

Экология, 2023, № 4, стр. 261-270
Уровни вторжения чужеродных растений в местообитаниях разных типов на Среднем Урале
Д. В. Веселкин a, Д. И. Дубровин a, А. А. Коржиневская a, Е. Н. Подгаевская a, Л. А. Пустовалова a, Т. Г. Ивченко b, Н. В. Золотарева a, *
a Институт экологии растений и животных УрО РАН
620144 Екатеринбург, ул. 8 Марта, 202, Россия
b Ботанический институт им. В.Л. Комарова РАН
197022 Санкт-Петербург, ул. Профессора Попова, 2, Россия
* E-mail: nvp@ipae.uran.ru
Поступила в редакцию 03.03.2023
После доработки 29.03.2023
Принята к публикации 31.03.2023
- EDN: RSVXTH
- DOI: 10.31857/S0367059723040121
Аннотация
Целью работы было оценить уровни вторжения чужеродных растений в разных естественных и антропогенных местообитаниях Среднего Урала при двух вариантах проявления урбанизации: в черте крупного города и на пригородных территориях. Проанализировано 965 геоботанических описаний, выполненных в естественных (болотных, пойменных, петрофитных, луговых, лесных) и антропогенных (пустыри, скверы, селитебные территории) местообитаниях. Естественные местообитания обследованы в черте крупного города и его пригородной зоне (на расстояниях 30–40 км от города). Характеристиками уровней вторжения были число и доля чужеродных (адвентивных) видов в описании. И в пригородных, и в городских условиях меньше всего чужеродных видов было в болотных местообитаниях (в среднем 0–0.6 вида в описании), а больше всего – в антропогенных местообитаниях (в среднем 9.8–17.6 видов). За городом средние значения числа чужеродных видов в естественных местообитаниях (исключая болота) варьировали в диапазоне 1.0–2.8 вида в описании, в городе – в диапазоне 3.8–6.2 вида. Средние доли чужеродных видов в естественных местообитаниях (исключая болота) за городом составили 1.8–5.6%, в городе – 8.1–16.2%, а в антропогенных местообитаниях – 22.8–32.8%. Пригородные и городские пустыри не различались по числу (10.3 и 12.9 видов в описании) и долям (25.3 и 30.2%) чужеродных видов. Установленные на пригородных территориях уровни вторжения сравнимы с уровнями, указываемыми для Европы.
Из основных этапов внедрения чужеродных растений в аборигенные сообщества во вторичных ареалах – занос, колонизация, натурализация (приживание), широкое распространение – первые три этапа преимущественно зависят от свойств чужеродных видов, а последний в том числе от свойств местообитаний, в которые происходит внедрение.
Свойство местообитаний или сообществ быть подверженными или устойчивыми к вторжению чужеродных растений обозначается как инвазибельность [1–3]. Количественной характеристикой инвазибельности обычно считают число [4–7] или долю [6–9] чужеродных видов в конкретном описании, местообитании или сообществе. Иногда признаки “число/доля чужеродных видов” относят к характеристике “уровень вторжения” (“invasion level”, “level of invasion”) [5, 8–12], а инвазибельность понимается как характеристика относительного или потенциального участия чужеродных растений в данном типе местообитаний или сообществ при выровненных внешних условиях (урбанизации, удаленности от человеческого жилья и мест распространения пропагул чужеродных растений, нарушенности и т.п.) [3]. Поэтому “уровни инвазибельности” рассчитываются математически более сложно, чем уровни вторжения [2, 3, 8, 13–15]. Например, инвазибельность может определяться на основе оценок не только числа, но и обилия или встречаемости чужеродных видов [2, 13–15].
Если уровень вторжения охарактеризован долей чужеродных растений, он идентичен показателю “индекс адвентизации” [16], использовавшемуся при анализе ситуаций и процессов на охраняемых [17–19] и иных [20] территориях.
Уровни вторжения и инвазибельности в масштабе сообществ в отличие от многочисленных оценок адвентизации флор и ценофлор, в отношении которых есть сотни публикаций, на пространстве российской части Евразии оценены фрагментарно [2, 15, 17–24]. По аналогии с показателем доли чужеродных растений во флорах [25–28] можно ожидать высокой географической изменчивости уровней вторжения и инвазибельности. Учитывая связь инвазибельности с антропогенными нарушениями [8, 9, 29, 30] и фрагментацией [31–33], можно также ожидать высокой экологической изменчивости уровней вторжения и инвазибельности.
Уровни трансформации флор вследствие вторжения чужеродных растений изменяются в российской части Евразии в диапазоне от 1.5–22% (флоры заповедников: [25]) до 14–50% (урбанофлоры: [27, 28]) и 40–50% (сегетальные флоры: [26]). Уровни инвазибельности в масштабе местообитаний и сообществ из-за предполагаемой региональной и экологической специфики, а также из-за недостаточности имеющихся сведений прогнозировать сложно. На частичное заполнение этого пробела направлены наши усилия.
Цель работы – охарактеризовать уровни вторжения чужеродных растений в разных естественных и антропогенных местообитаниях Среднего Урала при двух вариантах проявления урбанизации: в черте крупного города и на пригородных территориях.
МАТЕРИАЛ И МЕТОДЫ
Район. Екатеринбург – крупный город площадью около 1.1 тыс. км2 [34], четвертый по численности населения в России (1.5 млн жителей [35]), находится на границе Европы и Азии (56°51′ с.ш., 60°36′ в.д.) и значительно удален от морей и океанов. Город расположен в восточных предгорьях Среднего Урала, рельеф холмисто-увалистый с высотами 250–300 м. Климат умеренно континентальный, средняя температура января –12.6°C, июля +19.0°C. Среднегодовое количество осадков 450–500 мм; наибольшее их количество приходится на теплую часть года – 300–350 мм. Снежный покров устанавливается в начале ноября, его средняя высота 40–50 см. В регионе обычны резкие колебания температур и формирование погодных аномалий: оттепели зимой и заморозки летом. Согласно ботанико-географическому районированию, г. Екатеринбург расположен в подзоне южной тайги, где преобладают сосновые с лиственницей травяные, травяно-кустарничковые и зеленомошные леса на дерново-подзолистых почвах и буроземах [36].
Территория г. Екатеринбурга сильно загрязнена из-за большого числа промышленных предприятий и высокой плотности автотранспортной сети [37–39], а растительность города испытывает сильное рекреационное воздействие [40]. Хорошо выделяются сильно преобразованные центральные и менее преобразованные периферийные районы. Районы города, окружающие промышленные предприятия и застроенные жилыми кварталами, практически лишены естественной растительности. Лесопарки и городские леса – менее преобразованные территории, представленные преимущественно сосновыми лесами.
Город Екатеринбург – центр крупной городской агломерации площадью около 13 тыс. км2 с населением около 2.3 млн жителей, включающей города-спутники Верхняя Пышма, Берёзовский, Дегтярск, Первоуральск, Ревда, Среднеуральск, Арамиль, Сысерть, Полевской и другие населенные пункты. Это обусловливает антропогенную преобразованность ландшафтов, местообитаний и сообществ в пригородной зоне.
Флора собственно города (в пределах городской черты Екатеринбурга) хорошо изучена [41] и включает много чужеродных видов, но их распределение по группам местообитаний охарактеризовано преимущественно на уровне ценофлор или парциальных флор, т.е. на уровне γ-разнообразия [42, 43]. Уровни вторжения установлены для ограниченного круга типов местообитаний – лесопарков [23, 44], фрагментов насаждений с аборигенными и чужеродными видами деревьев [45].
Местообитания. Уровни вторжения определяли применительно к местообитаниям, а не сообществам, учитывая, что типы местообитаний унифицируются строже и однозначнее, чем типы сообществ. Определение уровней вторжения и уровней инвазибельности для типов местообитаний – стандартный подход, часто реализуемый при анализе больших массивов данных как на уровне сообществ [8, 9], так и на уровне флор [46, 47]. Для обеспечения возможности интеграции наших данных с данными других авторов при выборе и определении широты используемых категорий местообитаний опирались на EUNIS habitat classification [48], понимая условность такого подхода по отношению к территориям в азиатской части Евразии.
Исследовали следующие 7 типов естественных местообитаний: 1) болотные местообитания – низинные и верховые болота с древесным ярусом и без деревьев (EUNIS X04, D5); 2) пойменные местообитания – в основном пойменные уремные леса, образованные видами рода Salix, Padus, Alnus (EUNIS G1.1); 3) луга – вторичные суходольные луга (EUNIS E2, E3); 4) естественные лиственные леса с умеренным почвенным увлажнением, образованные видами рода Betula (EUNIS G1); 5) искусственные посадки деревьев – в основном Pinus sylvestris (EUNIS G3.4F); 6) естественные хвойные (с Pinus sylvestris) леса с умеренным почвенным увлажнением (EUNIS G3); 7) петрофитные местообитания – петрофитные степи класса Festuco-Brometea (EUNIS E1.2), поляны с синантропной растительностью у подножия скальных останцов (EUNIS E 5.1), сосновые леса на скальных выходах (EUNIS G3.4).
Кроме того, исследовали три типа антропогенных местообитаний, возникающих и существующих в результате целенаправленной человеческой деятельности: 8) пустыри – сильно нарушенные и постоянно нарушаемые местообитания с искусственным или сильно турбированным грунтом и сорной травяной растительностью (EUNIS E5.1, J1); 9) скверы – целенаправленно созданные в рекреационных и декоративных целях пространства с посадками древесных растений и участками травяной растительности (газонами), исключая участки клумбовых посадок (EUNIS X11, X22, X23); 10) селитебные – частично управляемые местообитания во дворах жилых домов, включая газоны, но исключая участки клумбовых посадок (EUNIS I2).
Местообитания 1–8 различались по проявлению урбанизации (рис. 1). Их подбирали внутри городской черты Екатеринбурга (городские или урбанизированные местообитания) и на пригородных участках (неурбанизированные местообитания), расположенных от городской черты на расстояниях не более 30–40 км. Пустыри на пригородных участках выбирали у дорог, а также вблизи населенных пунктов и на их окраинах. Местообитания 9–10 расположены только в черте Екатеринбурга.
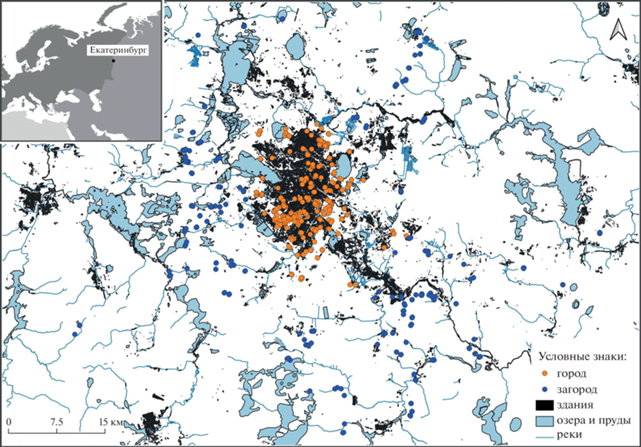
Рис. 1.
Схема расположения местообитаний, в которых выполняли описания, в черте г. Екатеринбурга (оранжевые точки) и на пригородных территориях (синие точки). Каждая точка может маркировать несколько близко расположенных описаний.
Описания. Всего проанализировали 965 геоботанических описаний (табл. 1). Между разными типами местообитаний число описаний различалось почти на порядок, но для каждого типа местообитаний число описаний урбанизированных и неурбанизированных вариантов было близким. Описания выполняли в 2013–2022 гг. на площадях 20 × 20 м в сообществах с древесным (кустарниковым) ярусом и на площадях 10 × 10 м в сообществах без древесного и кустарникового ярусов. Иногда описания были неквадратной формы. Регистрировали видовой состав травяно-кустарничкового, кустарникового и древесного ярусов. Описания начинали выполнять со второй декады июня и по первую декаду августа. Для рандомизации стремились случайно назначать конкретные описываемые сообщества во времени (от начала к концу сезона) и по исполнителям, чтобы один человек не описывал только один тип местообитаний.
Таблица 1.
Число описаний в разных типах местообитаний
| Типы местообитаний |
Уровни урбанизации | Сумма описаний | |
|---|---|---|---|
| неурбанизированные пригородные местообитания | урбанизированные местообитания в черте г. Екатеринбурга | ||
| Болотные | 30 | 28 | 58 |
| Пойменные | 35 | 31 | 66 |
| Луга | 30 | 33 | 63 |
| Лиственные леса | 30 | 28 | 58 |
| Посадки деревьев | 26 | 26 | 52 |
| Хвойные леса | 184 | 226 | 410 |
| Петрофитные | 36 | 6 | 42 |
| Пустыри | 33 | 28 | 61 |
| Скверы | Нет | 55 | 55 |
| Селитебные | Нет | 100 | 100 |
| Всего | 424 | 541 | 965 |
При выполнении описаний фиксировали все виды вегетирующих растений, встреченные на площади 100 или 400 м2. Виды, уверенно идентифицируемые в полевых условиях, регистрировали. Виды, не идентифицируемые в полевых условиях, гербаризировали и определяли в лабораторных условиях.
Аборигенные и чужеродные виды. Все виды растений отнесли к одной из двух категорий – аборигенные или чужеродные. Под чужеродными, или адвентивными, понимали виды, не свойственные местной флоре, занос которых на данную территорию не связан с естественным ходом флорогенеза, а является результатом прямой или косвенной деятельности человека [49]. По времени проникновения в регион – на археофиты и неофиты – адвентивные растения не разделяли. Основной источник, по которому разграничивали аборигенные и чужеродные виды, – конспект флоры Свердловской области [50–57].
Анализ данных. Различия средних значений числа и долей чужеродных видов в описаниях между разными типами (группами) местообитаний анализировали с помощью однофакторного дисперсионного анализа (ANOVA). Попарные различия между вариантами оценивали с помощью критерия Тьюки. В тексте приведены размахи между средними значениями в разных типах местообитаний. Другие обозначения: CV – коэффициент вариации; CI – доверительный интервал.
РЕЗУЛЬТАТЫ
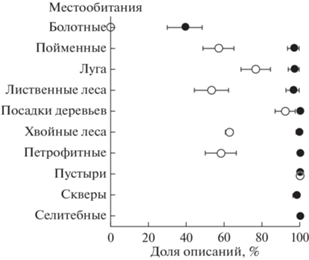
Встречаемость чужеродных видов в описании. Чужеродные растения полностью отсутствовали только на пригородных болотах (рис. 2). Во всех других типах местообитаний, как в городе, так и за городом, преобладали описания, в которых был зарегистрирован хотя бы один чужеродный вид. Вне города хотя бы один чужеродный вид встретился в 53–77% описаний в пойменных, луговых, лесных и петрофитных местообитаниях, в 92% описаний – в посадках деревьев и в 100% – на пустырях. В городских местообитаниях чужеродные растения встречались почти во всех (96–98%; поймы и луга, лиственные леса, скверы) или во всех (100%; хвойные леса и посадки деревьев, пустыри, петрофитные и селитебные местообитания) описаниях. В черте города чужеродные растения были зарегистрированы менее чем в половине описаний только на болотах (39% описаний).
Рис. 2.
Доли (±95% CI) описаний хотя бы с одним чужеродным видом в местообитаниях разных типов за городом (○) и в городском окружении (⚫).
Число чужеродных видов в описании. Абсолютный размах числа чужеродных видов во всем массиве и в городских местообитаниях был от 0 до 35 видов в описании, в пригородных местообитаниях – от 0 до 23 видов. Максимальное наблюдавшееся число чужеродных видов в естественном, т.е. неантропогенном, местообитании: за городом – 12 видов (в сосновом лесу), в городе – 21 вид (в лиственном лесу).
Исключая болота, естественные местообитания не сильно различались по среднему числу чужеродных видов. За городом число чужеродных видов в поймах, на лугах, в лесах, посадках и петрофитных местообитаниях варьировало в диапазоне 1.0–2.8 вида в описании (рис. 3). Попарные различия между большинством этих местообитаний по критерию Тьюки отсутствовали (P = 0.0737–0.9991 в однофакторном ANOVA, в который включили только пригородные участки). На уровнях значимости P = 0.0112–0.0336 по критерию Тьюки различались только средние числа чужеродных видов, с одной стороны, в лиственных лесах и поймах, где их было относительно мало (в среднем 1.0–1.2 вида в описании), и, с другой стороны, в посадках, где их было относительно много (в среднем 2.8 вида).
Рис. 3.
Средние числа (±95% CI) чужеродных видов в местообитаниях разных типов за городом (⚪) и в городском окружении (⚫).
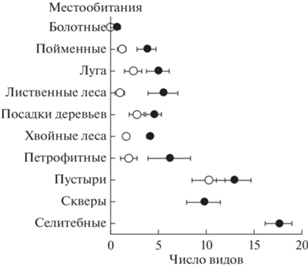
В городском окружении числа чужеродных видов в поймах, на лугах, в лесах, посадках и петрофитных местообитаниях варьировали в диапазоне 3.8–6.2 вида в описании и по критерию Тьюки не различались (P = 0.0770–0.9969 в однофакторном ANOVA, в который включили только городские участки). И в городской черте, и в пригородной зоне меньше всего чужеродных видов было на болотах (в среднем 0–0.6 вида в описании), а больше всего – в антропогенных местообитаниях (в среднем 9.8–17.6 видов), из них максимальное количество – в селитебных местообитаниях.
Таким образом, и в городе, и за городом выделили следующие группы местообитаний по возрастанию участия чужеродных растений: болота – все прочие естественные местообитания – антропогенные местообитания. Различия уровней вторжения между разными типами естественных пригородных местообитаний были меньше по амплитуде, чем различия уровней вторжения между тремя обозначенными группами. Влияние урбанизации проявлялось в том, что в одноименных местообитаниях в городе было на 2–4 чужеродных вида больше, чем за городом. Внутри типов местообитаний число чужеродных видов за городом было более изменчиво (коэффициент вариации для естественных местообитаний, исключая болота, CV = 69.5–132.8%), чем в городе (CV = = 34.7–73.0%).
Доли чужеродных видов варьировали в целом сходно с числом чужеродных видов в описании (рис. 4). Абсолютный размах доли чужеродных видов во всем массиве и в городских местообитаниях был от 0 до 56%, в пригородных местообитаниях – от 0 до 49%. Максимальная наблюдавшаяся доля чужеродных видов в естественном, т.е. неантропогенном, местообитании за городом – 22% (на лугу), в городе – 42% (в пойменном местообитании).
Рис. 4.
Средние доли (±95% CI) чужеродных видов в местообитаниях разных типов за городом (⚪) и в городском окружении (⚫).
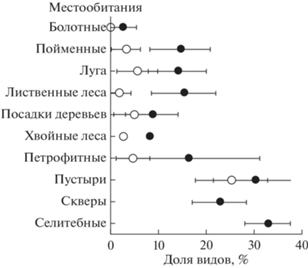
Естественные местообитания за городом по доле чужеродных видов, за исключением болот, где эти виды отсутствовали, подразделились на леса (хвойные и лиственные с долями чужеродных видов 1.8–2.7%) и прочие местообитания (поймы, луга, искусственные посадки деревьев и петрофитные местообитания с долями чужеродных видов 3.3–5.6%). По критерию Тьюки в однофакторном ANOVA, в который включили только пригородные участки, различия между лесами и прочими местообитаниями были на уровнях значимости P = 0.0004–0.0300. Однако доли чужеродных видов в поймах, на лугах, в искусственных посадках деревьев и петрофитных местообитаниях не различались (P = 0.1117–0.9998 по критерию Тьюки).
В городском окружении по доле чужеродных видов естественные местообитания (за исключением болот) были в основном гомогенны (14.0–16.2%; поймы, луга, лиственные леса и петрофитные местообитания; P = 0.0608–0.9998 по критерию Тьюки в однофакторном ANOVA, в который включили только городские участки). Невысокие значения доли чужеродных видов в городе были в хвойных лесах и посадках (8.1–8.7%), т.е. они были меньше, чем в поймах, на лугах и в лиственных лесах (P = 0.0001–0.0024 по критерию Тьюки).
В антропогенных местообитаниях в целом средние доли чужеродных видов варьировали в диапазоне 22.8–32.8%.
По признаку доли чужеродных видов, как и по их абсолютному числу выделили три группы местообитаний: болота – прочие естественные местообитания (хотя и не абсолютно гомогенные, но все же близкие между собой) – антропогенные местообитания. Влияние урбанизации проявлялось в том, что в одноименных местообитаниях в городе доля чужеродных видов была в 2–8 раз больше, чем за городом. Внутри типов местообитаний доли чужеродных видов за городом были более изменчивы (коэффициент вариации для естественных местообитаний, исключая болота, CV = 70.0–124.9%), чем в городе (CV = 36.5–73.9%).
Характерные диапазоны участия чужеродных видов в составе сообществ. Ожидаемо, что между средним числом и средней долей чужеродных видов существует сильная сопряженность (рис. 5; r = 0.97, n = 18, P ⪡ 0.0001). Средние оценки числа и долей чужеродных видов в местообитаниях разных типов использовали для определения качественно различающихся уровней вторжения (см. рис. 5). Выделили четыре дифференцированных друг от друга кластера значений числа и доли чужеродных видов.
Рис. 5.
Связь между средними числами и средними долями чужеродных видов в местообитаниях разных типов за городом (не залитые символы) и в городском окружении (залитые символы); для оценок по обеим осям приведены размахи в виде ±95% CI. Квадраты – естественные местообитания, треугольники – пустыри, ромбы – скверы и селитебные местообитания. Красными линиями ограничены кластеры наблюдений, обсуждаемые в тексте как относительно однородные.
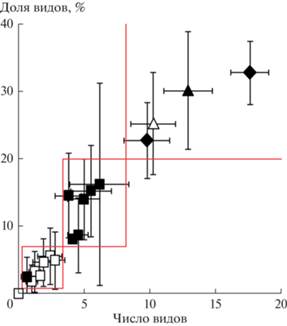
Кластер I: число чужеродных видов – ноль, доля чужеродных видов – ноль. Это пригородные болота с полным отсутствием чужеродных видов.
Кластер II: число чужеродных видов – 1–3 в описании, доля чужеродных видов – 1–7%. Сюда попали все естественные пригородные местообитания и городские болота, образующие на графике компактное скопление точек в левом нижнем углу диаграммы.
Кластер III: число чужеродных видов – 4–7 в описании, доля чужеродных видов – 8–20%. Это естественные местообитания в городском окружении. В кластере значения гомогенны по признаку числа чужеродных видов, но видна дифференцировка по доле чужеродных видов.
Кластер IV: число чужеродных видов – более 7 в описании, доля чужеродных видов – более 20%. Это антропогенные местообитания – как пригородные, так и городские. В кластере значения различаются по числу чужеродных видов, но гомогенны по их доле.
ОБСУЖДЕНИЕ
Наши данные свидетельствуют о том, что пригородные местообитания на Среднем Урале подвержены проникновению чужеродных видов растений. Большинство естественных местообитаний в радиусе 30–40 км от г. Екатеринбурга заняты растительными сообществами, в которых есть хотя бы один, а чаще – от 1 до 3 чужеродных видов. При выборе местообитаний для обследования мы старались соблюдать требования рандомизации (хотя, разумеется, с ограничениями, обусловленными требованиями логистики и бюджетом времени). Поэтому мы полагаем, что полученные оценки уровней вторжения в пригородные местообитания в целом близко отражают реальное распространение чужеродных растений.
Можно ли считать зарегистрированные в пригородных местообитаниях уровни вторжения средними региональными значениями? Немногочисленные сведения о встречаемости чужеродных растений в локалитетах Среднего Урала, расположенных не в пригородной зоне г. Екатеринбурга, позволяют считать, что местообитания в радиусе 30–40 км от города, по-видимому, не репрезентативны для региона в целом. В его окрестностях уровни вторжения чужеродных растений выше, чем в более отдаленных локалитетах. На отдаленных от города особо охраняемых территориях чужеродные растения встречаются только на антропогенно нарушенных участках, вблизи жилья, но не в лесах [18, 58]. С другой стороны, в раннесукцессионных постагрогенных лесах Среднего Урала доля чужеродных видов может составлять 2–3% [20].
Возможность сравнить уровни вторжения чужеродных растений в местообитаниях разных типов на Среднем Урале и в географически отдаленных регионах позволяет работа [8]. В ней приведены доли неофитов и археофитов в классифицированных по EUNIS местообитаниях в Чешской Республике и Великобритании (оценки, приведенные для Каталонии, мы не использовали). Часть местообитаний можно однозначно сопоставить с изученными нами. Для сравнения со своими данными, которые представляют долю чужеродных растений, мы сложили доли неофитов и археофитов, опубликованные в [8]. Значения уровней вторжения, установленные для пригородных местообитаний на Среднем Урале (СУ) и в Европе (Е), оказались в основном близкими: болота – 0% (СУ) и 0.1–0.2% (Е); луга – 5.6% (СУ) и 3.8–6% (Е); лиственные леса – 1.8% (СУ) и 1.7–4% (Е); хвойные леса – 2.7% (СУ) и 1–25.1% (Е); пустыри – 25.3% (СУ) и 27.8% (Е). Следовательно, уровни вторжения в некоторых типах местообитаний пригородных пространств в окрестностях г. Екатеринбурга и в Европе сопоставимы. Этот результат отчасти неожиданный. Для пригородных районов Екатеринбурга можно было ожидать более низких уровней вторжения, чем в Европе, учитывая различия в суровости климата, давности и интенсивности антропогенного освоения этих территорий.
Нельзя исключить, что влияние полуторамиллионного города сказывается на встречаемости чужеродных растений не только собственно в городе, но и на расстояниях до 30–40 км от него. В пользу возможности такого влияния свидетельствует большая трансформированность флоры г. Екатеринбурга, в которой из 960 зарегистрированных видов 313 чужеродные [41]. Таким образом, велика вероятность, что зарегистрированные в пригородных местообитаниях вблизи города уровни вторжения не характеризуют их региональное значение.
Уровни вторжения в разных местообитаниях. И на пригородных участках, и в городском окружении мы не обнаружили сильных различий между естественными местообитаниями по средним уровням вторжения. Только болота однозначно более закрыты для чужеродных растений, чем другие естественные местообитания. Различия уровней вторжения между прочими типами естественных местообитаний нельзя считать твердо установленными. Например, относительно малое число чужеродных видов в лиственных лесах по сравнению с прочими типами местообитаний обнаружено только за городом, но не в городском окружении. Другой пример: низкие уровни вторжения в хвойных лесах подтвердились только по доле чужеродных растений, но не по абсолютному числу в описании.
Подобно некоторым опубликованным результатам [8, 12], мы рассчитывали построить более детальный рейтинг сообществ по уровням вторжения, чем тот, который получился. Например, мы допускали, что повышенные уровни вторжения могут быть в поймах [8] или искусственных посадках деревьев. Интересно, что в г. Берлине, в зоне смешанных хвойно-широколиственных лесов, на городских территориях также не установлено большой детализации уровней вторжения, а выявлены только две группы местообитаний: 1) водно-болотные угодья и леса с низкими уровнями вторжения; 2) дворы, пустыри и зеленые городские пространства (скверы) с высокими уровнями вторжения [30]. Низкие уровни вторжения на болотах и высокие уровни в антропогенных местообитаниях, видимо, универсальные закономерности, установленные, например, в результате обобщения материалов 52 тыс. описаний в Европе [8]. Но в этой же работе [8] показано, что в приречных местообитаниях, как правило, много неофитов и археофитов, в то время как в г. Екатеринбурге и его окрестностях такой закономерности не выявлено.
На настоящем этапе исследований оптимально можно выделять три группы местообитаний по возрастанию уровней вторжения: болота – прочие естественные местообитания – антропогенные местообитания. На эти различия, обусловленные свойствами самих местообитаний или формирующихся в них сообществ, накладывается влияние внешних факторов, каким в нашей работе была урбанизация.
Уровни вторжения в зависимости от урбанизации. Уровни вторжения в естественных местообитаниях в черте города до 5–8 раз выше, чем в пригороде, а в антропогенных местообитаниях в разы выше, чем в естественных. Этот результат ожидаем, поскольку положительная зависимость между преобразованностью местообитаний в городах и адвентизацией растительности – обычная картина, наблюдающаяся в разных пространственных масштабах [7, 30, 59]. Показано, что урбанизация способствует увеличению долей археофитов и неофитов, но ее влияние меньше, чем влияние типов местообитания [3]. Мы количественно не сравнивали, как уровни вторжения зависят от урбанизации и принадлежности к разным местообитаниям, но, судя по рис. 3–5, можно предположить, что влияние этих факторов примерно сопоставимо. Механизмы, приводящие к увеличению встречаемости чужеродных растений вследствие урбанизации, хорошо изучены. Они сводятся к тому, что (1) в городах больше источников пропагул (зачатков) чужеродных растений [30, 60, 61] и (2) в городах больше нарушенных местообитаний, предоставляющих благоприятные условия для закрепления чужеродных растений [31, 62–64]. Ранее было показано, что в лесопарках г. Екатеринбурга доля чужеродных видов подлеска положительно коррелирует со степенью урбанизации [22], а доля чужеродных видов трав увеличивается вследствие краевого эффекта [23].
Показатели уровня вторжения: число и доля чужеродных видов – что лучше? Мы использовали в качестве характеристики уровня вторжения два показателя – число и долю чужеродных видов в описании. Какой из них больше подходит для количественного выражения и сравнения уровней вторжения? Несмотря на сильную взаимную обусловленность (см. рис. 5), интерпретации этих двух показателей различаются. Число чужеродных видов в описании показывает, какая часть из их регионального или ценотического пула смогла проникнуть и закрепиться в исследуемом местообитании/сообществе. Следовательно, число чужеродных видов в описании в первую очередь зависит от размера видового пула и досягаемости местообитания для зачатков чужеродных растений. Доля чужеродных видов зависит не только от числа чужеродных видов (прямо), но и от числа аборигенных видов в описании (обратно). Поэтому второй показатель больше, чем первый, учитывает благоприятность условий в местообитании и больше обусловлен потенциальным ценотическим фильтром на пути закрепления чужеродных растений в сообществах.
Несомненно, что доля чужеродных видов в описании предпочтительнее при необходимости сравнения (1) разнородных местообитаний/сообществ с разными средними уровнями видового богатства или (2) описаний, выполненных по разным методикам, например описаний разной площади.
Судя по нашим результатам, чувствительность двух показателей вторжения в смысле способности выделять гомогенные группы типов местообитаний примерно одинакова (см. рис. 5). Только по числу чужеродных видов можно было бы обсуждать пять групп местообитаний: пригородные болота – прочие пригородные естественные местообитания и городские болота – естественные описания в городской черте – пустыри и скверы – дворы. Только по доле чужеродных видов можно было бы обсуждать также пять кластеров типов местообитаний: пригородные болота – прочие пригородные естественные местообитания и городские болота – хвойные леса и посадки деревьев в городской черте – прочие естественные местообитания в городской черте – антропогенные местообитания в городе и за городом.
ЗАКЛЮЧЕНИЕ
Полученные нами результаты можно выразить в виде следующих утверждений. На пригородных территориях чужеродные растения встречаются постоянно. В некоторых типах естественных местообитаний в пригороде доля чужеродных видов сопоставима со значениями этих показателей, указанными для Чешской Республики и Великобритании. Очевидно, это связано с успешным распространением чужеродных растений по территории городской агломерации г. Екатеринбурга.
Для большинства типов естественных сообществ не удалось установить особенностей уровней вторжения или выявлены лишь небольшие особенности. Фактически мы только установили, что низкий уровень вторжения характерен для болот по сравнению с прочими естественными местообитаниями. Все остальные различия между группами местообитаний относятся к сравнениям “урбанизированные местообитания по сравнению с неурбанизированными” и “естественные местообитания по сравнению с антропогенными”. Вывод о низкой дифференциации типов естественных местообитаний по уровням вторжений не абсолютен. Требуется более детальный анализ уровней вторжения в разных местообитаниях с учетом таких характеристик, как наличие, возраст, видовое богатство и сомкнутость древесного яруса; видовое богатство или обилие травяно-кустарничкового яруса.
Наши данные подтверждают, что урбанизация обусловливает значительное и однонаправленное в разных типах местообитаний увеличение уровней вторжения. Это заключение вполне предсказуемо, хотя априорно для района нашего исследования степень различия между урбанизированными и неурбанизированными условиями по числу чужеродных видов была не известна.
Исследование выполнено за счет гранта Российского научного фонда № 22-24-20149, https:// rscf.ru/project/22-24-20149/ (проект финансируется совместно РНФ и Правительством Свердловской области).
Авторы декларируют отсутствие конфликта интересов.
Список литературы
Davis M.A., Grime J.P., Thompson K. Fluctuating resources in plant communities: a general theory of invasibility // Journal of Ecology. 2000. V. 88. P. 528–536. https://doi.org/10.1046/j.1365-2745.2000.00473.x
Акатов В.В., Акатова Т.В., Чефранов С.Г. и др. Уровень полночленности и потенциал инвазибельности растительных сообществ: гипотеза соотношения видовых фондов // Журн. общ. биол. 2009. Т. 70. № 4. С. 328–338.
Chytrý M., Wild J., Pyšek P. et al. Projecting trends in plant invasions in Europe under different scenarios of future land-use change // Global Ecology and Biogeography. 2012. V. 21. P. 75–87. https://doi.org/10.1111/j.1466-8238.2010.00573.x
Daehler C.C. Invisibility of tropical islands by introduced plants: partitioning the influence of isolation and propagule pressure // Preslia. 2006. V. 78. P. 389–404.
Richardson D.M., Pyšek P. Plant invasions: merging the concepts of species invasiveness and community invisibility // Progress in Physical Geography: Earth and Environment. 2006. V. 30. № 3. P. 409–431. https://doi.org/10.1191/0309133306pp490pr
Von der Lippe M., Kowarik I. Do cities export biodiversity? Traffic as dispersal vector across urban–rural gradients // Diversity and Distributions. 2008. V. 14. https://doi.org/10.1111/j.1472-4642.2007.00401.x
Aronson M.F.J., Handel S.N., La Puma I.P. et al. Urbanization promotes alien woody species and diverse plant assemblages in the New York metropolitan region // Urban Ecosyst. 2015. V. 18. № 1. P. 31–45. https://doi.org/10.1007/s11252-014-0382-z
Chytrý M., Jarošík V., Pyšek P. et al. Separating habitat invasibility by alien plants from the actual level of invasion // Ecology. 2008. V. 89. № 6. P. 1541–1553. https://doi.org/10.1890/07-0682.1. PMID: 18589519
Chytrý M., Pyšek P., Wild J. et al. European map of alien plant invasions based on the quantitative assessment across habitats // Diversity and Distributions. 2009. № 15. P. 98–107. https://doi.org/10.1111/j.1472-4642.2008.00515.x
Chytrý M., Pyšek P., Tichý L. et al. Invasions by alien plants in the Czech Republic: a quantitative assessment across habitats // Preslia. 2005. № 77. P. 339–354.
Hierro J.L., Maron J.L., Callaway R.M. A biogeographical approach to plant invasions: the importance of studying exotics in their introduced and native range // Journal of Ecology. 2005. V. 93. P. 5–15. https://doi.org/10.1111/j.0022-0477.2004.00953.x
González-Moreno P., Pino J., Carreras D. et al. Quantifying the landscape influence on plant invasions in Mediterranean coastal habitats // Landscape Ecology. 2013. V. 28. № 5. P. 891–903. https://doi.org/10.1007/s10980-013-9857-1
Акатов В.В., Акатова Т.В., Загурная Ю.С. и др. Инвазибельность растительных сообществ: прогноз на основе анализа ценотических параметров // Новые технологии. 2009. № 3. С. 112–119.
Guo H.-D., Zhang L., Zhu L.-W. Earth observation big data for climate change research // Advances in Climate Change Research. 2015. V. 6. P. 108–117. https://doi.org/10.1016/j.accre.2015.09.007
Чадаева В.А., Пшегусов Р.Х. Закономерности адвентивизации придорожных сообществ юга Российского Причерноморья // Уч. зап. Казанского ун˗та. Серия: Естественные науки. 2021. Т. 163. № 1. С. 115–136. https://doi.org/10.26907/2542-064X.2021.1.115-136
Силаева Т.Б. Значение флористических данных для оценки степени биологического загрязнения среды // Сравнительная флористика на рубеже III тысячелетия: достижения, проблемы, перспективы. СПб., 2000. С. 307–311.
Горчаковский П.Л., Золотарева Н.В., Коротеева Е.В., Подгаевская Е.Н. Фиторазнообразие Ильменского заповедника в системе охраны и мониторинга. Екатеринбург: Гощицкий, 2005. 192 с.
Горчаковский П.Л., Телегова О.В. Сравнительная оценка уровня синантропизации растительного покрова особо охраняемых природных территорий // Экология. 2005. № 6. С. 403–408. [Gorchakovskii P.L., Telegova O.V. Comparative assessment of the level of plant cover synanthropization in specially protected areas // Russ. J. of Ecology. 2005. V. 36. № 6. P. 365–370.]
Назаренко М.Н. Синантропизация флоры и растительности национальных парков “Зюраткуль” и “Таганай” // Вестник Тамбовского ун-та. Серия: Естественные и технические науки. 2009. Т. 14. № 2. С. 436–440.
Пустовалова Л.А., Никонова Н.Н. Антропогенная трансформация постаграрных лесов юго-восточной части Свердловской области [Электронный ресурс] // Вестник Оренбургского гос. педагогического ун-та. Электронный научный журнал. 2018. № 4(28). С. 37–43. https://doi.org/10.32516/2303-9922.2018.28.3.
Акатов В.В., Акатова Т.В., Ескина Т.Г. Факторы варьирования числа адвентивных видов в травяных сообществах Западного Кавказа // Экология. 2010. № 5. С. 344–351. [Akatov V.V., Akatova T.V., Eskina T.G. Factors of variation in the number of adventive species in herbaceous communities of the Western Caucasus // Russ. J. of Ecology. 2010. V. 41. № 5. P. 386–392. doi: 10.1134/S1067413610050048]
Веселкин Д.В., Коржиневская А.А. Пространственные факторы адвентизации подлеска в лесопарках крупного города // Изв. РАН. Сер. географич. 2018. № 4. С. 54–64. https://doi.org/10.1134/S2587556618040167
Веселкин Д.В., Коржиневская А.А., Подгаевская Е.Н. Краевой эффект для травяно-кустарничкового яруса урбанизированных южно-таежных лесов // Экология. 2018. № 6. С. 411–420. https://doi.org/10.1134/S0367059718060136
Veselkin D.V., Dubrovin D.I., Pustovalova L.A. High canopy cover of invasive Acer negundo L. affects ground vegetation taxonomic richness // Scientific Reports. 2021. V. 11. Art. 20758. P. 1–12. https://doi.org/10.1038/s41598-021-00258-x
Морозова О.В., Царевская Н.Г. Участие чужеродных видов сосудистых растений во флорах заповедников Европейской России // Изв. РАН. Сер. географич. 2010. № 4. С. 54–62.
Баранова О.Г., Третьякова А.С., Лунева Н.Н. и др. Межрегиональные особенности таксономического состава сегетальных флор // Труды по прикладной ботанике, генетике и селекции. 2022. Т. 183. № 1. С. 174–187. https://doi.org/10.30901/2227-8834-2022-1-174-18
Сенатор С.А., Костина Н.В., Саксонов С.В. Зависимость видового разнообразия урбанофлор от ряда факторов // Вестн. Удмурт. ун-та. Серия: Биология. Науки о Земле. 2013. №. 2. С. 23–29.
Третьякова А.С., Веселкин Д.В., Сенатор С.А. и др. Факторы богатства флор городов Урало-Поволжского региона // Экология. 2018. № 3. С. 165–173. [Tretyakova A.S., Veselkin D.V., Senator S.A. et al. Factors of richness of urban floras in the Ural-Volga region // Russ. J. of Ecology. 2018. V. 49. № 3. P. 201–208. https://doi.org/10.1134/S1067413618030098]https://doi.org/10.7868/S0367059718030010
Pyšek P., Chytrý M., Perg J. et al. Plant invasions in the Czech Republic: current state, introduction dynamics, invasive species and invaded habitats // Preslia. 2012. V. 84. № 3. P. 575–629.
Kowarik I., von der Lippe M., Cierjacks A. Prevalence of alien versus native species of woody plants in Berlin differs between habitats and at different scales // Preslia. 2013. V. 85. № 2. P. 113–132.
McDonald R.I., Urban D.L. Edge effects on species composition and exotic species abundance in the North Carolina Piedmont // Biol. Invasions. 2006. V. 8. № 5. P. 1049–1060. https://doi.org/10.1007/s10530-005-5227-5
Загурная Ю.С. Площадь и степень синантропизации и адвентивизации фрагментов широколиственных лесов Западного Предкавказья // Экологический вестник Северного Кавказа. 2010. Т. 6. № 4. С. 80–84.
Vilà M., Corbin J.D., Dukes J.S. et al. Linking plant invasions to environmental change // Terrestrial ecosystems in a changing world. Eds. Canadell J., Pataki D., Pitelka L. Berlin: Springer, 2006. P. 115–124.
Приказ Министерства строительства и развития инфраструктуры Свердловской области № 746-П. URL: http://www.pravo.gov66.ru/33322/ (accessed on 20.12.2022).
Итоги Всероссийской переписи населения 2020 г. URL: https://rosstat.gov.ru/vpn_popul (accessed on 20.12.2022).
Куликов П.В., Золотарева Н.В., Подгаевская Е.Н. Эндемичные растения Урала во флоре Свердловской области. Екатеринбург: Гощицкий, 2013. 612 с.
Veselkin D.V. Urbanization increases the range, but not the depth, of forest edge influences on Pinus sylvestris bark pH // Urban Forestry & Urban Greening. 2023. V. 79. 127819.https://doi.org/10.1016/j.ufug.2022.127819
Антропов К.М., Вараксин А.Н. Оценка загрязнения атмосферного воздуха г. Екатеринбурга диоксидом азота методом Land Use Regression // Экологические системы и приборы. 2011. № 8. С. 47–54.
Стурман В.И. Природные и техногенные факторы загрязнения атмосферного воздуха российских городов // Вестник Удмуртского ун˗та. Серия: Биология. Науки о земле. 2008. № 2. С. 15–29.
Шавнин С.А., Веселкин Д.В., Воробейчик Е.Л. и др. Факторы трансформации сосновых насаждений в районе города Екатеринбурга // Лесоведение. 2015. № 5. С. 346–355.
Tretyakova A.S., Yakimov B.N., Kondratkov P.V. et al. Phylogenetic diversity of urban floras in the Central Urals // Frontiers in Ecology and Evolution. 2021. V. 9. № MAR. P. 663244. https://doi.org/10.3389/fevo.2021.663244
Третьякова А.С. Распределение видового состава растений в естественных и антропогенных местообитаниях г. Екатеринбурга // Ботан. журн. 2014. Т. 99. № 11. С. 1277–1282.
Третьякова А.С. Закономерности распределения чужеродных растений в антропогенных местообитаниях Свердловской области // Российский журн. биол. инвазий. 2015. Т. 8. № 4. С. 117–128.
Веселкин Д.В., Коржиневская А.А., Подгаевская Е.Н. Состав и численность адвентивных и инвазивных кустарников и деревьев подлеска в лесопарках г. Екатеринбурга // Вестник Томского гос. ун-та. Биология. 2018. № 42. С. 102–118. [Veselkin D.V., Korzhinevskaya A.A., Podgayevskaya E.N. The species composition and abundance of alien and invasive understory shrubs and trees in urban forests of Yekaterinburg // Vestnik Tomskogo gosudarstvennogo universiteta. Biologiya = Tomsk State University Journal of Biology. 2018. V. 42. Р. 102–118. https://doi.org/10.17223/9988591/42/5. In Russian, English Summary]https://doi.org/10.17223/19988591/42/5
Veselkin D.V., Dubrovin D.I. Diversity of the grass layer of urbanized communities dominated by invasive Acer negundo // Russ. J. of Ecology. 2019. V. 50. № 5. P. 413–421. https://doi.org/10.1134/S1067413619050114
Lososová Z., Chytrý M., Tichý L. et al. Native and alien floras in urban habitats: a comparison across 32 cities of central Europe // Global Ecology and Biogeography. 2012. № 21. P. 545–555. https://doi.org/10.1111/j.1466-8238.2011.00704.x
Fenouillas P., Ah-Peng C., Amy E. et al. Quantifying invasion degree by alien plants species in Reunion Island // Austral Ecology. 2021. № 46. P. 1025–1037. https://doi.org/10.1111/aec.13048
EUNIS habitat classification. URL: https://www. eea.europa.eu/data-and-maps/data/eunis-habitat-classification-1 1 (accessed on 20.12.2022).
Виноградова Ю.К., Майоров С.Р., Хорун Л.В. Черная книга флоры Средней России: чужеродные виды растений в экосистемах Средней России. М.: Геос, 2010. 512 с.
Князев М.С., Золотарева Н.В., Подгаевская Е.Н. и др. Конспект флоры Свердловской области. Ч. I: Споровые и голосеменные растения // Фиторазнообразие Восточной Европы. 2016. Т. 10. № 4. С. 11–41.
Князев М.С., Подгаевская Е.Н., Третьякова А.С. и др. Конспект флоры Свердловской области. Ч. VI: Двудольные растения (Fabaceae – Lobeliaceae) // Фиторазнообразие Восточной Европы. 2020. Т. 14. № 3. С. 190–331. https://doi.org/10.24411/2072-8816-2020-10077
Князев М.С., Третьякова А.С., Золотарева Н.В. и др. Конспект флоры Свердловской области. Ч. II: Однодольные растения // Фиторазнообразие Восточной Европы. 2017. Т. 11. № 3. С. 4–108.
Князев М.С., Третьякова А.С., Золотарева Н.В. и др. Конспект флоры Свердловской области. Ч. III: Двудольные растения (Aristolochiaceae–Monotropaceae) // Фиторазнообразие Восточной Европы. 2018. Т. 12. № 2. С. 6–101. https://doi.org/10.24411/2072-8816-2018-10013
Князев М.С., Третьякова А.С., Золотарева Н.В. и др. Конспект флоры Свердловской области. Ч. IV: Двудольные растения (Empetraceae–Droseraceae) // Фиторазнообразие Восточной Европы. 2019. Т. 13. № 2. С. 130–196. https://doi.org/10.24411/2072-8816-2019-10046
Князев М.С., Чкалов А.В., Третьякова А.С. и др. Конспект флоры Свердловской области. Ч. V: Двудольные растения (Rosaceae) // Фиторазнообразие Восточной Европы. 2019. Т. 13. № 4. С. 305–352. https://doi.org/10.24411/2072-8816-2019-10046
Князев М.С., Подгаевская Е.Н., Золотарева Н.В. и др. Конспект флоры Свердловской области. Ч. 7: Двудольные растения (Asteraceae, Asteroideae) // Разнообразие растительного мира. 2021. № 4 (11). С. 5–33. https://doi.org/10.22281/2686-9713-2021-4-5-33
Князев М.С., Подгаевская Е.Н., Золотарева Н.В. и др. Конспект флоры Свердловской области. Ч. 8: Двудольные растения (Asteraceae, Asteroideae) // Разнообразие растительного мира. 2022. № 1 (12). С. 28–66. https://doi: 10.22281 / 2686-9713-2022-1-28-66
Пустовалова Л.А., Веселкин Д.В. Быстрые изменения растительных сообществ природных парков вследствие рекреационного использования // Экология. 2020. № 5. С. 323–331. [Pustovalova L.A., Veselkin D.V. Rapid changes in plant communities of natural parks due to recreational use // Russ. J. of Ecology. 2020. V. 51. № 5. P. 399–407. https://doi.org/10.1134/S1067413620050100]https://doi.org/10.31857/S0367059720050108
Deutschewitz K., Lausch A., Kühn I. et al. Native and alien plant species richness in relation to spatial heterogeneity on a regional scale in Germany // Global Ecol. Biogeogr. 2003. V. 12. № 4. P. 299–311. https://doi.org/10.1046/j.1466-822X.2003.00025
Pyšek P. Alien and native species in Central European urban floras: a quantitative comparison // J. Biogeogr. 1998. V. 25. № 1. P. 155–163. https://doi.org/10.1046/j.1365-2699.1998.251177.x
Hui C., Richardson D.M., Visser V. Ranking of invasive spread through urban green areas in the world’s 100 most populous cities // Biol. Invasions. 2017. V. 19. № 12. P. 3527–3539. https://doi.org/10.1007/s10530-017-1584-0
Spellerberg I.F. Ecological effects of roads and traffic: a literature review // Glob. Ecol. Biogeogr. Lett. 1998. V. 7. № 5. P. 317–333.
Nemec K.T., Allen C.R., Alai A. et al. Woody invasions of urban trails and the changing face of urban forests in the Great Plains, USA // Am. Midl. Nat. 2011. V. 165. № 2. P. 241–256. https://doi.org/10.1674/0003-0031-165.2.241
Pauchard A., Alaback P.B. Edge type defines alien plant species invasions along Pinus contorta burned, highway and clearcut forest edges // Forest Ecol. Manag. 2006. V. 223. № 1–3. P. 327–335.https://doi.org/10.1016/j.foreco.2005.11.020
Дополнительные материалы отсутствуют.