

Экология, 2023, № 4, стр. 291-301
Почвенные нематоды северных лугов и агроценозов как биоиндикаторы степени трансформации почвенных экосистем
Е. М. Матвеева a, А. А. Сущук a, *, Д. С. Калинкина a
a Институт биологии Карельского научного центра РАН, ФИЦ “КарНЦ РАН”
185910 Петрозаводск, ул. Пушкинская, 11, Россия
* E-mail: anna_sushchuk@mail.ru
Поступила в редакцию 01.08.2022
После доработки 02.03.2023
Принята к публикации 05.03.2023
- EDN: RQJBXO
- DOI: 10.31857/S0367059723040078
Аннотация
Изучено влияние интенсивности сельскохозяйственной нагрузки на сообщества почвенных нематод естественных лугов, сенокосных угодий и агроценозов с монокультурами в условиях Республики Карелия. Разнообразие фауны почвенных нематод естественных и сенокосных лугов было сходным и значимо снижалось в агроценозах. Численность нематод имела наибольшие значения в почве лугов и снижалась по мере усиления интенсивности сельскохозяйственного использования земель. В эколого-трофической структуре сообществ нематод всех типов биоценозов доминировали бактериотрофы с максимальной долей в агроценозах. Относительное обилие хищников и нематод, ассоциированных с растениями, в агроценозах было значительно ниже по сравнению с естественными и сенокосными лугами. Эколого-популяционные индексы сообществ нематод свидетельствовали о стабильной и многокомпонентной почвенной экосистеме луговых биоценозов. Индекс преобладающего пути разложения органического вещества CI показал, что деструкция идет с преимущественным участием бактерий. В агроценозах отмечено снижение значений индексов SI и CI и возрастание индекса EI. Такое соотношение индексов указывает на упрощенную трофическую сеть и нарушенную почвенную экосистему в агроценозах. Дискриминантный анализ показал, что достоверное разделение исследованных биоценозов возможно только между агроценозами и естественными лугами за счет индекса SI. Однако из-за выявленного положительного влияния фактора широтного положения на индекс SI в агроценозах можно предположить, что применение показателя в северных экосистемах имеет некоторые ограничения.
Нематоды (круглые черви) представляют одну из наиболее многочисленных и разнообразных групп в почвенной фауне. Повсеместное распространение, большое количество видов с разнообразными типами питания, тесная трофическая связь с бактериями, грибами, растениями и активное участие в процессах преобразования органического вещества обеспечивают группе высокую значимость в экологических исследованиях при оценке состояния почвенных экосистем [1–3]. Исследования отечественных и зарубежных авторов в последние десятилетия показали, что сообщества почвенных нематод могут применяться в качестве эффективного инструмента диагностики состояния и особенностей функционирования почвенных трофических сетей как в естественных биоценозах [4, 5], так и под воздействием различных видов трансформации среды [6, 7], включая сельскохозяйственные мероприятия [8–13]. Также проводились [14, 15] крупномасштабные нематологические исследования лугов, находящихся в естественном состоянии или использующихся в сельскохозяйственных целях. Большое практическое значение имеет изучение реакций сообществ почвенных нематод в агроценозах на внесение удобрений [16], выращивание монокультур [8, 17], различные типы землепользования [11, 18 и др.].
За три десятилетия существования эколого-популяционных индексов, разработанных на основе анализа фауны нематод, стали очевидными и некоторые ограничения в их использовании и интерпретации [19]. Отмечается, что применение индексов следует проводить с осторожностью в ходе первичной сукцессии [20], в экстремальных условиях тундровых экосистем [21] и полярных пустынь [22], в ряде случаев при оценке состояния почвенных агроэкосистем под влиянием землепользования разной интенсивности [23]. Ограничения в большинстве случаев связаны с фрагментарными знаниями об аутэкологии отдельных таксонов нематод [19, 24]. Следует обратить внимание на то, что индексы могут некорректно отражать степень нарушения и состояние почвенной трофической сети вследствие чувствительности к влиянию “нецелевых” факторов. Так, широта местности [25], количество осадков, тип почвы и глубина отбора проб [24, 26] могут оказывать влияние на значения индексов, приводя к несоответствию формируемой ими картины и наблюдаемых процессов [27].
В крупномасштабных экологических исследованиях почвенных нематод сведения о группе, особенностях структуры сообществ и эколого-популяционных индексах на территории Российской Федерации зачастую выглядят как “белое пятно”. Поэтому региональные исследования, позволяющие восполнить этот пробел и повысить доступность сведений для мирового научного сообщества, являются актуальными. Кроме того, отмечалось [23, 24], что для использования нематод в качестве биоиндикаторов в больших географических масштабах необходима калибровка индексов относительно экосистемы, климата и типа почвы. Таким образом, крупные региональные исследования, охватывающие значительный диапазон биоценозов, позволяют дополнительно протестировать эффективность концепции использования нематод в биоиндикации состояния почвенных экосистем и выявить возможные региональные особенности применения биоиндикационного подхода.
Значительный массив данных по сообществам почвенных нематод, накопленных в ходе многолетних исследований на территории Республики Карелия, позволил сформировать ряд биоценозов, различающихся по интенсивности сельскохозяйственного использования – от естественных лугов, не подверженных заметному антропогенному воздействию, к сенокосным лугам, испытывающим сельскохозяйственную нагрузку низкой интенсивности, и к агроценозам с пропашными культурами как пример высокого уровня трансформации. Собранный материал лег в основу анализа, нацеленного на выявление показателей нематод, чувствительных к сельскохозяйственной нагрузке разной интенсивности в условиях северных экосистем. Вытянутая в широтном направлении территория Республики Карелия стала удобным полигоном для оценки широтной изменчивости различных характеристик сообществ почвенных нематод лугов и агроценозов.
В представленной работе мы предполагаем, 1) что различные характеристики сообществ почвенных нематод будут информативны при оценке степени трансформации почвенных экосистем агроценозов в условиях Севера и 2) что широта расположения исследуемых биоценозов может иметь значение для чувствительности нематологических параметров/индексов к антропогенной трансформации.
МАТЕРИАЛ И МЕТОДЫ
Изучение влияния интенсивности сельскохозяйственной нагрузки на сообщества нематод проведено на большом массиве данных по лугам и землям сельскохозяйственного использования различного назначения. Всего на территории Республики Карелия исследовано 56 естественных лугов, 17 сенокосных угодий с частичным использованием под пастбища, 31 агроценоз с монокультурами (картофель и ряд других пропашных культур) (рис. 1). Почвы исследованных луговых биоценозов относятся к дерновым, дерново-подзолистым, торфяно-дерновым и шунгитовым типам. На пашне (агроценозы с монокультурами) почва утрачивает строение, характерное для естественных почв, что позволяет отнести ее к отделу агроземов.
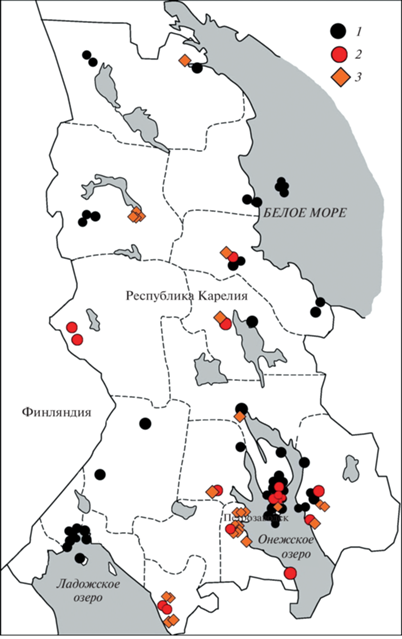
Образцы почвы были отобраны случайным образом из каждого биоценоза в пределах типичного однородного участка растительности (2 × 2 м) с использованием почвенного бура (диаметр 20 мм) на глубину 0–15 см в 9 повторностях. Таким образом, в общей сложности были обследованы 104 пробные площадки и отобрано 936 проб. Нематод выделяли модифицированным методом Бермана с экспозицией 48 ч, фиксация – ТАФом (триэтаноламин : формалин : вода в соотношении 2 : 7 : 91) [28]. Идентификацию нематод осуществляли на врéменных глицериновых препаратах. Устанавливали систематическую принадлежность (до уровня рода) не менее чем 100 особей нематод из каждой пробы с помощью микроскопа при ×400–600. Каждый таксон нематод относили к одной из шести эколого-трофических групп: бактериотрофы (Б), микотрофы (М), политрофы (П), хищники (Х), паразиты растений (Пр) и нематоды, ассоциированные с растением (Аср) [29, 30].
Для оценки состояния сообществ нематод и особенностей функционирования почвенных экосистем использованы следующие параметры: плотность популяций нематод (экз/100 г сырой почвы); таксономическое разнообразие (количество родов), индекс биоразнообразия Шеннона H ' [31]; эколого-трофическая структура сообществ с построением рядов доминирования; эколого-популяционные индексы сообществ нематод, которые включают индекс зрелости сообществ ΣMI [1] и индексы, характеризующие почвенную трофическую сеть – индексы структурирования SI и обогащения EI почвенной трофической сети, индекс преобладающего пути разложения органического вещества в почве CI [3].
Индекс ΣMI рассчитывается на основе состава и соотношения таксонов нематод с различными экологическими особенностями, которые связаны с их морфологией, биологией и экологией и выражены в значениях, присвоенных каждому таксону по c-р-шкале Бонгерса [1]. Колонизаторы с наименьшим значением (c-p = 1) устойчивы к неблагоприятным условиям существования и имеют быстрые темпы размножения. Персистеры c наибольшим значением (c-p = 5) чувствительны к факторам окружающей среды и имеют противоположные колонизаторам характеристики.
Индексы SI, EI и CI разработаны на основе концепции функциональных групп нематод [3], которые в свою очередь определяются объединением трофических групп [29] и с-р-классов [1]. Индекс EI (enrichment index) основан на чувствительности функциональных групп нематод, не связанных с растениями, к возрастанию доступности пищевых ресурсов и характеризует обогащение почвы органикой. Индекс SI (structure index) вычисляется на основе функциональных групп нематод с высокими значениями по с-р-шкале и отражает увеличение числа трофических связей в экосистеме, степень зрелости почвенной трофической сети, сложность и стабильность среды обитания в целом. Фаунистический профиль, построенный при совмещении значений индексов SI и EI, дает возможность охарактеризовать состояние почвенных трофических сетей и почвенной экосистемы в целом [3]. Индекс CI (channel index) показывает преобладающий путь разложения органического вещества в почвенной экосистеме. Низкие значения индекса указывают на активное участие бактерий в разложении органики, высокие – свидетельствуют о преобладании грибов в данном процессе.
Числовые данные представлены в форме М ± SE (среднее значение ± стандартная ошибка среднего). Средние значения рассчитаны для каждой пробной площадки. Статистическая обработка данных проведена с использованием U-критерия Манна-Уитни (Mann-Whitney, U-test). Различия между группами считали достоверными при р < 0.05.
Ординация сообществ почвенных нематод исследованных биоценозов проведена методом неметрического многомерного шкалирования (nMDS) с использованием индекса Брея-Кертиса на основе относительного обилия таксонов нематод. Оценку различий сообществ нематод между группами исследованных биоценозов проводили с помощью анализа группового сходства (ANOSIM). Для классификации исследованных лугов и агроценозов на основе сообществ почвенных нематод применяли дискриминантный анализ. Были сформированы три выборки: луговые биоценозы, сенокосные угодья, агроценозы. Дискриминантные функции рассчитывали на основе трех признаков – значениях индексов SI, EI и CI. Для каждой точки отбора почвенных проб рассчитывали канонические коэффициенты и вероятность отнесения локального сообщества нематод к одному из трех априорно заданных типов биоценозов (луга, сенокосы, агроценозы). Дискриминантный анализ проводили в два этапа: на первом этапе составляли уравнение, разделяющее био- и агроценозы, на втором определяли для локального сообщества нематод наиболее вероятную группу биоценоза.
Для оценки влияния фактора широтного положения биоценоза на относительное обилие трофических групп нематод и эколого-популяционные индексы использовали регрессионный анализ. Все числовые нематологические данные предварительно подвергали логарифмированию для нормализации выборки.
Статистические анализы выполнены при помощи программы PAST 3.19 [32] и программного пакета R [33].
Для выполнения исследования использовано научное оборудование Центра коллективного пользования Федерального исследовательского центра “Карельский научный центр Российской академии наук”.
РЕЗУЛЬТАТЫ
Таксономическое разнообразие, численность, структура сообществ почвенных нематод лугов и агроценозов
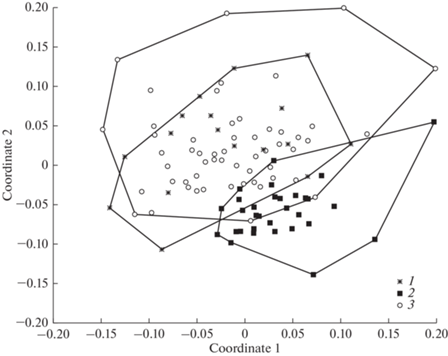
Таксономическое разнообразие почвенных нематод естественных лугов и сенокосных угодий было сходным и значимо уменьшалось в агроценозах (табл. 1). Ординация, проведенная методом неметрического многомерного шкалирования на основе относительного обилия таксонов нематод, продемонстрировала весьма четкое и статистически значимое разделение сообществ почвенных нематод лугов и агроценозов (ANOSIM-тест; R = = 0.302; p = 0.0001), а также сенокосных угодий и агроценозов (R = 0.503; p = 0.0001). Различия сообществ нематод лугов и сенокосных угодий, напротив, минимальны и статистически незначимы (R = 0.08; p = 0.09) (рис. 2).
Таблица 1.
Таксономическое разнообразие и численность нематод в почве лугов, сенокосных угодий и агроценозов Республики Карелия
| Биоценоз | n | Число родов | Плотность популяций, экз/100 г почвы | Н ' | ||
|---|---|---|---|---|---|---|
| среднее | min–max | среднее | min–max | |||
| Луговые биоценозы | 56 | 30.7 ± 1.03а | 13–46 | 2403.6 ± 217.89а | 116–6244 | 3.87 ± 0.08а |
| Сенокосные угодья | 17 | 30.2 ± 1.99а | 12–45 | 1812.0 ± 285.82аb | 87–3824 | 3.70 ± 0.15аb |
| Агроценозы | 31 | 24.3 ± 0.91b | 16–35 | 1387.1 ± 193.16b | 146–4490 | 3.49 ± 0.09b |
Примечание. Здесь и в табл. 2, 3: n – число исследованных биоценозов; значения с различными буквенными обозначениями статистически различаются (p < 0.05); Н ' – индекс разнообразия Шеннона.
Рис. 2.
Ординационная диаграмма nMDS сообществ почвенных нематод на основе относительного обилия таксонов нематод из трех типов биоценозов Республики Карелия: 1 – сенокосные угодья, 2 – агроценозы, 3 – луга.
Плотность популяций нематод имела самые высокие значения в лугах и самые низкие – в агроценозах; промежуточное значение показателя было отмечено в почве сенокосных угодий. Естественные луга и агроценозы достоверно отличались по всем параметрам (см. табл. 1).
В эколого-трофической структуре сообществ нематод исследованных биоценозов доминирующей группой были бактериотрофы с максимальной долей в агроценозах (табл. 2). В естественных лугах субдоминантами выступали микотрофы, три трофические группы (П, Пр, Аср) имели почти одинаковый вклад в фауну нематод. В условиях вмешательства человека отмечена тенденция увеличения вклада паразитов растений в сообщества почвенных нематод, однако различия статистически незначимы. В сенокосных угодьях доля группы Пр была близка к микотрофам, в агроценозах они выступали субдоминантами в ряду доминирования трофических групп. Обратная тенденция – постепенное снижение доли в сообществе при увеличении интенсивности сельскохозяйственной нагрузки – характерна для политрофов. В агроценозах доля двух трофических групп (Х и Аср) была значительно ниже по сравнению с естественными и сенокосными лугами (см. табл. 2).
Таблица 2.
Эколого-трофическая структура сообществ почвенных нематод луговых биоценозов и сельскохозяйственных земель Республики Карелия с различной интенсивностью использования
| Трофические группы | Луговые биоценозы (n = 56) |
Сенокосные угодья (n = 17) |
Агроценозы (n = 31) |
|---|---|---|---|
| Б | 49.6 ± 2.24а | 48.0 ± 4.70а | 60.7 ± 2.68b |
| М | 12.3 ± 0.90а | 14.4 ± 2.74а | 11.5 ± 1.52а |
| П | 11.4 ± 1.16а | 10.1 ± 1.81аb | 7.8 ± 1.43b |
| Х | 5.3 ± 1.47а | 4.8 ± 1.45а | 1.2 ± 0.35b |
| Аср | 10.4 ± 1.39а | 8.5 ± 1.40а | 2.8 ± 0.57b |
| Пр | 11.1 ± 1.52а | 14.2 ± 3.86а | 16.0 ± 2.89а |
| Ряды доминирования трофических групп | |||
| Б → М → П ≈ Пр → Аср → Х | Б → М ≈ Пр → П → Аср → Х | Б → Пр → М → П → Аср → Х | |
Эколого-популяционные индексы сообществ почвенных нематод лугов и агроценозов
Установлено, что по большинству эколого-популяционных индексов сообщества нематод ненарушенных лугов и сенокосных угодий были сходны и значимо различались с агроценозами. Луга и сенокосные угодья характеризовались высокими значениями индекса зрелости ΣMI и структурирования SI, умеренным уровнем обогащения почв (EI < 50). Индекс CI показал, что деструкция органического вещества почвы идет с преимущественным участием бактерий. Использование земель для выращивания монокультур привело к снижению значений индексов SI и CI. Индекс EI, напротив, возрастал, что связано с обогащением почвы органикой (табл. 3).
Таблица 3.
Эколого-популяционные индексы, рассчитанные для сообществ почвенных нематод луговых биоценозов и сельскохозяйственных земель Республики Карелия с различной интенсивностью использования
| Биоценоз | n | ΣMI | SI | EI | CI |
|---|---|---|---|---|---|
| Луговые биоценозы | 56 | 2.61 ± 0.02а | 70.62 ± 1.74а | 47.78 ± 2.59а | 25.26 ± 2.73аb |
| Сенокосные угодья | 17 | 2.66 ± 0.06а | 61.43 ± 5.66a | 39.96 ± 3.87а | 27.98 ± 5.45а |
| Агроценозы | 31 | 2.42 ± 0.03b | 45.88 ± 3.35b | 57.03 ± 2.83b | 17.67 ± 2.67b |
Оценка состояния и степени трансформации почвенной экосистемы на основе фаунистического профиля сообществ нематод
Состояние трофических сетей и степень нарушенности почвенных экосистем были оценены с помощью фаунистического профиля сообществ нематод, полученного на основе эколого-популяционных индексов SI и EI и представленного графически в двумерном пространстве.
Если индекс структурирования SI имел высокие значения (луговые биотопы), то биоценозы попадали в квадраты В или С профиля, что указывает на зрелую и структурированную трофическую сеть (рис. 3). Большинство агроценозов (более 60%) располагались в квадратах А и D профиля, что определяется низкими значениями SI. Индекс обогащения EI, характеризующий количество доступной органики в почве, сильно варьировал (в среднем от 30 до 80). Выявленное соотношение индексов для большинства агроценозов указывает на упрощение трофических сетей. На графике фаунистического профиля выделялась группа агроценозов с высокими (>50) значениями индекса SI: они, как и луга, находились в квадратах В и С, несмотря на трансформацию почвенного покрова (см. рис. 3).
Рис. 3.
Фаунистический профиль, характеризующий почвенную трофическую сеть сенокосных угодий (1), агроценозов (2) и луговых биоценозов (3) Республики Карелия: А – слаборазвитые почвенные экосистемы с высоким уровнем нарушенности (disturbed); В – зрелые экосистемы с низким или средним уровнем нарушенности (maturing); С – структурированные ненарушенные экосистемы (structured); D – деградированные почвенные экосистемы, находящиеся в условиях стресса (degraded) [3].
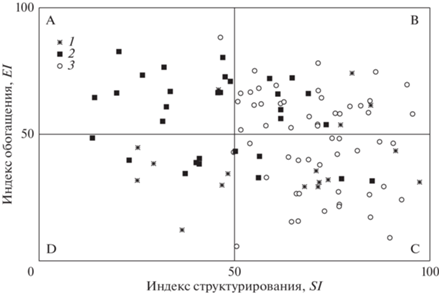
Сенокосные угодья занимали промежуточное положение: одна их часть характеризовалась как стабильные мало- или ненарушенные экосистемы (10 точек из 17), имея сходство с лугами (квадраты В и С), другая часть (6 точек) – как находящиеся в условиях стресса с деградированной трофической сетью (квадрат D профиля) (см. рис. 3).
Применение регрессионного анализа для оценки влияния фактора широтного положения лугов и агроценозов на различные характеристики сообществ почвенных нематод
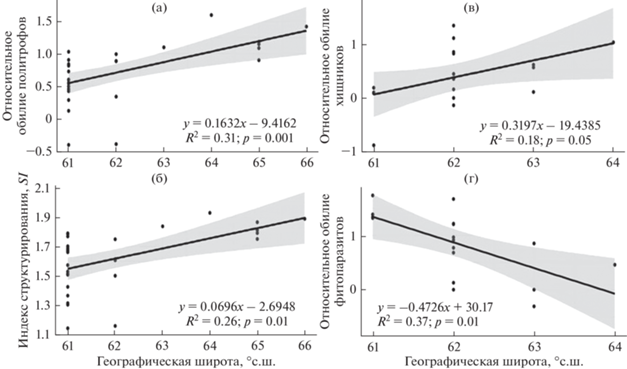
Проведенный регрессионный анализ показал статистически значимое влияние фактора широты расположения биоценоза на обилие нематод-политрофов и индекс структурирования SI в агроценозах (рис. 4а, 4б). При этом и политрофы, и индекс SI в агроценозах прямо пропорциональны широте. Для сенокосных угодий статистически значимым является влияние широты на обилие паразитов растений, которое носит отрицательный характер, и на хищников – с их положительной реакцией на фактор (рис. 4в, 4г). Согласно коэффициентам детерминации (R2), исследованный фактор достаточно хорошо описывает изменчивость вышеописанных характеристик сообществ почвенных нематод. Для остальных нематологических параметров в сенокосных лугах и агроценозах достоверных связей с данным фактором не установлено, также не выявлены значимые связи и в естественных лугах.
Рис. 4.
Зависимость некоторых нематологических параметров в агроценозах и сенокосных угодьях от географической широты. Приведены только статистически достоверные связи фактора с нематологическими параметрами: а – обилие нематод-политрофов, б – индекс структурирования, в – обилие хищников, г – обилие паразитов растений; R2 – коэффициент детерминации; p – уровень значимости полученной модели. Ось ординат логарифмирована.
Применение дискриминантного анализа для классификации исследованных биоценозов на основе эколого-популяционных индексов
С помощью дискриминантного анализа определена функциональная роль эколого-популяционных индексов сообществ почвенных нематод для классификации биоценозов с различной степенью трансформации. Для проверки качества дискриминации использовано значение лямбды Уилкса (Wilks lambda, λ), величина которой меняется от 1 (нет дискриминации) до 0 (полная дискриминация). Установлено, что наибольшую функциональную нагрузку несет первая дискриминантная функция (Wilks λ = 0.705193, p < 0.0001), разделяющая биотопы на основе SI, рассчитанного для локальных сообществ нематод. Вторая и третья функции (CI, EI), используемые в анализе, незначимы.
Общая успешность классификации биоценозов, полученной с помощью дискриминантной функции на основе индексов сообществ нематод, оценивается в 52.88% (табл. 4). Среди сенокосных угодий не отмечено ни одного “соответствия” заданной группе. Следовательно, данные биоценозы не проявляют выраженных особенностей и по значениям индекса SI имеют сходство как с агроценозами (за счет низких значений в ряде биоценозов), так и с лугами (по высоким значениям). Для лугов и агроценозов доля соответствия заданной группе была на уровне 61–64%.
Таблица 4.
Классификация разнотипных биоценозов Республики Карелия на основе эколого-популяционных индексов сообществ нематод с использованием дискриминантного анализа
| Биоценоз | Число биоценозов | Прогнозируемый тип биоценоза (доля соответствия, %)* | ||
|---|---|---|---|---|
| луговые биоценозы | сенокосные угодья | агроценозы | ||
| Луговые биоценозы | 56 | 36 (64.29%) | 15 (26.79%) | 5 (8.93%) |
| Сенокосные угодья | 17 | 10 (58.82%) | 0 (0.00%) | 7 (41.18%) |
| Агроценозы | 31 | 4 (12.90%) | 8 (25.81%) | 19 (61.29%) |
Таким образом, дискриминантный анализ показал, что на основе индекса SI, рассчитанного для сообществ почвенных нематод, можно достоверно классифицировать луга и агроценозы. Индексы EI и CI вносят вклад в разделение биоценозов с различной интенсивностью антропогенного воздействия, однако роль данных дискриминантных функций незначима.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Анализ численности и разнообразия почвенных нематод лугов и агроценозов в сравнительном аспекте показал, что агроценозы выделяются как наименьшим числом выявленных родов, так и низкой общей численностью нематод. Полученные нами результаты выявили снижение числа родов (–21%) и численности нематод (–42%) в агроценозах по сравнению с лугами на уровне, сходном с закономерностями, полученными другими авторами как в региональном (Чехия; число родов: –56%, численность: –46%), так и глобальном (–32 и –27% соответственно) масштабах [25, 34]. Известно [5], что луга среди различных типов естественных биоценозов выделяются высоким разнообразием фауны почвенных нематод, уступающим на территории европейской части России только широколиственным лесам и степям Центрально-Черноземного района. Согласно собственным данным [10] и сведениям других исследователей [11, 12, 34, 35], под влиянием сельскохозяйственной нагрузки в пахотных почвах отмечается снижение разнообразия нематодофауны. С другой стороны, сенокошение как форма управления луговыми угодьями не оказывает влияния на разнообразие нематод и индекс Шеннона [18].
Перестройка эколого-трофической структуры сообществ нематод под влиянием различных нарушений среды, как правило, связана с увеличением доли бактериотрофов и фитопаразитов и уменьшением доли хищников и политрофов [35]. В почве всех типов исследованных биоценозов нами выявлено преобладание бактериотрофов с максимумом в агроценозах, что было показано и в других работах [9, 11, 12, 34]. Увеличение численности нематод-паразитов растений отмечалось в результате чрезмерного внесения минеральных удобрений, интенсификации аграрных мероприятий, при долгосрочном выращивании монокультуры [6]. По нашим данным, в условиях вмешательства человека наблюдалась тенденция увеличения вклада паразитов растений в сообщества нематод, однако различия статистически незначимы.
В агроценозах относительное обилие хищников и нематод, ассоциированных с растениями, было значительно меньше по сравнению с естественными и сенокосными лугами. Изменение их вклада в сообщество нематод обусловлено увеличением интенсивности сельскохозяйственной нагрузки. Известно, что присутствие хищных нематод, относящихся к персистерам или К-стратегам, является показателем стабильности сообществ почвенных нематод, и он снижается в условиях трансформации среды обитания [1]. По литературным данным [34] в условиях Центральной Европы численность нематод, ассоциированных с растениями, ниже в почве интенсивно используемых сельскохозяйственных полей по сравнению с разнотравным лугом. В данном и ранее проведенном нами исследовании [10] были получены сходные результаты – численность нематод этой группы значительно снижалась в почве агроценозов.
Агроценозы обычно характеризуются низкими значениями индекса преобладающего пути разложения органического вещества CI [10, 12], однако отмечены и отклонения от данной закономерности. Так, высокие (>50) значения индекса CI выявлены в почве действующих сельскохозяйственных полей [34], преобладание грибного пути в разложении органики отмечено и под влиянием сенокошения [18]. Полученные нами минимальные величины индекса CI в агроценозах подтверждают, что в данных почвах бактериальный компонент играет большую роль в процессе деструкции органики. Это может быть связано с использованием различных агротехнических приемов, применяемых для выращивания пропашных культур (внесение удобрений, вспашка, борьба с сорняками и т.п.). Значение микодеструкции (возрастание индекса CI) лишь незначительно возрастает в лугах и сенокосных угодьях.
Известно, что агроценозы с монокультурами обычно характеризуются низкими значениями индексов MI, SI и высокими индекса EI [12, 34]. Графическое представление в двумерном пространстве условий почвенной трофической сети на основе этих индексов позволило наглядно показать, что большинство агроценозов характеризуются упрощенными трофическими сетями и являются нарушенными почвенными экосистемами, которые испытывают воздействие агротехнических приемов (вспашка и внесение органики) или других неблагоприятных факторов среды (климатических или антропогенных) [3]. Под влиянием сенокошения индексы SI и MI также снижаются, но лишь незначительно и только в условиях высокой частоты скашивания [13]. Результаты нашего исследования показали, что в почве сенокосных угодий эколого-популяционные индексы сообществ нематод не снижались по сравнению с луговыми биотопами. Последние являются стабильными местообитаниями со сложными многокомпонентными трофическими сетями, на что указывают высокие значения SI.
Однако следует отметить, что среди агроценозов выделялась группа, которая также характеризовалась высокими значениями индекса SI. По соотношению индексов EI и SI они, как и луга, попадали в квадраты В и С фаунистического профиля, несмотря на трансформацию почвенного покрова, связанную с ежегодной обработкой пахотного слоя почвы, внесением удобрений и выращиванием монокультур. Детальный анализ особенностей данных местообитаний (специфики биотопов) позволил выявить, что значения индексов обусловлены их географическим положением (большинство нетипичных агроценозов находятся в северных районах Республики Карелия, выше 63° с.ш.). Высокие значения SI в этих точках обусловлены увеличением в сообществах доли нематод-политрофов с высокими значениями по с-р-шкале Бонгерса.
Проведенный регрессионный анализ подтвердил статистически значимое положительное влияние фактора широтного положения на обилие политрофов и индекс SI в агроценозах. По нашим данным, в агроценозах Севера (63–66° с.ш.) политрофы занимают вторую–третью позиции в ряду доминирования эколого-трофических групп, тогда как в центральных и южных районах Карелии (61–62° с.ш.) их доля в фауне заметно снижается [8]. Отмечается, что некоторые таксоны нематод, если рассматривать их трофическую или c-p-группы, могут вести себя или реагировать иначе, чем ожидалось, или быть неожиданно многочисленными в определенных условиях [19]. Например, политрофы, являющиеся персистерами, чувствительными к нарушениям среды, могут ввести себя как колонизаторы [20], положительно реагировать на обогащение почвы органикой [36] и демонстрировать высокую численность в экстремальных условиях [22, 37]. Кроме того, численность политрофов и хищников имела значимую сильную положительную связь с широтой в глобальном масштабе [25] и была наибольшей в северных тундровых экосистемах [15]. По нашим данным, наряду с политрофами в агроценозах хищники также продемонстрировали сходную широтную изменчивость в сенокосных угодьях. Сообщалось [37], что некоторые виды нематод-политрофов in vitro питались другими нематодами, проявляя хищническое поведение, инцистированными амебами, гифами грибов, водорослями и даже бактериальными колониями. Можно предположить, что именно всеядность является условием успешной адаптации к неблагоприятным условиям среды.
Результаты дискриминантного анализа показали, что достоверно классифицировать луга и агроценозы можно на основе индекса SI, а роль индексов EI и CI в разделении сообществ незначима. Полученные нами результаты подтверждают литературные данные [7]: при изучении различных типов естественных и нарушенных местообитаний показано, что именно индексы SI и MI являются наиболее чувствительными для выявления различий между местообитаниями. С другой стороны, индекс SI, наиболее чувствительный к антропогенной трансформации экосистем, продемонстрировал в агроценозах и широтную изменчивость с максимальными значениями в северных экосистемах. Поэтому, применяя показатель для оценки состояния почвенных экосистем агроценозов в условиях Севера, интерпретировать полученные результаты следует осторожно.
Сенокосные угодья, характеризующиеся низкой интенсивностью сельскохозяйственного использования, имели высокое сходство с естественными лугами и по большинству анализируемых показателей достоверно с ними не различались. Ранее также сообщалось [23] о низкой чувствительности индексов для агроэкосистем под влиянием разной интенсивности землепользования.
ЗАКЛЮЧЕНИЕ
В результате нашего исследования установлены изменения разнообразия фауны, численности и структуры сообществ почвенных нематод в условиях сельскохозяйственного использования земель. Среди рассмотренных параметров наиболее чувствительными к высокой интенсивности сельскохозяйственной нагрузки оказались количество таксонов нематод и численность, процентный вклад бактериотрофов и нематод-К-стратегов (политрофов и хищников) в структуру сообщества. В агроценозах по сравнению с лугами отмечена тенденция увеличения доли паразитов растений, однако различия статистически незначимы.
Сельскохозяйственные мероприятия влияли и на значения эколого-популяционных индексов. Индексы, отражающие процессы деструкции органики и уровень обогащения почвы (CI и EI), вносят определенный вклад в разделение биоценозов с различной интенсивностью антропогенного воздействия. Так, агроценозы, в большей степени, чем другие биотопы, характеризуются высокими значениями индекса EI и низкими индекса CI. Однако с использованием дискриминантного анализа установлено, что только на основе индекса структурирования SI, несущего значимую функциональную нагрузку, можно достоверно классифицировать луга и агроценозы. С другой стороны, индекс SI, наиболее чувствительный к антропогенной трансформации экосистем, продемонстрировал и широтную изменчивость в агроценозах с максимальными значениями в северных экосистемах. Поэтому применение показателя для оценки состояния почвенных экосистем агроценозов в условиях Севера следует проводить осторожно и аккуратно интерпретировать полученные результаты.
Сенокосные угодья, характеризующиеся низкой интенсивностью сельскохозяйственного использования, имели высокое сходство с естественными лугами и по большинству анализируемых показателей достоверно с ними не различались. Можно констатировать, что индексы и другие нематологические параметры имеют низкую эффективность в небольшом градиенте воздействия и на начальных этапах нарушений.
Таким образом, в результате исследования выявлены показатели, чувствительные к высокой интенсивности сельскохозяйственной нагрузки и информативные при биоиндикационной оценке состояния и степени трансформации почвенных экосистем агроценозов. Установлено статистически значимое влияние фактора широты расположения биоценоза на различные характеристики сообществ нематод, которое может приводить к ряду ограничений в использовании нематологических параметров/индексов при антропогенной трансформации.
Исследование выполнено при финансовой поддержке средств федерального бюджета на выполнение государственного задания ИБ КарНЦ РАН (тема № 122032100130-3). Авторы выражают признательность д.б.н., профессору Е.П. Иешко за консультацию и обсуждение результатов.
Авторы заявляют об отсутствии конфликта интересов.
Авторы подтверждают, что в работе с животными соблюдались применимые этические нормы.
Список литературы
Bongers T. The maturity index: an ecological measure of environmental disturbance based on nematode species composition // Oecologia. 1990. V. 83. P. 14–19.
Bongers T., Ferris H. Nematode community structure as a bioindicator in environmental monitoring // Trends in Ecology & Evolution. 1999. V. 14. № 6. P. 224–228.
Ferris H., Bongers T., de Goede R.G.M. A framework for soil food web diagnostics: extension of the nematode faunal analysis concept // Applied Soil Ecology. 2001. V. 18. P. 13–29.
Матвеева Е.М., Сущук А.А. Особенности сообществ почвенных нематод в различных типах естественных биоценозов: информативность параметров оценки // Изв. РАН. Сер. биологич. 2016. № 5. С. 551–560. [Matveeva E.M., Sushchuk A.A. Features of soil nematode communities in various types of natural biocenoses: effectiveness of assessment parameters // Biology Bulletin. 2016. V. 43. № 5. P. 474–482. https://doi.org/10.1134/S1062359016040099]10.1134/S1062359016040099]https://doi.org/10.7868/S0002332916040093
Sushchuk A.A., Matveeva E.M., Kalinkina D.S. Soil nematode communities of the European part of Russia: latitudinal aspect // Russ. J. of Nematology. 2021. V. 29. № 2. P. 200. https://doi.org/10.24412/0869-6918-2021-2-186-203
Wasilewska L. Soil invertebrates as bioindicators, with special reference to soil-inhabiting nematodes // Russ. J. of Nematology. 1997. V. 5. № 2. P. 113–126.
Briar S.S., Jagdale G.B., Cheng Z. et al. Indicative value of soil nematode food web indices and trophic group abundance in differentiating habitats with a gradient of anthropogenic impact // Environmental Bioindicators. 2007. V. 2. Iss. 3. P. 146–160. https://doi.org/10.1080/15555270701590909
Груздева Л.И., Матвеева Е.М. Расширение ареала картофельной цистообразующей нематоды на Северо-Западе России // Труды Центра паразитологии. Т. XLVI: Биоразнообразие и экология паразитов. М.: Наука, 2010. С. 71–80.
Мигунова В.Д., Кураков А.В. Структура микробной биомассы и трофические группы нематод в дерново-подзолистых почвах постагрогенной сукцессии в южной тайге (Тверская область) // Почвоведение. 2014. № 5. С. 584–589. [Migunova V.D., Kurakov A.V. Structure of the microbial biomass and trophic groups of nematodes in soddy-podzolic soils of a postagrogenic succession in the southern taiga (Tver oblast) // Eurasian Soil Science. 2014. V. 47. № 5. P. 453–458. https://doi.org/10.1134/S1064229314050160]https://doi.org/10.7868/S0032180X14050165
Матвеева Е.М., Сущук А.А., Калинкина Д.С. Сообщества почвенных нематод агроценозов с монокультурами (на примере Республики Карелия) // Труды Карельского научного центра РАН. Сер.: Экологические исследования. 2015. № 2. С. 16–32. https://doi.org/10.17076/eco16
Freckman D.W., Ettema C.H. Assessing nematode communities in agroecosystems of varying human intervention // Agriculture, Ecosystems and Environment. 1993. V. 45. P. 239–261.
Háněl L. An outline of soil nematode succession on abandoned fields in South Bohemia // Applied Soil Ecology. 2010. V. 46. P. 355–371.
Pan F.J., Yang L.Y., Wang C.L. et al. Effects of mowing frequency on abundance, genus diversity and community traits of soil nematodes in a meadow steppe in northeast China // Plant and Soil. 2022. V. 473. Iss. 1–2. P. 89–107. https://doi.org/10.1007/s11104-020-04740-9
De Goede R.G.M., Bongers T. (eds.). Nematode communities of northern temperate grassland ecosystems. Giessen: Focus Verlag, 1998. 338 p.
van den Hoogen J., Geisen S., Routh D. et al. Soil nematode abundance and functional group composition at a global scale // Nature. 2019. V. 572. P. 194–198.
Gruzdeva L.I., Matveeva E.M., Kovalenko T.E. Changes in soil nematode communities under the impact of fertilizers // Eurasian Soil Science. 2007. V. 40. № 6. P. 681–693.
van Eekeren N., Bos M., de Wit J. et al. Effect of individual grass species and grass species mixtures on soil quality as related to root biomass and grass yield // Applied Soil Ecology. 2010. V. 45. P. 275–283.
Li Y., Liang S., Du X. et al. Mowing did not mitigate the negative effects of nitrogen deposition on soil nematode community in a temperate steppe // Soil Ecology Letters. 2021. V. 3. Iss. 2. P. 125–133. https://doi.org/10.1007/s42832-020-0048-0
Du Preez G., Daneel M., De Goede R. et al. Nematode-based indices in soil ecology: Application, utility, and future directions // Soil Biology and Biochemistry. 2022. V. 169. P. 108640. https://doi.org/10.1016/j.soilbio.2022.108640
Austin E., Semmens K., Parsons Ch., Treonis A. Granite rock outcrops: an extreme environment for soil nematodes? // Journal of Nematology. 2009. V. 41. № 1. P. 84–91.
Кудрин А.А., Конакова Т.Н., Таскаева А.А. Сообщества почвенных нематод различных тундровых фитоценозов, отличающихся степенью развития кустарникового яруса // Экология. 2019. № 6. С. 419–428. [Kudrin A.A., Konakova T.N., Taskaeva A.A. Communities of soil nematodes of various tundra phytocenoses differing in the development level of the shrub layer // Russ. J. of Ecology. 2019. V. 50. № 6. P. 526–534. https://doi.org/10.1134/S1067413619060092]https://doi.org/10.1134/S036705971906009X
Peneva V., Lazarova S., Elshishka M. et al. Nematode assemblages of hair-grass (Deschampsia spp.) microhabitats from polar and alpine deserts in the Arctic and Antarctic // Species and Communities in Extreme Environment. Sofia – Moscow: Pensoft Publishers & KMK Scientific Press, 2009. P. 419–438.
Vonk J.A., Breure A.M., Mulder C. Environmentally-driven dissimilarity of trait-based indices of nematodes under different agricultural management and soil types // Agriculture, Ecosystems and Environment. 2013. V. 179. P. 133–138. https://doi.org/10.1016/j.agee.2013.08.007
Neher D.A. Role of nematodes in soil health and their use as indicators // Russ. J. of Nematology. 2001. V. 33. P. 161–168.
Song D., Pan K., Tariq A. et al. Large-scale patterns of distribution and diversity of terrestrial nematodes // Applied Soil Ecology. 2017. V. 114. P. 161–169. https://doi.org/10.1016/j.apsoil.2017.02.013
Puissant J., Villenave C., Chauvin C. et al. Quantification of the global impact of agricultural practices on soil nematodes: a meta-analysis // Soil Biology and Biochemistry. 2021. V. 161. P. 108383. https://doi.org/10.1016/j.soilbio.2021.108383
Кудрин А.А., Сущук А.А. Методы исследования сообществ почвенных нематод // Russ. J. of Ecosystem Ecology. 2022. V. 7 (2). https://doi.org/10.21685/2500-0578-2022-2-5
van Bezooijen J. Methods and techniques for nematology. Wageningen: Wageningen University Press, 2006. 112 p.
Yeates G.W., Bongers T., de Goede R.G.M. et al. Feeding habits in soil nematode families and genera: An outlain for soil ecologists // Journal of Nematology. 1993. V. 25. № 3. P. 315–331.
Yeates G.W., Wardle D.A., Watson R.N. Relationships between nematodes, soil microbial biomass and weed-management strategies in maize and asparagus cropping systems // Soil biology and biochemistry. 1993. V. 25. № 7. P. 869–876.
Одум Ю. Основы экологии. Пер. с англ. М.: Мир, 1975. С. 186–187.
Hammer Ø., Harper D.A.T., Ryan P.D. Past: paleontological statistics software package for education and data analysis // Paleontological Electronica. 2001. V. 4 (1). P. 1–9. http://palaeo-electronica.org/2001_1/ past/issue1_01.htm
R Core Team. R: A language and environment for statistical computing. R Foundation for Statistical Computing [caйт]. – Vienna, Austria, 2020. URL: http://www.r-project.org/index.html
Háněl L. Recovery of soil nematode populations from cropping stress by natural secondary succession to meadow land // Applied Soil Ecology. 2003. V. 22. P. 255–270.
Wasilewska L. Impact of human activities on nematode communities in terrestrial ecosystems // Ecology of Arable Land. Eds. Clarholm M., Bergstrom L. Dordrecht: Kluwer Academic Publishers, 1989. P. 123–132.
Zhao J., Neher D.A. Soil nematode genera that predict specific types of disturbance // Applied Soil Ecology. 2013. V. 64. P. 135–141. https://doi.org/10.1016/j.apsoil.2012.11.008
Muschiol D., Traunspurger W. Life at the extreme: meiofauna from three unexplored lakes in the caldera of the Cerro Azul volcano, Galápagos Islands, Ecuador // Aquatic Ecology. 2009. V. 43. P. 235–248. https://doi.org/10.1007/s10452-008-9202-y
Дополнительные материалы отсутствуют.