

Журнал эволюционной биохимии и физиологии, 2021, T. 57, № 1, стр. 3-16
КОНЦЕПЦИИ КАНАЛИЗИРОВАННОСТИ И ГЕНЕТИЧЕСКОЙ АССИМИЛЯЦИИ В БИОЛОГИИ РАЗВИТИЯ. СОВРЕМЕННЫЕ ПОДХОДЫ И ИССЛЕДОВАНИЯ
А. В. Спиров 1, *, В. Ф. Левченко 1, **, М. А. Сабиров 1, ***
1 Институт эволюционной физиологии и биохимии им. И.М. Сеченова РАН
194223 Санкт-Петербург, проспект Тореза, д. 44, Россия
* E-mail: alexander.spirov@gmail.com
** E-mail: lew@lew.spb.org
*** E-mail: ne-merr-eno@mail.ru
Поступила в редакцию 08.02.2020
После доработки 24.07.2020
Принята к публикации 25.07.2020
Аннотация
Две взаимосвязанные части концепции Конрада Уоддингтона – концепция канализированности (canalization) и концепция генетической ассимиляции (ассимиляции генов, genetic assimilation) – сформулированные в 40-е годы прошлого века, продолжают привлекать внимание исследователей. Это один из самых впечатляющих примеров трансдисциплинарного развития концепций.
Индивидуальное развитие любого организма происходит в условиях постоянных и неустранимых изменений факторов внешней среды и внутренних возмущений молекулярно-физиологических процессов. Учитывая, что организмы в популяции различаются генетически, реализация генетической программы должна быть поэтому относительно устойчива к генетической изменчивости.
По Уоддингтону индивидуальное развитие канализировано, т.е. происходит внутри некоторого канала условий, ограничивающего изменчивость траектории развития. Однако сильные изменения среды и значительные внутренние возмущения способны “выбрасывать” траектории индивидуального развития за пределы канала. При этом появляются аберрантные фенотипы, некоторые из которых способны участвовать в последующем отборе. При сохранении условий, систематически приводящих к такому смещению индивидуального развития, особенности этих фенотипов через ряд поколений могут зафиксироваться отбором и генетически. Иначе говоря, отбор приводит к появлению наиболее генетически подходящих при сложившихся обстоятельствах вариантов фенотипов, у которых траектории развития соответствующим образом изменены. Популяции организмов с измененными траекториями и отличающимися генотипами продолжают существовать и в дальнейшем, когда действие возмущающих факторов прекращается. Этот механизм, приводящий к ситуации “сходные фенотипы, но отличающиеся генотипы”, Уоддингтон называл ассимиляцией генов.
Недавние результаты из области системной эволюционной биологии позволили подвести количественную базу под классические концепции Конрада Уоддингтона об устойчивости индивидуального развития и генетической ассимиляции. Появилась возможность развивать эту концепцию дальше в свете новых экспериментальных и теоретических результатов. Особенный прогресс достигнут в анализе молекулярных механизмов канализированности. Обсуждение результатов этой области системной биологии, полученных средствами компьютерного моделирования в сравнении с заключениями по экспериментальным данным, и составляет предмет данной статьи.
1. ВВЕДЕНИЕ
Система воззрений Конрада Уоддингтона, известная по своим ключевым положениям о канализированности эмбрионального развития и об ассимиляции генов в микроэволюции, была сформулирована в 40–50-х годах прошлого века. Сам Уоддингтон исходил, в частности, из учения Шмальгаузена о стабилизирующем отборе, изложенного в его монографии на английском, выпущенной в Филадельфии (Пенсильвания) в 1949 г. Последующие десятилетия сам Уоддингтон и другие авторы с близкими воззрениями развивали это направление в эволюционной биологии [1–3], первые же публикации на эту тему можно найти еще в конце XIX века [4].
Идеи канализированности и ассимиляции были сформулированы изначально как остроумные заключения из небольшого числа экспериментов и наблюдений. Последующие десятилетия теоретической и экспериментальной работы позволили развить их в общебиологические учения и продемонстрировали их применимость в современной эволюционной биологии развития (т.н. “evo-devo”). Результатом работы недавних десятилетий стали не только детальные молекулярно-биологические исследования механизмов канализирования, но также и полевые исследования микроэволюционных процессов [5–11]. Не все авторы согласны с ключевыми выводами Уоддингтона и его последователей, и поэтому эта область требует новых исследований. Но сам масштаб исследований впечатляет и свидетельствует о непреходящем интересе к исходным идеям о канализированности и генетической ассимиляции.
Эти идеи получили свои, во многом неожиданные, интерпретации в областях молекулярной биологии и сравнительной молекулярной генетики в рамках направления evo-devo [5–7, 9–11] и в последние десятилетия были развиты и подтверждены экспериментально в новых принципиальных аспектах.
В настоящее время идеи Уоддингтона все больше анализируются методами современной вычислительной биологии, преимущественно средствами компьютерных моделей и, в частности, моделями эволюции генных регуляторных сетей (ГРС) [12–16 ]. В этом новом направлении наблюдается существенный прогресс (см. обзоры [17, 18]). Однако новые результаты компьютерных экспериментов нуждаются в экспериментальной верификации. И именно в этой области на стыке системной и вычислительной биологии11 публикуется больше всего работ, которым посвящен этот обзор.
Стоит отметить, что компьютерное моделирование эволюции ГРС методологически относится к современной кибернетике (т.н. computer science) и в этом плане идеи Уоддингтона выходят за рамки теории биологической эволюции и рассматриваются также в области кибернетики в масштабных компьютерных экспериментах [17, 18].
Еще одно новое приложение идей Уоддингтона – экспериментальная эволюция биомолекул, известная как направленная эволюция или “эволюция в пробирке” [19, 20]. Здесь тоже наблюдается прогресс в обобщении этих идей и формировании новых направлений на стыке синтетической биологии и биоинженерии.
Наконец, сформулированная Уоддингтоном идея эпигенетического ландшафта, являющаяся, по-сути, визуализацией концепции канализированности, оказала влияние на разработку математической теории катастроф, что неоднократно отмечал ее автор, французский математик Рене Том [21].
Таким образом, история науки демонстрирует, как наблюдения за вылуплением птенцов страуса и эмбриогенезом мушек привели к формулировке таких концепций, область приложения и развития которых уже давно вышла за границы собственно биологии. На наш взгляд, было бы важно и интересно очертить эти области.
2. ИСХОДНЫЕ ИДЕИ УОДДИНГТОНА
Две части учения Уоддингтона – концепция канализированности (canalization) индивидуального развития и концепция генетической ассимиляции (genetic assimilation) как процесса закрепления модифицированного признака – изначально взаимосвязаны. Уоддингтон заметил, что уровни фенотипических вариаций в природных популяциях обычно ниже по сравнению с ожидаемыми при наблюдаемых уровнях генетических вариаций и варьирования условий среды. Иначе говоря, хотя большинство видов, если не все, характеризуются значительной генетической вариабельностью, например, имеют разные аллели одного гена, отличающиеся из-за однонуклеотидного полиморфизма (single nucleotide polymorphism – SNP), и при этом постоянно подвергаются действию меняющихся факторов среды, фенотипическое разнообразие в пределах вида существенно ниже того, которое можно было бы ожидать по этим причинам. Из этого следует вывод, что индивидуальное развитие устойчиво к внутренним генетическим возмущениям и варьированию факторов среды. Этот эффект также называют устойчивостью (“робастностью” – robustness) индивидуального развития [1, 3, 22]. Идея канализированного развития дополнялась Уоддингтоном концепцией “генетической ассимиляции”, в соответствии с которой на генетическом уровне происходит стабилизация фенотипических изменений, вызванных действием среды [23].
Таким образом, концепция канализированного индивидуального развития исходит из наблюдений о низкой фенотипической изменчивости в популяциях, несмотря на заметное генетическое разнообразие и варьирование факторов среды [2]. В ее основу Уоддингтон положил результаты экспериментальных наблюдений за индивидуальным развитием организмов (в частности, развитие крыльев у дрозофилы), серьезные генетические или средовые (например, тепловой шок) возмущения обусловливали появление целого спектра аберрантных фенотипов, не наблюдаемых в норме. На основе наблюдений было сделано несколько выводов, в частности, о том, что индивидуальное развитие организма обладает “запасом прочности”, иначе говоря, этот процесс в норме устойчив – когда организм испытывает не сильные, но повторяющиеся возмущающие воздействия. В то же время методом искусственного отбора Уоддингтон показал, что фенотипическая пластичность, изначально проявляющаяся только при сильном воздействии, имеет под собой генетическую основу и может быть усилена генетически изменением генетического фона22.
Если бы процесс индивидуального развития не был канализирован, то вариабельность внутренних (включая генетические) и внешних факторов обусловливала бы целый спектр фенотипов. Такой спектр характеризовался бы, видимо, колоколообразной кривой частотного распределения, с редкими фенотипами по краям. Искусственный отбор позволял бы при этом отбирать желаемые редкие фенотипы и получать новые, необычные формы путем скрещивания носителей таких фенотипов. Если предположить, что аналогично может быть описан и естественный отбор (например, при резком и долговременном изменении среды или при колонизации новой территории), то должна была бы наблюдаться разница фенотипов в разных фрагментах популяции. Однако это не наблюдается, поскольку процесс индивидуального развития существенно канализирован и при отсутствии экстремальных внешних возмущений почти все особи в потомстве выглядят сходно, несмотря на генетическую вариабельность.
Из этого следует, что фактически генная организация жестко не детерминирует фенотип, а определяет его характеристики в пределах нормы реакции на варьирование факторов среды. Это называют фенотипической пластичностью.
При сохранении условий, приводящих к смещению траекторий индивидуального развития, особенности аберрантных фенотипов и их потомства через ряд поколений могут зафиксироваться отбором и генетически. Иначе говоря, отбор приведет к появлению наиболее генетически подходящих при сложившихся обстоятельствах вариантов фенотипов, у которых траектории развития соответствующим образом изменены. Со временем генетическая вариабельность уменьшится и канализированность установится в новых границах. В итоге подавляющее большинство потомства будет снова фенотипически сходно, хотя генетически будет уже отличаться от исходной популяции. Этот механизм вслед за Уоддингтоном называют ассимиляцией генов или генетической ассимиляцией [8, 22, 24].
Идея канализированности индивидуального развития непосредственно связана с концепцией Уоддингтона об эпигенетическом ландшафте. С ее помощью можно проиллюстрировать идею генетической ассимиляции. Идея эпигенетического ландшафта эволюционировала от начальных деклараций в 1940 г. не только благодаря Уоддингтону, но и многим другим авторам [2, 25–28].
Уоддингтон предложил представить развитие (эмбриона, зиготы, эмбриональной клетки) как движение шарика вниз по эпигенетическому ландшафту – генетически заданной поверхности со сложным рельефом, причем такой, что всегда имеется путь сверху, где изначально находится шарик, вниз, к “цели” развития – см. рис. 1. Таких путей (и соответственно потенциальных траекторий) может быть несколько, как и целей развития. Типичная картина событий выглядит так: шарик начинает катиться по наклонной “долине”, окруженной “холмами” – стенками долины. В этом случае развитие канализировано рельефом. За стенками-холмами находится соседняя долина. Шарик при движении подвергается возмущениям и поэтому может иногда быть выброшен за пределы своей долины и далее продолжить катиться по соседней долине с иными особенностями рельефа.
Рис. 1.
Канализированность и генетическая ассимиляция в концепции эпигенетического ландшафта. (а) Обычное развитие, когда шарик скатывается по долине х. Показана часть эпигенетического ландшафта с основной долиной, ведущей к признаку взрослого организма X и другой ветви (слева), ведущей к Y (стрелки); структуры развивающегося эмбриона не попадут на путь к Y, если только сильное внешнее возмущение (полая стрелка) не вытолкнет развивающуюся систему за пределы основной долины х. (б–г) Разные сценарии, в которых приобретается признак Y, который далее может быть включен в генотип (ассимилирован генетически). (б) Исходное внешнее возмущение (как на (а)) заменяется мутантным аллелем (темная стрелка), который появляется случайно. (в) Порог, защищающий дикий тип X, понижается из-за изменения генетического фона, и вероятность переброски шарика в долину, ведущую к Y, повышается. (г) Генотип и генетический фон изменяются так, что порог исчезает, и шарик всегда попадает в долину y.
Fig. 1. Canalization and genetic assimilation in the Waddington’s concept of epigenetic landscape. (a) Normal development, when the ball rolls down the valley x. The diagram shows a part of the epigenetic landscape with a main valley leading to the adult organism’s trait X and another branch (left) leading to Y (arrows); structures of the developing embryo will not get on the trajectory to Y unless a strong external perturbation (hollow arrow) kicks the developing system out of the main valley x. (b−d) Different scenarios, in which the trait Y is acquired, being able to be then included in the genotype (i.e. genetically assimilated). (b) The initial external perturbation (as in (а)) is replaced by a mutant allele (dark arrow) which appears by accident. (c) The threshold protecting the wild type X decreases due to a change in the genetic background, and the probability of the ball being thrown to the valley leading to Y increases. (d) Genotype and genetic background change in such a way that the threshold disappears, and the ball invariably gets into the valley y.
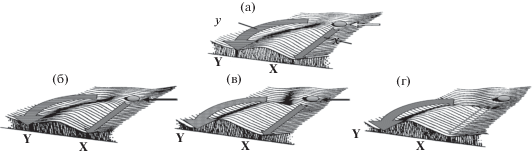
Данный наглядный подход позволяет проиллюстрировать основные идеи Уоддингтона относительно не только канализированности развития, но и генетической ассимиляции признаков. На рис. 1 сверху изображена переброска шарика сильным направленным импульсом среды из долины x, где формируется признак X, в долину y с признаком Y. Это иллюстрирует механизм приобретения признака под действием сильного и направленного внешнего фактора. Предполагается, что генетическая ассимиляция в процессе отбора связана с постепенной деформацией ландшафта таким образом, что шарик все чаще меняет траекторию и оказывается в долине y, где двигается по направлению к признаку Y, даже когда действие возмущающего фактора прекращается – рис. 1.
На протяжении последующих десятилетий идеи Уоддингтона развивались многими авторами по целому спектру новых направлений.
2.1. Развитие идей канализированности и ассимиляции в биологии
Эксперименты с искусственными возмущениями остаются главным подходом к выявлению механизмов устойчивости развития. Уоддингтон изучал простые внешние воздействия, такие как тепловой шок, эфир или разные концентрации тех или иных солей в среде [23], позднее исследователи применяли более сложные методы, приводящие к индуцированным мутациям в ключевых генах, отвечающих за процесс развития [6]. Новейшие экспериментальные подходы позволили расширить исследования до использования генетически модифицированных модельных организмов с мутантными генами, утратившими функцию (loss-of-function) или получившими новые функции (gain-of-function). Различные дозы малых интерферирующих РНК (small interfering RNA или siRNA) также использовались для модуляции активности конкретных генов (см. [32] и ссылки там). Все это позволило создать серию контролируемых по силе возмущений и тем самым увеличить разрешающую способность методов поиска изменений в регуляторных генах, обеспечивающих механизм устойчивости индивидуального развития. Во всех этих случаях относительную устойчивость оценивали количественно – как степень изменения специфических (“диагностических”) признаков после проведения эксперимента с искусственным возмущением.
В последние годы наблюдается существенный прогресс как в области системных исследований эволюции генных и клеточных регуляторных сетей, так и в области системной биологии развития, начиная с Балдвина, Шмальгаузена, Уоддингтона, а также их современников и последователей [1–4].
3. МОЛЕКУЛЯРНАЯ БИОЛОГИЯ КАНАЛИЗИРОВАННОСТИ
В современной молекулярной биологии индивидуального развития предложены механизмы, которые, как считается, обеспечивают канализированность развития. Это можно рассматривать как дальнейший прогресс в разработке концепций Уоддингтона.
3.1. Скрытая генетическая вариабельность
Скрытая генетическая вариабельность – СГВ (cryptic genetic variation), вместе с молекулярными механизмами ее обеспечивающими, рассматривается в биологии развития при обсуждении и конкретизации уоддингтоновских идей (см. [29]). Именно Уоддингтон описал проявление СГВ в своих пионерских работах [2, 22, 23].
Согласно определению Мэйзел и Троттера [30], скрытое генетическое разнообразие – имеющиеся в популяции генетические вариации, которые обычно не заметны в фенотипе. При сильных возмущениях система может потерять свою устойчивость, и тогда эффекты, связанные со скрытыми генетическими вариациями, могут проявиться в фенотипе.
Биологические системы организмов весьма устойчивы к мутационным нарушениям, генетической рекомбинации и возмущениям среды, такая устойчивость позволяет накапливать генетические вариации в скрытом состоянии. Считается, что СГВ вносит вклад в эволюционный ответ на изменения условий среды и последующий долговременный отбор. Это происходит, когда изменения условий среды ведут к сбоям в механизмах помехоустойчивости генома. Это ведет к тому, что скрытые до того мутации проявляются на уровне фенотипа, следовательно, становятся доступными для действия отбора [14, 31 ] .
В ряде работ этот аргумент находит поддержку. Особи табачного бражника Manduca sexta с мутантным фенотипом черного цвета (нормальные особи зеленые) демонстрируют разные уровни температурной чувствительности: некоторые особи после воздействия теплового шока становятся зелеными [5]. Авторы исследования произвели селекцию этих типов и успешно основали две линии – одну с выраженной температурной чувствительностью, другую без нее.
Другой пример – эволюция глаз пещерных рыбок мексиканских тетр Astyanax mexicanus [9]. Для них триггером изменений в развитии может быть перенос рыб в среду пещеры. В этих условиях рыбы испытывают физиологический стресс, ведущий к нарушению функции шаперона Hsp90. Как следствие, проявляется накопленная СГВ: появляются особи как с маленькими, так и с большими глазами. Очевидно, в условиях пещеры преимущество получают мелкоглазые или незрячие формы. Постепенно в ходе отбора в новой среде генетическое разнообразие снижается, остаются только мелкоглазые формы.
Следовательно, СГВ, способствующие накоплению изменений генома в процессе стабилизирующего отбора, при резком изменении среды могут обеспечивать изменения фенотипа.
3.2. Гены – “фенотипические конденсаторы”
В недавнее время были выявлены биомолекулярные механизмы, ответственные за удержание генетической вариабельности именно в скрытой форме, нарушение таких механизмов может эту вариабельность высвобождать [32]. Уоддингтону удалось одному из первых “высвободить” СГВ в экспериментах. Работы в этом направлении привели к формированию понятия “фенотипического конденсатора” [6].
Фенотипический конденсатор – это биологический триггерный механизм, способный при определенных условиях активировать СГВ [6]. Понятие появилось по аналогии с электрическим конденсатором, который накапливает, хранит и отдает заряд. Некоторые такие конденсаторы – это протеины и РНК, кодируемые соответствующими генами. В норме они повышают устойчивость развития, блокируя работу некоторых генов, и поэтому их дисфункция приводит к проявлению СГВ.
Как отмечают Мэйзел и Сигал [32], существует, по крайней мере, три основанных на анализе мутаций подхода к изучению устойчивости и канализированности развития, с использованием фенотипических конденсаторов. Первый подход – скрининг всего генома на наличие генетических нарушений, влияющих на разброс характеристик рассматриваемого признака. Такие работы проведены на дрожжах Saccharomyces cerevisiae и на арабидопсисе Arabidopsis thaliana [10] (см. также [32] и ссылки там).
Второй подход сосредотачивает внимание на конкретной модельной системе в процессе ее эмбрионального развития при воздействии разнообразных возмущений. Хорошим модельным объектом оказалось эмбриональное развитие вульвы у нематод рода Caenorhabditis. Эмбриогенез этого органа у нематод в норме жестко детерминирован конкретным порядком клеточных делений. Нарушение развития вульвы у C. elegans из-за мутаций или резкого изменения условий среды выявляет вариабельность в молекулярных механизмах клеточных сигнальных путей33 [33]. Компенсаторные взаимодействия между этими сигнальными путями обеспечивают стабильное развитие фенотипа [33].
При третьем подходе рассматриваются определенный вид возмущений с хорошо изученным эффектом и разнообразные биологические системы, которые ему подвергаются. Широко известный пример – белок теплового шока, шаперон Hsp90, влияющий на многие процессы развития [6]. Наиболее хорошо он изучен у дрозофилы (Drosophila melanogaster). Дисфункция Hsp90 ведет к проявлению скрытого генетического разнообразия для различных групп эволюционно далеких организмов: дрожжей, мух, позвоночных и некоторых растений [9].
Таким образом, современные молекулярно-генетические исследования демонстрируют, что канализированность реализуется функционированием фенотипических конденсаторов. Они демпфируют генотипические возмущения при нормальных условиях, тем самым способствуя накоплению СГВ. В случаях, когда нормальное функционирование такого конденсатора нарушается, наблюдается увеличение фенотипического разнообразия.
4. ФЕНОТИПИЧЕСКАЯ ПЛАСТИЧНОСТЬ И ГЕНЕТИЧЕСКАЯ АССИМИЛЯЦИЯ В ПРИРОДЕ
Уоддингтон показал, что искусственный отбор фенотипа, который проявляется лишь у нескольких особей после негенетических возмущений, может приводить к генетическому закреплению признака [22]. Позже другие исследователи наблюдали сходные эффекты для различных признаков, причем у разных видов [5, 34, 35]. Таким образом, искусственный отбор может превращать исходный фенотип через его альтернативные нестабильные варианты в новый стабильный фенотип. По результатам наблюдений природных популяций можно заключить, что во многих случаях появление фенотипа, отличающегося от предкового, может способствовать появлению новых, генетически зафиксированных адаптивных признаков [34]. Так, суровые среды обитания усиливают фенотипические различия между различными линиями плодовых мух, а увеличение температуры при отсутствии затенения приводит к увеличению частоты отклонений от нормы по морфологическим признакам [34].
Связь между устойчивостью и пластичностью может быть особенно важна для эволюции [7, 35]. Большое разнообразие фенотипов, образуемое за счет пластичности, может увеличивать выживаемость популяции в новых, враждебных для нее средах, что позволяет выигрывать время и накапливать адаптивные мутации [4]. Кратковременные возмущения, снижающие устойчивость, также могут делать в норме стабильные фенотипы пластичными, что создает материал для действия естественного отбора. Это может приводить к преимуществу нового фенотипа, или способствовать развитию механизма постепенного ответа – так называемого “генетического приспособления”, в том числе и в отсутствие исходных возмущений [5].
4.1. Фенотипическая пластичность в популяциях: наблюдения и эксперименты
В последние десятилетия выполнено немало работ по полевым исследованиям и экспериментам с популяциями разных организмов с целью охарактеризовать фенотипическую пластичность и канализированность [36]. Пластичность может способствовать адаптации на примере различных фенотипов (см. [13] и ссылки там). В качестве некоторых примеров можно привести вариации поверхностного слоя клеток жабр рыб семейства цихлид Cichlidae [11], характера окраски рачков Daphnia melanica, размера головы змей Notechis scutatus [37]. Несмотря на множество подобных примеров, роль феномена пластичности в механизмах адаптивной эволюции не является общепризнанной. До сих пор не известно, представляют ли собой данные наблюдений природных популяций или результатов искусственного отбора просто нетипичные отклонения, или же они указывают на общие механизмы адаптивной эволюции [35].
Способность к ускоренной фенотипической адаптации путем изменения пластичности может играть важную роль в долговременной устойчивости вида. Внезапные изменения среды часто происходят при инвазиях чужеродных видов, а также при колонизациях новых местообитаний.
Успех колонизаций, а именно природных инвазий или искусственных интродукций может зависеть от уровня пластичности, реализуемой в процессе адаптации к новым биотопам за пределами сложившегося ареала вида [36]. Вариабельность генетической пластичности обычно наблюдается в пределах популяции или среди нескольких популяций вида. При этом популяции, заселяющие новые или экстремальные биотопы, часто (но не всегда) демонстрируют повышенную пластичность [11].
4.2. Ассимиляция генов в популяциях: наблюдения и эксперименты
Все больше фактов свидетельствует о том, что признаки, проявляющиеся при действии негенетических (средовых или экспериментальных) факторов, важны для эволюционных инноваций. Во-первых, теоретические исследования показывают, что ассимиляция может происходить в различных условиях [13, 15, 38, 39]. Во-вторых, эксперименты в лабораторных условиях также показывают ее возможность [5, 6, 23]. В-третьих, исследования природных популяций свидетельствуют, что генетическая ассимиляция признаков, вызванная негенетическими факторами, не является редкостью [31, 40].
Эксперименты с недавно возникшей, малой популяцией показывают, что сильный искусственный отбор может способствовать быстрым изменениям фенотипа ([15] и ссылки там). Часто такие изменения достигают значений нескольких стандартных отклонений от, условно говоря, “среднего” фенотипа в течение нескольких десятков поколений [41]. Для больших популяций, испытывающих стремительные изменения условий среды в месте своего обитания, постоянный сильный направленный отбор может привести к адаптации за счет редкой аллели. Однако если соответствующий ген в гомозиготном состоянии приводит к сильному снижению приспособленности, это может помешать фиксации аллели в новых условиях, и она будет элиминирована ([15] и ссылки там). Стоит также отметить, что большие популяции при резком изменении условий среды могут распадаться на несколько, с различными фенотипами и генотипами [42], причем для каждой из новых популяций верны указанные закономерности. Например, группы организмов с генетически контролируемой право- и левосторонней симметрией тела, часто развиваются от предковых групп, у которых направление симметрии пластично, т.е. генетически не закреплено [40]. Это в ряде случаев выражается для множества признаков, например, стороны, на которой развиваются глаза у плоских рыб (Pleuronectiformes), или стороны с более развитой передней клешней у десятиногих раков (Thalassinidea) ([40] и ссылки там). Это свидетельствует, что генетическая ассимиляция варианта асимметрии признака изначально зависит от ненаследственных факторов, поскольку не детерминируется генетически. Наблюдения зависимости фенотипических и генотипических вариаций от условий среды (совместно с анализом генотипических вариаций в пределах нормы реакции) позволяют исследовать феномен канализированности и генетической ассимиляции в условиях нетипичных изменений среды.
Детальный анализ идеи генетической ассимиляции показывает, что для таковой требуется фенотипическая пластичность – способность одного генотипа давать различные фенотипы под воздействием факторов среды [31, 43]. Разнообразие ответов на внешние возмущения может также рассматриваться с позиций СГВ, которая способна проявляться в ответ на стрессирующие возмущения среды. Предполагается, что СГВ накапливается за счет нейтрального дрейфа и остается в скрытом состоянии до возмущающего воздействия [14 ] .
Когда СГВ накапливаются, это вызывает увеличение вариантов генетического фона, в рамках которого могут проявляться новые мутации. Таким образом, появление новой мутации будет зависеть от генетического фона: могут быть варианты фона, где она будет иметь частичный эффект, будет полностью проявляться, или где она вовсе не будет вызывать какого-либо эффекта. Разнообразие вариантов генетического фона дает популяции как целому доступ к новым фенотипам [24]. Этот общий аргумент в пользу вовлеченности СГВ в эволюционные процессы был отражен в математических моделях так называемых нейтральных генных сетей в генотипическом пространстве (позднее названых генотипическими сетями), и находит некоторую поддержку в экспериментальных данных ([20], см. обзор [7]).
Эмпирические исследования показывают, что для эволюционной истории инвазивных видов характерны нарушения среды обитания (см. обзор [8]). Таким нарушениям соответствуют перемены направления отбора. В рамках эволюционной генетики предполагается, что профили флуктуирующего во времени отбора могут приводить к возникновению генетических архитектур трех типов. Если изменения среды обитания стремительны, преимущество имеют генетические архитектуры, соответствующие неспециализированному фенотипу, т.е. “генералисты”. Когда изменения внешних условий происходят часто, преимущества имеют быстро эволюционирующие популяции. Наконец, если такие изменения редки – оптимизация приспособленности популяции происходит в сторону “специалистов” за счет единственного “надежного” фенотипа. Балдуин [4], Симпсон [44], а также Уоддингтон [22] предполагали, что фенотипическая пластичность может быть выгодна при появлении новых условий.
Наблюдения Уоддингтона, что искусственный отбор способен приводить к генетической ассимиляции редких фенотипов, позволяют ожидать сходного эффекта от действия естественного отбора. Наблюдения в природе и эксперименты, когда стрессирующие воздействия на популяции приводили к генетической ассимиляции, подтверждают такие ожидания. Однако оценка значимости таких механизмов в эволюции требует дальнейших исследований.
5. КАНАЛИЗИРОВАННОСТЬ В ИСКУССТВЕННОМ ОТБОРЕ МАКРОМОЛЕКУЛ
Соотношения между генотипом и фенотипом, которые лежат в основе многих разбираемых в этой статье аспектов развития концепций Уоддингтона, могут быть отслежены и для соотношений структуры и функций РНК и полипептидов.
5.1. “Фенотипическая” пластичность и устойчивость к мутациям формы молекул РНК
Первым, кто обратил внимание на применимость концепции канализированности к эволюции РНК, был Уолтер Фонтейн [19]. Известно, что первичная структура молекулы РНК (последовательность оснований) определяет в конечном итоге варианты ее геометрии (конформеры), т.е. вторичную и третичную структуры. Именно пространственная структура молекулы (ее форма и расположение активных центров) ответственна за ее функции (см., напр., [45]). Соответственно, как указал Фонтейн, с эволюционных позиций можно интерпретировать последовательность оснований как “генотип”, который подвергается мутациям, а форму молекулы (основа ее функций) как ее фенотип, подверженный возмущениям среды. Ключевым для рассуждений Фонтейна было то наблюдение, что отнюдь не всякая мутация данной РНК приводит к изменению ее конформации – вариантов конформеров может быть меньше, чем возможных последовательностей данной длины. Он показал численными экспериментами, что так понимаемое соотношение “генотип–фенотип” для РНК позволяет исследовать молекулярную эволюцию в понятиях фенотипической пластичности, эпистаза44 и канализированности.
Известные в эволюционной биологии понятия канализированности к возмущениям факторов среды и к генетическим возмущениям на уровне РНК понимаются так: средовая канализированность – это устойчивость формы молекулы к шуму факторов среды, а генетическая канализированность – это устойчивость формы молекулы к мутациям генотипа.
Ключевое обобщение своих исследований Фонтэйн сформулировал как представление о пластогенной конгруэнтности (plastogenetic congruence) [19]. Это понятие отражает коррелированность, сопряженность низкой фенотипической пластичности и низкой генетической вариабельности (нечувствительности к мутациям). Для молекулы РНК канализированность к факторам среды – это редукция пластичности посредством приобретения большей устойчивости, приводящая к единственности формы молекулы (что соответствует глубокому минимуму свободной энергии). Средовая канализированность влечет за собой генетическую канализированность в виде сокращения фенотипической новизны, возможной посредством мутаций.
При пластогенной конгруэнтности генетическая канализированность обеспечивает отбор в направлении большей канализированности к факторам среды как побочный продукт. Сходная гипотеза была выдвинута Гюнтером Вагнером и соавт. [38] на основании анализа моделей популяционной генетики. А именно: давление отбора, приводящее к редукции пластичности, может быть обусловлено снижением выживаемости (fitness cost) организмов с большей фенотипической пластичностью.
5.2. Пластичность и робастность для рибозимов
В последние годы лаборатория Андреа Вагнера [20, 46] выполнила серию работ по экспериментальному анализу эволюции рибозимов55. Общая методология этих экспериментов такова: популяции молекул рибозимов подвергаются направленной эволюции (мутированию с последующим искусственным отбором), при этом отслеживаются разнообразие конкретных последовательностей в популяции, уровень, а также специфика их ферментативной активности.
Для своих экспериментов авторы выбрали хорошо изученный рибозим из бактериальной молекулы-предшественника транспортной РНК для изолейцина. Этот рибозим катализирует сплайсинг интронов группы 1. Он очень устойчив и сохраняет активность в широком диапазоне параметров среды. Авторы рассчитывали, что такой рибозим способен накапливать мутации без потери каталитической активности (т.е. в рамках представлений о СГВ – см. выше).
Для накопления СГВ популяция рибозимов подвергалась мутагенезу, после которого производился отбор молекул, сохранивших исходную активность. Хотя накопленные мутации действительно остались скрытыми и не сказывались на каталитической активности, авторы предположили, что СГВ в этом случае могли бы влиять на скорости искусственного отбора. Далее отбор производился на каталитическую активность с химически модифицированным субстратом этого рибозима [20].
Эволюционные эксперименты “в пробирке” показали, что у популяций рибозимов с предварительно накопленными СГВ модификации каталитической активности были более вероятны, нежели у популяции без СГВ, т.е., условно говоря, они могли быстрее “находить” решение эволюционной задачи. Более того, эксперименты показали, что варианты последовательностей, пригодных для изменения каталитической активности, имелись в больших количествах в популяциях с СГВ. Таким образом, роль СГВ в адаптации к новым условиям была показана на уровне молекулярной эволюции [20, 46].
Последующие эксперименты также показали, что с помощью направленного отбора РНК можно добиться повышения чувствительности отбираемых молекул как к мутациям, так и к возмущениям факторов среды (что определяют как состояние де-канализированности) [20, 46].
6. ИДЕИ УОДДИНГТОНА И МОДЕЛИРОВАНИЕ ЭВОЛЮЦИИ ГЕННЫХ СЕТЕЙ
В современной системной биологии математические модели создаются на основе эмпирических данных. Далее результаты моделирования верифицируются новыми наблюдениями и экспериментами, дающими новый задел для развития моделей (см. обзоры [17, 47, 48]).
Можно сказать, что стержневыми для данной области являются именно работы по исследованию эволюции эмбриональных ГРС и их молекулярных компонент. Такие работы дополняются исследованиями из смежных областей (например, исследования метаболических сетей и их компонент). Недавние результаты в данной области позволили подвести количественную базу под классические концепции Конрада Уоддингтона об устойчивости индивидуального развития и ассимиляции некоторых мутационных изменений. Более того, появилась возможность развивать эту концепцию дальше в свете новых результатов [18].
6.1. Канализированность в моделях эволюции
Понимание феномена канализированности развития значительно продвинулось вперед в последние годы благодаря развитию эволюционного моделирования (обзор [17, 18]). Ряд различных вычислительных подходов подводил к идее о канализированности, выявляя различные аспекты моделируемых эволюционных механизмов. Моделирование процессов, приводящих к феномену канализированности и устойчивости, – одна из доминирующих тем в области системной биологии.
Исследованиям концепций Уоддингтона в рамках этого направления было посвящено немало работ. Современные теоретические исследования в этой области резонно начать рассматривать с работ Андреа Вагнера, начиная с его первых публикаций [49]. В дальнейшем подход был развит Вагнером и соавт., а также другими исследователями (в частности [12, 13, 20, 24, 29]).
Конкретизируя идеи Уоддингтона и близких к нему авторов, Вагнер использует термин “эпигенетической стабильности” [49]. Он трактует ее как важную составляющую уоддингтоновской канализированности развития на метафорическом эпигенетическом ландшафте. Под ней он понимает такую организацию эпигенетических взаимоотношений в индивидуальном развитии (а именно, особенностей организации ГРС), которая способна сглаживать или компенсировать генные мутации. Однако, поскольку эпигенетическая стабильность, зависящая от особенностей ГРС, сама предопределяется генетически, то естественно ожидать, что и она подвержена действию отбора. В компьютерных экспериментах Вагнер [49] показал, что длительный стабилизирующий отбор приводит к снижению доли мутаций, вызывающих серьезные нарушения паттернов генной экспрессии. Хотя при моделировании приходится абстрагироваться от ряда деталей, вывод о том, что стабилизирующий отбор приводит к эпигенетической стабильности, выглядит весьма правдоподобным.
Многие авторы в этой области рассматривают стабилизирующий отбор как одну из самых значимых эволюционных сил (например, [24, 38, 50]). Его вклад в процесс канализирования (приводящий к фенотипической устойчивости) был показан при моделировании на разных уровнях (особи, популяции). Так, Гаврилец и Хэстингс [50] моделировали ситуацию, когда генотипы производят фенотипы с различающейся чувствительностью к внутренним возмущениям процессов индивидуального развития (так называемый “шум развития”), и показали, что под действием стабилизирующего отбора канализированность развития усиливается. Моделирование на популяционно-видовом уровне в работе Вагнера и соавт. [38] показало, что канализированность, рассматриваемая в этом случае как нечувствительность по отношению к варьированию факторов среды, увеличивается с ростом силы стабилизирующего отбора. Однако нечувствительность по отношению к факторам “шума” из-за мутационного процесса увеличивается лишь ограниченно.
6.2. Канализированность и генетическая ассимиляция в моделях
В ходе классического эксперимента Уоддингтона 1953 г. личинки мухи-дрозофилы подвергались тепловому шоку, в том числе особи, представляющие фенокопии мутации, при которой нет поперечных жилок на крыльях. После отбора данного фенотипа в условиях шока далее он воспроизводился и в отсутствие возмущения. Предполагалось, что появление данных фенотипов определяется вкладом множества генов. Оказалось, что отбор в условиях теплового шока увеличивает частоту аллелей, определяющих отсутствие жилок, преодолевая барьер и обусловливая фенотип без жилок также и в отсутствие шока. В попытке альтернативного объяснения Фелконер и МакКей предположили, что тепловой шок понижает порог экспрессии этих генов (пороговая модель [41]), которая предполагает, что если отбор ассимилирующего фенотипа без жилок будет ослаблен, генетической ассимиляции не произойдет.
Джоанна Мейзел в 2004 г. [39] опубликовала результаты компьютерных экспериментов с модифицированной моделью Вагнера, выполненные с целью исследовать возможные механизмы, повлиявшие на результаты Уоддингтона. Ее работа продемонстрировала, что эффект генетической ассимиляции (в данном случае, когда приобретаемый признак теряет зависимость от “включающего” действия среды и становится наследуемым) может проявляться и без действия отбора по рассматриваемому признаку. Такой результат соответствует концепции Уоддингтона, но не соответствует объяснениям Фэлконера и МакКея [41].
Эспиноза-Сото и соавт. [13], используя классическую модель Вагнера, показали, что фенотипическая пластичность на уровне эмбриональных ГРС способна обусловливать появление новых паттернов генной активности, дающих селективные преимущества. Генотипы в данной версии модели рассматриваются как порождающие простые эмбриональные ГРС. Основной вывод авторов – генотипы, способные с некоторой вероятностью обусловливать более благоприятные в данных условиях фенотипы, при накоплении мутаций оказываются способными быстрее порождать новые генотипы, для которых более благоприятный фенотип становится генетически обусловленным [13].
Далее Эспиноза-Сото и соавт. изучали взаимоотношения между устойчивостью фенотипа по отношению к мутациям и потенциальной способностью популяции производить новый фенотипический вариант [13]. Они показали, что индивиды, характеризующиеся устойчивостью фенотипа в ответ на негенетические возмущения, эффективнее обеспечивают фенотипическую вариабельность потомков, тогда как мутации такого эффекта не дают. Авторы делают вывод, что негенетические возмущения способны чаще инициировать инновации в случае мутационной устойчивости эмбриональных ГРС.
6.3. Скрытая генетическая вариабельность в моделях
Ивасаки и соавт. [14 ] методом численных эволюционных экспериментов на популяциях моделей ГРС анализировали, как мутации и факторы среды влияют на уровень СГВ. Разные ГРС в модели рассматривались как отдельные особи, причем популяции ГРС эволюционировали при различных условиях стабилизирующего отбора. В данном случае приспособленность особи количественно оценивалась по близости поведения ГРС к эталонному, при низкой приспособленности шансы элиминации особи были выше. По окончании теста оценивалась их генетическая и фенотипическая вариабельность.
Авторы показали, что СГВ способна накапливаться и приводить к фенотипическому разнообразию в популяциях, внося свой вклад в их эволюцию [14 ] . Этот результат соответствует широко обсуждаемым гипотезам о роли СГВ в появлении новых признаков [29–32].
6.4. Пластичность и ассимиляция в моделях
Очевидно, что масштабные интродукции, реинтродукции и масштабные полевые исследования очень ресурсоемки. Поэтому использование компьютерных моделей изменчивости популяций помогает выявлять ключевые моменты процессов изменчивости, чтобы затем прицельно их исследовать в полевых условиях.
Численные эволюционные эксперименты Ленде [15, 43] имели целью оценить связь между динамикой фенотипической пластичности и генетической ассимиляцией. Рассматривалась возрастающая пластичность, когда возможны эволюционные преобразования в условиях возмущений параметров среды. Далее пластичность снижалась для фиксации и поддержания нового оптимума. В эксперименте у первого поколения в новой среде приспособленность резко падала, а типичный фенотип имел тенденцию быстро изменяться, чтобы лучше соответствовать новому оптимуму. Последующая адаптация популяции происходила в две фазы. Фаза быстрой эволюции посредством выросшей фенотипической пластичности позволяла фенотипу быстро приблизиться к новому оптимуму. Во второй фазе новый фенотип подвергался более медленному процессу генетической ассимиляции, с уменьшением пластичности. При этом происходил отбор подходящих генных мутаций, формирующих границы нового “канала”.
Лэнде дает свое определение генетической ассимиляции – это уменьшение фенотипической пластичности и замещение этой пластичности отбором генетических мутаций в изменяющейся среде. При этом подходящий фенотип, первоначально предоставленный пластичностью, закрепляется отбором для существования в условиях изменившейся среды. Это первоначальное резкое увеличение пластичности в модели и ее снижение во второй фазе эволюционных изменений весьма принципиальны. Поэтому возникает вопрос о проверке таких выводов в реальных экспериментах с реальными популяциями. Имеющиеся экспериментальные данные (как это позднее отмечал и сам Лэнде) демонстрируют более сложную картину (см. обзор [43 и ссылки там]).
В какой степени эволюционная история фенотипической пластичности может влиять на скорость адаптации к новым условиям среды? Является ли такая пластичность условием для генетической ассимиляции, или же изменения условий среды вызывают ее, увеличивая вероятность генетических изменений? Жоанна Фёрст [16], используя модель Вагнера, попыталась ответить на эти вопросы. Она, постоянно варьируя параметры среды, сравнивала популяцию, проходящую отбор на фенотипическую пластичность, с популяцией в константных условиях. Было показано, что эволюционная предыстория фенотипической пластичности может сокращать время проявления тех или иных фенотипов в результате новых мутаций и рекомбинаций. Иначе говоря, популяция, отбиравшаяся на фенотипическую пластичность, быстрее адаптируется в новой среде, и чем более жестким был такой отбор, тем быстрее идет адаптация. Вероятно, фенотипическая пластичность играет роль краткосрочной альтернативы и может облегчать адаптацию и эволюцию генотипа в дальнейшем. Как отмечает Фёрст [16], процесс адаптации популяции к новой среде предполагает в конечном итоге процессы генетической ассимиляции по Уоддингтону.
Таким образом, согласно современным системным исследованиям, когда результаты компьютерного моделирования верифицируются последующими экспериментами, генетическая ассимиляция должна существенно зависеть от таких факторов, как фенотипическая пластичность, СГВ, а также канализированность по Уоддингтону.
7. РАСШИРИТЕЛЬНЫЕ ТОЛКОВАНИЯ ИДЕЙ УОДДИНГТОНА
Возрастающий интерес к концепциям Уоддингтона в последние десятилетия нашел свое выражение и в попытках расширительных толкований и обобщений идей канализации и ассимиляции. Здесь уместно упомянуть работы Евы Яблóнки (в соавторстве с Мэрион Лэмб), где концепция ассимиляции распространяется на механизмы эпигенетического наследования в эволюции, в частности, на передачу “негенетической наследственности” на уровне поведения в эволюции животных (известная монография [51]). Труды Яблонки привлекли внимание эволюционистов, хотя и оцениваются неоднозначно [52]. Сама Яблонка всегда уделяла внимание концепциям Уоддингтона (см., например, недавнюю статью [53]).
В этой связи интересно отметить, что именно Уоддингтон предложил в 1942 г. термины эпигенотип и эпигенетика, которые он использовал в качестве концептуальной модели взаимодействия генов со своим окружением при формировании фенотипа [54]. В настоящее время эпигенетика привлекает все большее внимание, число публикаций по этой тематике растет стремительно (см., например, недавние обзоры [55, 56]). Отметим здесь же, что идея эпигенетики в последнее время активно развивается в области медицинской биологии (например, [57] и ссылки там).
Еще одним из направлений, где используются идеи Уоддингтона, является исследование гомеореза66 и устойчивости гомеостаза в процессе онтогенеза (см., например, [58]). Детальное рассмотрение этой обширной области биологии развития выходит за рамки нашего обзора.
8. ИДЕИ УОДДИНГТОНА ЗА ПРЕДЕЛАМИ БИОЛОГИИ
Как уже говорилось выше, Уоддингтон, развивая идею канализированности, ввел понятие эпигенетического ландшафта, метафорически отображающего канализированное развитие организма и влияние на развитие внутренних и внешних возмущений – см. рис. 1 [2, 25–28, 59]. Гильберт оценил ее как одно из средств интеграции генетики, эмбриологии и теории эволюции [26]. Визуализация эпигенетического ландшафта, как способ иллюстрации не только качественных, но и количественных данных, используется и в современных работах (например, [28]).
На создание и развитие Уоддингтоном идеи эпигенетического ландшафта как модели индивидуального развития повлияла теория динамических систем, истоки которой связаны с именем А. Пуанкаре. Поэтому можно сказать, что эпигенетический ландшафт – метафора динамической системы (см. [27]). Это влияние особенно заметно в поздних публикациях Уоддингтона. Так, к началу 70-х годов прошлого столетия Уоддингтон писал, что фенотип можно представить в виде ветвящейся системы траекторий, распространяющихся в фазовом пространстве вдоль временной оси [25].
Взгляды Уоддингтона близки современным представлениям о развитии нелинейных динамических систем [59]. Применение концепций нелинейной динамики для понимания функционирования генных сетей дает возможность установить соответствие между ГРС и эпигенетическим ландшафтом Уоддингтона. Последний можно представить как пространство для реализации ГРС, что объясняет множественность и гетерогенность клеточных фенотипов, определяемых одним генотипом.
Концепция эпигенетического ландшафта способствовала разработке французским математиком Рене Томом теории катастроф [21, 60]. Как отмечал сам Том [60], разработке его теории способствовало чтение руководств по эмбриологии, в частности, книг Уоддингтона, представления которого о так называемых “креодах77” и “эпигенетическом ландшафте” хорошо укладываются в абстрактную конструкцию его теории. Это, безусловно, впечатляющий случай влияния биологических идей на разработку такой общей математической теории, как теория катастроф.
Отметим также, что развитие компьютерного моделирования в связи с разработками концепций Уоддингтона приводит к результатам, интересным не только биологам, но и перспективным в области эволюционной робототехники [61].
Таким образом, за прошедшие почти восемь десятилетий концепции Конрада Уоддингтона развивались как вглубь, так и вширь, оказывая влияние не только на развитие эволюционной биологии, но и на кибернетику, некоторые разделы компьютерных наук и математики.
Список литературы
Schmalhausen I.I. Factors of Evolution: The Theory of Stabilizing Selection. Chicago. University of Chicago Press. 1949.
Waddington C.H. The Strategy of the Genes. A Discussion of Some Aspects of Theoretical Biology. London. Allen and Unwin. 1957.
Rendel J.M. Canalization of the scute phenotype of Drosophila. Evolution 13: 425–439. 1959. https://doi.org/10.1111/j.1558-5646.1959.tb03033.x
Baldwin J.M. A new factor in evolution. Am. Nat. 30: 441–451, 536–553. 1896.
Suzuki Y., Nijhout H.F. Evolution of a polyphenism by genetic accommodation. Science. 311 (5761): 650–652. 2006. https://doi.org/10.1126/science.1118888
Rutherford S.L., Lindquist S. Hsp90 as a capacitor for morphological evolution. Nature. 396: 336–342. 1998. https://doi.org/10.1038/24550
Siegal M.L., Leu J.Y. On the nature and evolutionary impact of phenotypic robustness mechanisms. Annual Review of Ecology, Evolution, and Systematics. 45: 496–517. 2014. https://doi.org/10.1146/annurev-ecolsys-120213-091705
Lee C.E., Gelembiuk G.W. Evolutionary origins of invasive populations. Evol. Appl. 1: 427–448. 2008. https://doi.org/10.1111/j.1752-4571.2008.00039.x
Rohner N., Jarosz D.F., Kowalko J.E., Yoshizawa M., Jeffery W.R., Borowsky R.L., Lindquist S., Tabin C.J. Cryptic variation in morphological evolution: HSP90 as a capacitor for loss of eyes in cavefish. Science. 342: 1372–1375. 2013. https://doi.org/10.1126/science.1240276
Hall M.C., Dworkin I., Ungerer M.C., Purugganan M. Genetics of microenvironmental canalization in Arabidopsis thaliana. Proc. Natl. Acad. Sci. U. S. A. 104: 13717–13722. 2007. https://doi.org/10.1073/pnas.0701936104
Chapman L.J., Galis F., Shinn J. Phenotypic plasticity and the possible role of genetic assimilation: Hypoxia–induced trade–offs in the morphological traits of an African cichlid. Ecol. Lett. 3 (5): 387–393. 2000. https://doi.org/10.1046/j.1461-0248.2000.00160.x
Siegal M.L., Bergman A. Waddington’s canalization revisited: developmental stability and evolution. Proc. Natl. Acad. Sci. U. S. A. 99: 10528–10532. 2002. https://doi.org/10.1073/pnas.102303999
Espinosa-Soto C., Martin O.C., Wagner A. Phenotypic plasticity can facilitate adaptive evolution in gene regulatory circuits. BMC Evol. Biol. 11: 5. doi:. 2011a.
Iwasaki W.M., Tsuda M.E., Kawata M. Genetic and environmental factors affecting cryptic variations in gene regulatory networks. BMC Evol. Biol. doi:. 2013.
Lande R. Adaptation to an extraordinary environment by evolution of phenotypic plasticity and genetic assimilation. J. Evol. Biol. 22: 1435–1446. 2009. https://doi.org/10.1111/j.1420-9101.2009.01754.x
Fierst J.L. A history of phenotypic plasticity accelerates adaptation to a new environment. J. Evol. Biol. 24:1992–2001. 2011. https://doi.org/10.1111/j.1420-9101.2011.02333.x
Spirov A., Holloway D. Using EA to study the evolution of GRNs controlling biological development. In: Noman N., Iba H. eds. Evolutionary Algorithms in Gene Regulatory Network Research. Wiley Interscience. 2015. pp. 240–268. https://doi.org/10.1002/9781119079453.ch10
Spirov A.V., Sabirov M.A., Holloway D.M. Systems evolutionary biology of Waddington’s canalization and genetic assimilation. In: Evolutionary Physiology and Biochemistry – Advances and Perspectives, Levchenko V.F. ed. InTech Press. 2018. pp. 167–185. https://doi.org/10.5772/intechopen.73662
Ancel L., Fontana W. Plasticity, evolvability and modularity in RNA. J. Exp. Zool. 288: 242–283. 2000. https://doi.org/10.1002/1097-010X(20001015)288
Hayden E.J., Ferrada E., Wagner A. Cryptic genetic variation promotes rapid evolutionary adaptation in an RNA enzyme. Nature. 474: 92–95. 2011. https://doi.org/10.1038/nature10083
Том Р. Структурная устойчивость и морфогенез. М. Логос. 2002. [René Thom. Stabilité structurelle et morphogenèse, InterÉditions, Paris, 1972].
Waddington C.H. Canalization of development and the inheritance of acquired characters. Nature. 150: 563–565. 1942. https://doi.org/10.1038/150563a0
Waddington C.H. Genetic assimilation of the bithorax phenotype. Evolution 10: 1–13. 1956. https://doi.org/0.1111/j.1558-5646.1956.tb02824.x
Wagner A. The role of robustness in phenotypic adaptation and innovation. Proc. Biol. Sci. 279: 1249–1258. 2012. https://doi.org/10.1098/rspb.2011.2293
Уоддингтон К.Х. Основные биологические концепции. На пути к теоретической биологии. I. Пролегомены. Ред. Астауров Б.Л. М. Мир. 1970. [Waddington C.H. (Editor). Towards a Theoretical Biology. vol. I. Edinburgh: Edinburgh University Press. 1968-72].
Gilbert S.F. Epigenetic landscaping: Waddington’s use of cell fate bifurcation diagrams. Biol. Philos. 6: 135–154. 1991. https://doi.org/10.1007/BF02426835
Slack J.M.M. Conrad Hal Waddington: the last Renaissance biologist? Nature Reviews Genetics. 3: 889–695. 2002. https://doi.org/10.1038/nrg933
Jamniczky H.A., Boughner J.C., Rolian C., Gonzalez P.N., Powell C.D., Schmidt E.J., Parsons T.E., Bookstein F.L., Hallgrímsson B. Rediscovering Waddington in the post–genomic age. Operationalising Waddington’s epigenetics reveals new ways to investigate the generation and modulation of phenotypic variation. Bioessays. 32: 1–6. 2010. https://doi.org/10.1002/bies.200900189
Zheng J., Payne J.L., Wagner A. Cryptic genetic variation accelerates evolution by opening access to diverse adaptive peaks. Science. 365 (6451): 347–353. 2019. https://doi.org/10.1126/science.aax1837
Masel J., Trotter M.V. Robustness and Evolvability. Trends Genet. 26 (9): 406–414. 2010. https://doi.org/10.1016/j.tig.2010.06.002
Pigliucci M., Murren C.J. Genetic assimilation and a possible evolutionary paradox: can macroevolution sometimes be so fast as to pass us by? Evolution. 57 (7): 1455–1464. 2003. https://doi.org/10.1111/j.0014-3820.2003.tb00354.x
Masel J., Siegal M.L. Robustness: mechanisms and consequences. Trends Genet. 25 (9): 395–403. 2009. https://doi.org/10.1016/j.tig.2009.07.005
Braendle C., Felix M.A. Plasticity and Errors of a Robust Developmental System in Different Environments. Dev. Cell 15: 714–724. 2008. https://doi.org/10.1016/j.devcel.2008.09.011
Kelly M. Adaptation to climate change through genetic accommodation and assimilation of plastic phenotypes. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 374 (1768): 20180176. 2019. https://doi.org/10.1098/rstb.2018.0176
Levis N.A., Pfennig D.W. Plasticity-led evolution: A survey of developmental mechanisms and empirical tests. Evol. Dev. doi.org/. 2019.https://doi.org/10.1111/ede.12309
Price T., Sol D. Introduction: Genetics of colonizing species. Am. Nat. 172: S1–S3. 2008. https://doi.org/10.1086/588639
Aubret F., Shine R. Genetic assimilation and the postcolonization erosion of phenotypic plasticity in island tiger snakes. Curr. Biol. 19: 1932–1936. 2009. https://doi.org/10.1016/j.cub.2009.09.061
Wagner G.P., Booth G., Bagheri–Chaichian H. A population genetic theory of canalization. Evolution. 51: 329–347. 1997. https://doi.org/10.1111/j.1558-5646.1997.tb02420.x
Masel J. Genetic assimilation can occur in the absence of selection for the assimilating phenotype, suggesting a role for the canalization heuristic. J. Evol. Biol. 17 (5): 1106–1110. 2004. https://doi.org/10.1111/j.1420-9101.2004.00739.x
Palmer A.R. Symmetry breaking and the evolution of development. Science. 306 (5697): 828–833. 2004. https://doi.org/10.1126/science.1103707
Falconer D.S., Mackay T.F.C. Introduction to Quantitative Genetics. Essex. Longman. 1996.
Тимофеев-Ресовский Н.В., Воронцов Н.Н., Яблоков А.В. Краткий очерк теории эволюции. М. “Наука”. 1977. [Timofeev-Ressovsky N.V., Vorontsov N.N., Yablokov A.V. Kratkiy ocherk teoriyi evoluciyi [Short Essay on Evolution Theory]. Moscow. Nauka. 1977.]
Lande R. Evolution of phenotypic plasticity in colonizing species. Mol. Ecol. doi.org/. 2015. https://doi.org/10.1111/mec.13037
Simpson G.G. The Baldwin effect. Evolution. 7: 110–117. 1953. https://doi.org/10.1111/j.1558-5646.1953.tb00069.x
Тихонов Д., Жоров Б. Методы молекулярного моделирования в изучении строения ионных каналов и их модуляции лигандами. Рос. физиол. журн. им. И.М. Сеченова. 105: 1333–1348. 2019. Tikhonov D., Zhorov B. Methods of Molecular Modeling in Studies of Ion Channels and their Modulation by Ligands. Russ. J. Physiol. 105: 1333–1348. 2019. (In Russ.)] https://doi.org/10.1134/S0869813919110116
Hayden E.J., Wagner A. Environmental change exposes beneficial epistatic interactions in a catalytic RNA. Proc. R. Soc. B. 279: 3418–3425. 2012. https://doi.org/10.1098/rspb.2012.0956
Spirov A.V., Holloway D.M. New approaches to designing genes by evolution in the computer. In Real–World Applications of Genetic Algorithms. In: InTech Press (O. Roeva ed.). 2012. https://doi.org/10.5772/36817
Spirov A.V., Holloway D.M. Using evolutionary computations to understand the design and evolution of gene and cell regulatory networks. Methods. 62: 39–55. 2013. https://doi.org/10.1016/j.ymeth.2013.05.013
Wagner A. Does evolutionary plasticity evolve? Evolution. 50: 1008–1023. 1996. https://doi.org/10.1111/j.1558-5646.1996.tb02342.x
Gavrilets S., Hastings A. A Quantitative–Genetic Model for Selection on Developmental Noise. Evolution. 48: 1478–1486. 1994. https://doi.org/10.1111/j.1558-5646.1994.tb02190.x
Jablonka E., Lamb M.J. Evolution in Four Dimensions: Genetic, Epigenetic, Behavioral, and Symbolic Variation in the History of Life. MIT Press. 2005.
Dickens T., Rahman Q. The extended evolutionary synthesis and the role of soft inheritance in evolution. Proc. Biol. Sci. 279 (1740): 2913–2921. 2012. https://doi.org/10.1098/rspb.2012.0273
Jablonka E., Noble D. Systemic Integration of Different Inheritance Systems. Current Opinion in Systems Biology. 13: 52–58. 2019. https://doi.org/10.1016/j.coisb.2018.10.002
Waddington C.H. The epigenotype (1942). Int. J. Epidemiol. 41 (1): 10–13. 2012. https://doi.org/10.1093/ije/dyr184
Noble D. Conrad Waddington and the origin of epigenetics. J. Exp. Biol. 218: 816–818. 2015. https://doi.org/10.1242/jeb.120071
Duclos K.K., Hendrikse J.L., Jamniczky H.A. Investigating the evolution and development of biological complexity under the framework of epigenetics. Evol. Dev. e12301. doi:. 2019.https://doi.org/10.1111/ede.12301
Cavalli G., Heard E. Advances in epigenetics link genetics to the environment and disease, Nature. 571: 489–499. 2019. https://doi.org/10.1038/s41586-019-1411-0
Котолупов В.А., Левченко В.Ф. Мультифункциональность и гомеостаз. Закономерности функционирования организма, важные для поддержания гомеостаза. Ж. эвол. биохим. физиол. 45: 443–451. 2009. [Kotolupov V.A., Levchenko V.F. Multifunctionality and homeostasis. Regularities of the body’s functioning, important for maintaining homeostasis. J. Evol. Biochem. and Physiol. 45: 538–547. 2009.https://doi.org/10.1134/S0022093009040148
Озернюк Н.Д., Исаева В.В. Эволюция онтогенеза. М. КМК. 2016.
Том Р. Комментарии. Динамическая теория морфогенеза. На пути к теоретической биологии. I. Пролегомены (Ред. Астауров Б.Л.) М. Мир. 1970.
Kriegman S., Cheney N., Bongard J. How morphological development can guide evolution. Sci. Rep. 8 (1): 13934. 2018.
Дополнительные материалы отсутствуют.
Инструменты
Журнал эволюционной биохимии и физиологии