969 г. под руководством члена-корр. АН СССР Н.И. Пьявченко были начаты комплексные научно-исследовательские работы по изучению структуры и динамики болот и заболоченных лесов в естественном состоянии и под влиянием мелиорации на территории Южной Карелии в подзоне средней тайги на лесоболотном научном стационаре “Киндасово” в заказнике Койву-Ламбасуо. В климатическом отношении территория стационара характеризуется следующими средними многолетними данными: продолжительность вегетационного периода – 148 дней, температура воздуха за вегетационный период – 11.7°С, количество атмосферных осадков за год – 565 мм, за вегетационный период – 316 мм [17].
Мы рассматриваем динамику видового состава растительного покрова мезотрофного травяно-сфагнового болотного массива Койвусуо площадью 40 га, который относится к южно-карельскому варианту аапа болот и имеет торфяную залежь мощностью около 1.5 м. Пробная площадь болотного массива составляет 0.4 га (61°46′22.7″ с.ш. и 33°29′22.9″ в.д.). Структуру растительного покрова изучали на постоянной пробной площади (50 м2), заложенной в центральной части болотного массива в мезотрофном кочковато-топяном травяно-сфагновом комплексе, который занимал 27% от площади массива. Мониторинг изменений видового состава велся на 15 постоянных площадках размером 1 м2, заложенных на понижениях, которые были расположены рассеянно в пределах пробной площади изучаемого болотного комплекса. На постоянных метровых площадках проективное покрытие видов (в %) травяно-кустарничкового и мохового ярусов оценивали с периодичностью 1–2 года, реже до 7 лет. Первые описания были сделаны в год осушения. В задачи исследования не входило измерение уровня почвенно-грунтовых вод (УПГВ) в течение вегетационного периода, но на постоянной пробной площади был измерен УПГВ в понижениях до осушения, а также через 25 и 50 лет.
Исследования динамики видового состава и структуры растительного покрова выполняли апробированными и модифицированными методами [18, 19]. Таксономия сосудистых растений приводится по [20] с учетом более поздней работы [21], мхов – по [22]. Экологические группы сосудистых растений и мхов по факторам обводненности, трофности и приуроченности к экотопам приводятся по работам [23, 24] Для установления экологических особенностей выделенных сообществ и определения их положения в экологическом пространстве был использован градиентный анализ, выполненный методом неметрического шкалирования (NMS) [25] в прикладном пакете для обработки геоботанических данных PC-ORD 6.12. Нагрузки на оси ординации рассчитаны after-the-fact посредствам сравнения дисперсии, объясняемой отдельной осью с общей дисперсией в таблице эмпирических данных [26]. В качестве анализируемого параметра использовался показатель проективного покрытия видов (в %) для растительных сообществ постоянных метровых площадок за разные годы исследования. Поскольку прямые измерения факторов среды отсутствовали, для интерпретации ординационных градиентов были использованы значения экологических факторов, рассчитанные методом экологических шкал Х. Элленберга [27]. Характеристика в шкалах Элленберга представлена для 67% видов из общего списка. Кластеризация материала была проведена методом гибкой беты (flexible beta) при значении β = –0.25 [28]. Для вычисления матрицы расстояний использовалась относительная дистанция Сьеренсена.
РЕЗУЛЬТАТЫ
При помощи градиентного анализа и по шкалам Элленберга выявлены 4 основных фактора дифференциации растительного покрова в каждый год исследования (табл. 1). Отмечены незначительные уменьшения степени освещенности (от 8.3 – полностью световые растения до 7.5 – преимущественно световые растения, в основном произрастают на освещенных участках, но иногда встречаются в затененных местах); степени увлажнения почвы (от 9 – эти растения растут на сырых, плохо аэрируемых почвах до 8.5 – растения растут на влажных, не просыхающих почвах); кислотности почвы (от 2.5 до 2.1 – растения растут на кислых почвах) и богатства почвы минеральным азотом (от 2.4 до 1.6 – растения растут на бедных азотом почвах) [27].
Таблица 1.
Основные экологические факторы растительных сообществ мезотрофного травяно-сфагнового болотного массива за 50 лет исследования (по экошкалам Элленберга). Пунктирные линии указывают на переходы между периодами осушения (рис. 2)
| Годы исследования | Экологический фактор | |||
|---|---|---|---|---|
| L* | F | R | N | |
| 1971 | 8.3 | 9.0 | 2.5 | 2.4 |
| 1972 | 8.3 | 9.1 | 2.2 | 2.4 |
| 1973 | 8.2 | 9.1 | 2.6 | 2.5 |
| 1975 | 8.1 | 9.0 | 2.4 | 2.2 |
| 1977 | 8.4 | 9.1 | 2.6 | 2.5 |
| 1978 | 8.3 | 8.8 | 2.7 | 2.7 |
| Средние значения I периода | 8.3 | 9.0 | 2.5 | 2.4 |
| 1984 | 8.2 | 9.0 | 2.5 | 1.9 |
| 1985 | 8.2 | 9.1 | 2.6 | 2.1 |
| 1986 | 7.7 | 8.5 | 2.1 | 1.9 |
| 1989 | 8.0 | 9.1 | 2.7 | 2.1 |
| 1990 | 8.0 | 9.1 | 2.3 | 2.1 |
| 1992 | 8.1 | 9.0 | 2.4 | 1.9 |
| 1996 | 7.8 | 8.5 | 2.4 | 2.0 |
| 1997 | 8.0 | 8.9 | 2.3 | 2.0 |
| 1998 | 7.9 | 8.9 | 2.3 | 2.1 |
| 1999 | 7.6 | 8.7 | 2.1 | 1.9 |
| 2000 | 7.6 | 8.9 | 2.2 | 1.9 |
| Средние значения II периода | 7.9 | 8.8 | 2.4 | 2.0 |
| 2001 | 7.8 | 8.8 | 2.4 | 1.9 |
| 2002 | 7.7 | 8.7 | 2.4 | 2.0 |
| 2003 | 7.6 | 8.7 | 1.9 | 1.8 |
| 2004 | 7.8 | 8.6 | 2.1 | 1.9 |
| 2007 | 7.7 | 8.6 | 1.7 | 1.6 |
| 2009 | 7.5 | 8.5 | 2.3 | 1.6 |
| 2016 | 7.5 | 8.5 | 2.1 | 1.5 |
| 2018 | 7.5 | 8.5 | 2.1 | 1.5 |
| 2021 | 7.5 | 8.5 | 2.1 | 1.6 |
| Средние значения III периода | 7.6 | 8.6 | 2.1 | 1.7 |
Примечание. Здесь и в табл. 2: L – освещение, F – увлажнение почвы, R – кислотность почвы, N – богатство почвы азотом.
С помощью неметрического шкалирования (NMS) была создана ординационная диаграмма распределения геоботанических описаний за 50 лет (рис. 2). Проведена корреляция осей ординации и экологических факторов. Первая ось показывает 61.8% общей дисперсии, вторая – 32.6%. Корреляция факторов с первой осью не проявилась, зато заметна сильная отрицательная корреляция второй ординационной оси с богатством почв минеральным азотом (r = –0.840), освещенностью (r = –0.826), немного меньше с кислотностью почв (r = –0.733) и увлажненностью (r = –0.645) (табл. 2).
Рис. 2.
NMS-ординация растительных сообществ на постоянных метровых площадках за 50 лет исследований. Римские цифры – периоды увлажнения: I – 1971–1978 гг.; II – 1984–2000 гг.; III – 2001–2021 гг. Векторы экологических значений (см. в табл. 1): L – освещение, F – увлажнение почвы, R – кислотность почвы, N – богатство почвы азотом.
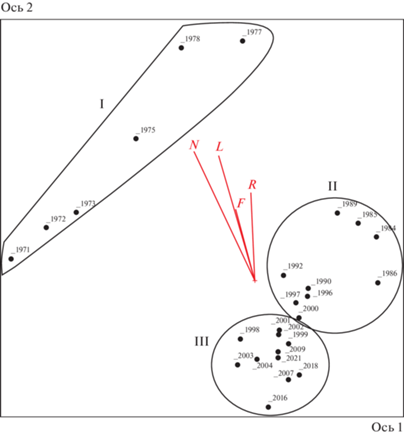
Таблица 2.
Коэффициенты корреляции Пирсона (r) и Кендалла (τ) ординационных осей с факторами среды по экошкалам Элленберга
| Экологический фактор | Ось 1 | Ось 2 | ||
|---|---|---|---|---|
| r | τ | r | τ | |
| L | –0.445 | –0.141 | –0.826 | –0.698 |
| F | –0.326 | –0.103 | –0.625 | –0.606 |
| R | –0.159 | –0.026 | –0.733 | –0.680 |
| N | –0.576 | –0.206 | –0.840 | –0.708 |
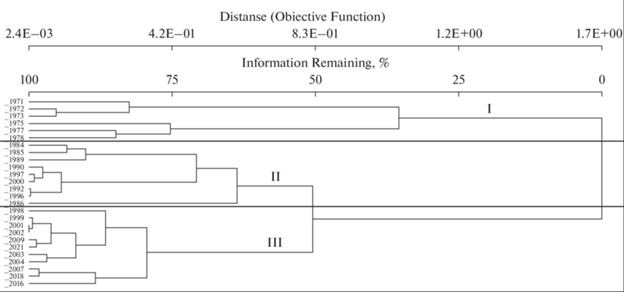
На ординационной диаграмме и по результатам кластерного анализа (рис. 3) геоботанические описания образуют три ярко выраженные группы, соответствующие определенным временны́м периодам. Группы (периоды) достаточно четко очерчены в ординационном пространстве NMS и не перекрываются друг с другом. Разрыв между I и II группами может быть связан с отсутствием данных за длительный период времени. Изменения факторов освещенности и трофности являются основными причинами смен видового состава на местообитаниях, подвергшихся осушению, а фактор увлажнения зависит от степени трофности.
Рис. 3.
Кластерный анализ растительных сообществ на постоянных метровых площадках за 50 лет исследований. Периоды увлажнения: I – 1971–1978 гг.; II – 1984–2000 гг.; III – 2001–2021 гг.
До осушения (1971 г.). Болото осушено в 1971 г., расстояние между осушительными каналами 100 м, в настоящее время каналы заросли травянистой растительностью. До осушения во флористическом составе участка отмечено 20 видов: деревьев – 2, кустарничков – 4, трав – 8, мхов – 6. Отдельные сосны и березы, высотой 1–2 м, были приурочены к осоково-сфагновым кочкам.
Для изучения изменений структуры растительного покрова в центральной части участка было выполнено крупномасштабное картирование. Кочки занимали 36%, растительный покров которых был представлен фитоценозами Andromeda polifolia–Carex lasiocarpa–Sphagnum divinum + S. angustifolium. Сообщества ковров Carex lasiocarpa–Sphagnum subsecundum и топей Carex limosa–Menyanthes trifoliata занимали 64% (табл. 3). Проективное покрытие кустарничково-травяного яруса повышений – 80%, мхов – 100%, понижений – соответственно 65 и 55%.
Таблица 3.
Изменение структуры растительного покрова мезотрофного кочковато-топяного травяно-сфагнового комплекса под влиянием осушения
| Фитоценозы | Микрорельеф | До осушения, % |
Периоды осушения | ||||
|---|---|---|---|---|---|---|---|
| I | II | III | |||||
| Год картирования | 1975 г. | 1984 г. | 2000 г. | 2007 г. | 2018 г. | ||
| Andromeda polifolia–Carex lasiocarpa–Sphagnum divinum + S. angustifolium | Кочки | 36 | – | – | – | – | – |
| Betula nana + Andromeda polifolia–S. divinum + S. angustifolium | То же | – | 33 | – | – | – | – |
| Betula nana–S. angustifolium + S. divinum | » | – | – | 35 | – | – | – |
| Betula nana–Polytrichum strictum + Polytrichastrum longisetum | » | – | – | 15 | 10 | 10 | 10 |
| Carex lasiocarpa–S. subsecundum | » | 18 | – | – | – | – | – |
| Carex limosa–Menyanthes trifoliate | Топи | 46 | – | – | – | – | – |
| Carex lasiocarpa–Menyanthes trifoliate | Ковры | – | 67 | – | – | – | – |
| Carex lasiocarpa–S. angustifolium | То же | – | – | 50 | – | – | – |
| Pinus sylvestris–Betula nana–S. angustifolium | » | – | – | – | 90 | 90 | 90 |
В нашей работе анализ динамики растительного покрова приводится на примере растительных сообществ Carex lasiocarpa–Sphagnum subsecundum, приуроченных к коврам, и Carex limosa– Menyanthes trifoliata, приуроченных к топям (см. табл. 3); УПГВ – 2–3 см ниже поверхности сфагнового покрова.
I период осушения (1971–1978 гг.). В данный период осушения на месте прежнего кочковато-топяного комплекса сформировался кочковато-равнинный (см. табл. 3). Кочки Betula nana + Andromeda polifolia–S. divinum + S. angustifolium занимают 33%, ковры Carex lasiocarpa–Menyanthes trifoliata – 67%. Растительный покров ковров представлен мезотрофными гидрогигрофильными болотными видами сосудистых растений и мхов. Диагностическими видами сообществ данного периода являются Andromeda polifolia, Carex lasiocarpa, C. chordorrhiza, Menyanthes trifoliata, Equisetum fluviatile, Sphagnum angustifolium, S. divinum. Эти виды по-прежнему сохраняют высокую константность, но проективное покрытие их снижается (табл. 4).
Таблица 4.
Постмелиоративная динамика видового состава мезотрофного кочковато-топяного травяно-сфагнового комплекса
| Видовой состав | Периоды осушения и годы | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| I | II | III | |||||||||||||
| 1971* | 1972 | 1973 | 1975 | 1978 | 1984 | 1986 | 1990 | 1997 | 2000 | 2001 | 2007 | 2009 | 2018 | 2021 | |
| Andromeda polifolia | V8 | V9 | IV 7 | V8 | V5 | II2 | IV 2 | V5 | IV 2 | IV 5 | IV 3 | V5 | IV 3 | V3 | V3 |
| Betula nana | IV6 | IV 7 | IV 7 | IV 12 | IV 14 | V37 | V28 | V24 | V27 | V28 | V32 | V31 | V25 | V26 | V23 |
| Chamaedaphne calyculata | II+ | II1 | II1 | II+ | II+ | – | – | – | – | I+ | – | – | – | I+ | I+ |
| Oxycoccus palustris | IV4 | V 6 | IV 5 | V 3 | III 1 | III8 | V4 | V16 | V13 | V17 | V14 | V18 | V14 | V12 | V10 |
| Salix myrtilloides | I+ | – | – | – | – | – | – | I+ | I+ | II+ | II2 | II+ | II+ | II2 | II2 |
| Vaccinium uliginosum | – | – | – | – | – | – | – | I+ | I+ | I+ | – | I1 | I2 | II+ | II+ |
| V. vitis-idaea | – | – | – | – | – | – | III+ | – | I1 | I1 | II1 | II2 | III3 | III4 | III3 |
| Carex chordorrhiza | V21 | V16 | V8 | V5 | III+ | – | – | – | – | – | – | – | – | I+ | I+ |
| C. dioica | – | – | – | – | – | – | – | – | – | – | – | – | I+ | I+ | I+ |
| C. lasiocarpa | V15 | V16 | V12 | IV7 | V11 | V5 | III2 | V2 | V+ | II+ | V+ | III+ | I+ | II+ | II+ |
| C. limosa | III8 | III8 | – | – | II+ | – | – | – | – | – | – | – | – | – | – |
| C. pauciflora | – | – | – | – | – | – | – | – | – | – | – | – | II+ | II+ | II+ |
| C. paupercula | – | – | – | – | – | – | – | – | – | – | – | – | – | I+ | I+ |
| Chamaenerion angustifolium | – | – | – | – | II+ | – | – | – | – | – | – | – | – | – | – |
| Comarum palustre | I+ | I+ | – | – | – | – | – | – | – | – | – | – | – | – | – |
| Drosera rotundifolia | I+ | – | – | – | – | II+ | – | – | – | – | – | I+ | – | I+ | I+ |
| Equisetum fluviatile | IV1 | V+ | IV+ | II+ | II+ | – | II+ | II+ | II+ | II+ | II+ | II+ | – | – | – |
| Eriophorum angustifolium | V5 | IV3 | IV2 | IV2 | III1 | IV9 | IV1 | IV2 | III+ | IV2 | V2 | – | IV+ | II+ | II+ |
| E. vaginatum | – | – | – | – | – | – | – | I+ | – | II1 | II+ | I+ | I+ | IV+ | IV+ |
| Menyanthes trifoliata | IV4 | V7 | V2 | V3 | V3 | I+ | I+ | I+ | – | – | – | – | – | – | – |
| Aulacomnium palustre | II+ | I+ | – | I+ | I+ | – | – | III2 | II+ | II+ | I+ | I+ | – | IV1 | IV+ |
| Dicranum polysetum | – | – | – | – | – | – | – | – | – | – | – | – | – | I+ | I+ |
| Pleurozium schreberi | I4 | – | I+ | – | – | – | II28 | – | I+ | – | – | I+ | – | III2 | III2 |
| Polytrichum commune | – | – | – | – | – | – | II1 | III8 | I1 | – | II3 | – | II7 | II+ | II+ |
| Polytrichum strictum | II3 | I+ | – | – | III3 | V39 | IV21 | V36 | V34 | V36 | V21 | V17 | V18 | V20 | V19 |
| Sphagnum angustifolium | III19 | IV18 | IV19 | III12 | III7 | II1 | III12 | V26 | V25 | V30 | V32 | V57 | V39 | V54 | V53 |
| S. capillifolium | – | – | – | – | – | – | – | – | – | III3 | II3 | II4 | – | III3 | III3 |
| S. divinum | IV31 | V31 | IV28 | III12 | III2 | IV6 | IV4 | V7 | V15 | V13 | V17 | V10 | V12 | V7 | V6 |
| S. fuscum | – | – | – | – | – | – | – | – | I2 | II6 | II6 | II2 | – | II1 | II1 |
| S. subsecundum | III34 | – | II2 | – | – | – | – | – | – | – | – | – | – | – | – |
II период осушения (1984–2000 гг.). Во втором периоде отмечено дальнейшее сглаживание микрорельефа. Через 13 лет (1984 г.) после осушения кочки занимают 50%, а через 30 лет (2000 г.) – 10%; напочвенный покров ковров через 13 лет сменяется на Carex lasiocarpa–S. angustifolium (50%), а через 30 лет – на Pinus sylvestris–Betula nana–S. angustifolium (90%) (см. табл. 3).
Растительный покров ковров образован сообществами мезоолиготрофных, гигрофильных болотных видов. Различные виды болотных растений реагируют на осушение неодинаково. Из растительного покрова исчезли гидрогигрофильные и гигрофильные виды Chamaedaphne calyculata, Carex limosa, C. chordorrhiza, Sphagnum subsecundum, а другие испытывают явное угнетение и отмирают постепенно – Menyanthes trifoliata и Equisetum fluviatile (см. табл. 4). Встречаются виды, которые развиваются даже лучше, и их роль в покрове заметно увеличивается после осушения: Betula nana, Oxycoccus palustris, Sphagnum angustifolium, S. divinum. Проективное покрытие в растительных сообществах в отличие от первых лет осушения резко возрастает – в среднем с 5–10% до 20–50%. В связи с улучшением аэрации корнеобитаемого слоя Betula nana усиленно разрастается. В сообществах на коврах разрастаются Vaccinium vitis-idaea и V. uliginosum, а также Sphagnum fuscum и S. capillifolium, которые ранее росли на высоких кочках. Зеленые мхи, а именно Polytrichum commune и P. strictum, полностью заселяют участки с мертвым Sphagnum subsecundum, также здесь поселяется и Polytrichastrum longisetum (УПГВ – 25–30 см). В это же время здесь началось поселение сосны. Общее число стволов составляет до 1103 экз/га, в том числе сосны – 937, березы – 166.
III период осушения (2001–2021 гг.). В данный период кочки занимали 10%, а на коврах сформировалось сосново-ерниково-сфагновое сообщество (Pinus sylvestris–Betula nana–S. angustifolium (90%)) (см. табл. 3). Растительный покров ковров образован олиготрофными, мезогигрофильными лесоболотными видами. Растения гигрофильные угнетаются, конкуренция с их стороны практически устраняется. В напочвенном покрове за этот период осушения большинство болотных видов (Betula nana, Carex lasiocarpa, Eriophorum angustifolium, Polytrichum strictum) продолжают сохраняться, но жизненность и проективное покрытие их снижаются. С застойным режимом увлажнения и значительным снижением уровня почвенно-грунтовых вод (до 35–50 см ниже поверхности сфагнового покрова) связано исчезновение Equisetum fluviatile. Происходит расселение Sphagnum angustifolium по коврам, проективное покрытие его возрастает в два раза. В данный период осушения происходит внедрение лесоболотных видов (Carex dioica, C. poupercula, C. pauciflora, Eriophorum vaginatum, Dicranum polysetum), и в сложении напочвенного покрова они уже играют значительную роль. Эти виды указывают на переменность водного режима и динамические связи болот и лесов.
Общее количество стволов деревьев 1766 шт/га, в том числе сосны – 1233, ели – 125, березы – 408. Относительная полнота древостоя 0.4.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
В данной работе для мезотрофных травяно-сфагновых болот Южной Карелии с помощью градиентного анализа и экошкал Элленберга выявлена общая направленность растительных сукцессий в течение 50 лет после осушения, с акцентом внимания на детальные экологические характеристики растительных сообществ и изменения видового состава в них. Наши исследования позволяют полнее раскрыть механизм влияния мелиорации не только на древесный ярус, но и на почвенный покров и экологические условия осушенных участков.
Анализ литературы показывает, что большая часть материалов об изменении растительного покрова болот и заболоченных лесов получена на основе однократных учетов, проведенных через достаточно большой период после осушения. Финские исследователи Л. Хейкурайнен [29] и Ю. Лайне [30] отмечают, что на осушенных болотах напочвенный покров достигает устойчивого, отличного от первоначального типа, растительного сообщества только спустя более 40 лет после осушения. По нашим данным, характер происходящих изменений и чередование этапов динамики растительного покрова после осушения разнообразны и зависят не только от степени осушения, но и от исходного типа болотного участка. Растительный покров осушенного мезотрофного травяно-сфагнового болотного участка спустя 50 лет после осушения не достигает устойчивого состояния.
В Западной Сибири С.П. Ефремовым [31] подведены итоги 20-летних стационарных исследований, в которых основное внимание уделено комплексной оценке процесса формирования пионерных древостоев на осушенных безлесных и слабооблесенных болотах. Наши данные показывают, что через 50 лет после осушения на мезотрофном травяно-сфагновом болоте сформировался олиготрофный сосняк ерниково-сфагновый. Общее количество стволов деревьев составило 1766 шт/га, в том числе сосны – 1233, ели – 125, березы – 408. Относительная полнота древостоя 0.4.
Исследования растительного покрова под влиянием осушения безлесных травяно-сфагновых переходных болот и сосняков сфагновых с давностью осушения 30–50 лет [13] показали, что при интенсивном осушении травяно-сфагновых переходных болот доминантами травяно-кустарничкового и мохового ярусов в сформировавшихся производных лесных сообществах стали лесные растения и лесные мхи.
Финские исследователи J. Laine et al. [10] изучали изменения напочвенного покрова в сосняках сфагновых с давностью осушения от 3 до 55 лет. Отмечено, что в первые годы после осушения исчезают гидрофильные осоки. Покрытие болотных кустарничков снижается с возрастанием сомкнутости древесного яруса, а участие сфагновых мхов – с уменьшением освещенности. J. Hotonen et al. [11] изучали влияние лесной мелиорации на 6 аапа болотах, расположенных в северной бореальной зоне на 33 болотных участках. Описания растительности были сделаны в год осушения (1933 г.), а затем в 1943, 1950 и 1994 гг. По их данным, на первоначально влажных лесных болотах, в которых доминировали кустарнички, через 60 лет после осушения напочвенный покров полностью сменился лесными видами, тогда как по нашим данным в сосняке кустарничково-сфагновом [32] до осушения на кочках и коврах преобладали болотные кустарнички, осоки и сфагновые мхи, а в напочвенном покрове ковров и через 50 лет после осушения болотные виды продолжают сохраняться, но жизненность их снижается – они не цветут и не плодоносят.
ЗАКЛЮЧЕНИЕ
Через 50 лет после осушения на месте мезотрофного кочковато-топяного травяно-сфагнового комплекса сформировался олиготрофный кочковато-равнинный сосняк ерниково-сфагновый. На основе обобщенной характеристики растительного покрова болотного массива, NMS-ординации и кластерного анализа за весь период осушения с 1971 г. по 2021 г. при помощи градиентного анализа и экошкал Элленберга выявлены факторы дифференциации растительного покрова (увлажнение, освещенность, кислотность почвы, богатство почвы минеральным азотом). Отмечены незначительные уменьшения степени освещенности, увлажнения, кислотности почвы, а также богатства почвы минеральным азотом. Вдоль временно́го градиента выделены три группы геоботанических описаний, каждая из которой соответствует определенному периоду осушения, которые упорядочены в осях главных экологических факторов.
Причиной смен видового состава в растительных сообществах являются изменения освещенности, увлажнения и трофности. В течение 50 лет после мелиорации на месте кустарничково-травяно-сфагновых сообществ, представленных мезотрофными гидрогигрофильными болотными видами, сформировался сосняк ерниково-сфагновый, образованный олиготрофными, мезогигрофильными лесоболотными видами.
Растительный покров до осушения был представлен 20 видами сосудистых растений и мхов, который через 50 лет после осушения увеличился до 25 видов. В сообществах появляются лесоболотные и лесные виды, характерные для таежных лесов.
Авторы выражают благодарность сотрудникам Института биологии КарНЦ РАН – ведущему биологу П.Н. Токареву за составление карт, Института леса КарНЦ РАН – канд. с. х. наук В.А. Ананьеву и канд. с. х. наук А.Н. Пеккоеву за помощь в полевых исследованиях.
Работа выполнена по госзаданию Института биологии КарНЦ РАН: FMEN-2022-0008 № 122031700449-3.
Авторы подтверждают отсутствие конфликта интересов, а также что настоящая статья не содержит исследований с участием людей или животных в качестве объектов работы.
Список литературы
Пьявченко Н.И., Коломыцев В.А. Влияние осушительной мелиорации на лесные ландшафты Карелии // Болотно-лесные системы Карелии и их динамика. Л.: Наука, 1980. С. 52–71.
Медведева В.М. Формирование лесов на осушенных землях среднетаежной подзоны. Петрозаводск: Кар.- Финн. АН СССР, 1989. 168 с.
Саковец В.И., Германова.Н.И., Матюшкин В.А. Экологические аспекты гидромелиорации в Карелии. Петрозаводск: КарНЦ РАН, 2000. 155 с.
Елисеева В.Н. К вопросу об изменении естественной растительности под влиянием осушения // Изв. Томского отд. ВБО. 1964. Вып. 5. С. 85–87.
Нешатаев В.Ю., Коновалова И.В. Влияние осушения на растительность сфагновых сосняков на верховых торфах // Вестник Ленинградского ун-та. 1986. Сер. 3. Вып. 4. С. 29–34.
Нешатаев В.Ю., Федорчук В.Н. Типы осушенных лесов и их происхождение // Динамическая типология леса. М.: Агропромиздат, 1989. С. 178–192.
Федорчук В.Н., Нешатаев В.Ю., Кузнецова М.Л. Лесные экосистемы северо-западных районов России. Типология, динамика, хозяйственные особенности. СПб.: ЗАО Хромис, 2005. 382 с.
Sarasto Ju. Metsan kasvattamiseksi ojiteeujen soiden aluskasvillisunden rakenteesta ja kehityksesta Suomen etelapuoliskossa // Acta Forestation Fennica. 1957. V. 65(7). P. 1–108.
Буш К.К., Аболинь А.А. Строение и изменение растительного покрова важнейших типов леса под влиянием осушения // Вопросы гидролесомелиорации. Рига: Зинатне, 1968. С. 71–126.
Laine J., Vasander H., Laiho R. Long-term effects of water level drawdown on the vegetation of drained pine mires in southern Finland // J. of Applied Ecology. 1995. V. 32. P. 785–802.
Hotonen J.P., Nousiainen H., Paalamo P. Vegetation succession and diversity on Teuravuoma experimental drainage area in northern Finland // Suo. 1999. V. 50. № 2. P. 55–82.
Ниценко А.А. Наблюдения над изменениями растительного покрова под влиянием осушения // Бот. журн. 1951. Т. 36. № 4. С. 349–355.
Юрковская Т.К. Изменение растительного покрова переходных болот Южной Карелии под влиянием осушения // Уч. зап. Тартуского гос. ун-та. 1963. Вып. 145. № 7. С. 337–345.
Гузлена А.Д. Изменения растительного покрова низинного болота Бейбежи под влиянием осушения и освоения // Уч. зап. Тартуского гос. ун-та. 1963. Вып. 145. № 7. С. 298–305.
Платонов Г.М. Смена растительности болот под влиянием осушения // Взаимоотношения леса и болота. М.: Наука, 1967. С. 128–140.
Елина Г.А., Кузнецов О.Л., Максимов А.И. Структурно-функциональная организация и динамика болотных экосистем Карелии. Л.: Наука, 1984. 128 с.
Чесноков В.А. Изменение стока с заболоченных водосборов Южной Карелии под влиянием лесоосушения: Автореф. дис. … канд. геогр. наук. Л., 1979. 25 с.
Грабовик С.И. Динамика растительного покрова болотных массивов мезотрофного травяно-сфагнового типа под влиянием осушения // Бот. журн. 1989. Вып. 74. № 12. С. 1757–1768.
Грабовик С.И., Канцерова Л.В., Кутенков С.А. Влияние осушения на динамику растительного покрова мезотрофных травяно-сфагновых болот Южной Карелии // Бот. журн. 2019. Вып. 104. № 6. С 888–899. https://doi.org/10.1134/S000681361906005X
Черепанов С.К. Сосудистые растения России и сопредельных государств (в пределах бывшего СССР). СПб.: Наука, 1995. 992 с.
Кравченко А.В. Конспект флоры Карелии. Петрозаводск: КарНЦ РАН, 2007. 403 с.
Ignatov M.S., Afonina O.M., Ignatova E.A. Check-list of mosses of East Europe and North Asia // Arctoa. 2006. V.15. P. 1–130.
Ellenberg H. Zeigerwerte der Gefässpflanzen Mitteleuropas. Gottingen: Golrze, 1974. 97 S.
Цыганов Д.Н. Фитоиндикация экологических режимов в подзоне хвойно-широколиственных лесов. М.: Высшая школа, 1983. 105 с.
Kruskal J.B., Wish M. Multidimensional Scaling. Sage University Paper serieson Quantitative Applications in the Social Sciences, № 07-011. 1978. 93 p.
McCune B., Mefford M.J. PC-ORD. Multivariate Analysis of Ecological Data. Version 6.12. Gleneden Beach. Oregon: MjM Software. 2011.
Ellenberg H., Weber H.E., Dull R.I. et al. Zeigerwärte von Pflanzen in Mitteleuropa (Indicator Values of Plants in CentralEurope). Scripta Geobotanica 18. Göttingen: Golrze, 1991. 248 S.
Lance G.N., Williams W.T. A general theory of classification sorting strategies. I. Hierarchical systems // Computer Journal. 1967. V. 9. P. 373–380.
Хейкурайнен Л. Болота. Перевод с финск. М.: Наука, 1983. 40 с
Laine J. Metsaojitetujen soiden luokittelu (English summary: Classification of peatlands drained for forestry) // Suo. 1989. V. 40. № 2. P. 37–51.
Ефремов С.П. Пионерные древостои осушенных болот. Новосибирск: Наука, 1987. 248 с.
Grabovik S.I., Kantserova L.V., Ananyev V.A. Changes in plant cover of a mire in southern Karelia, Russia over 50 years following drainage.// Mires and Peat. 2021. V. 27. № 24. 11 p. https://doi.org/10.19189/MaP.2021.OMB.StA.2161
Дополнительные материалы отсутствуют.