

Журнал эволюционной биохимии и физиологии, 2022, T. 58, № 5, стр. 447-454
ДИНАМИКА ДВИГАТЕЛЬНОЙ АКТИВНОСТИ И ТЕМПЕРАТУРЫ ТЕЛА У ДАЛЬНЕВОСТОЧНЫХ ЛЕСНЫХ И ДОМАШНИХ КОТОВ В ОСЕННЕ-ЗИМНИЙ ПЕРИОД
В. М. Ковальзон 1, *, А. Д. Комарова 1, Г. С. Алексеева 1, М. Н. Ерофеева 1, С. В. Найденко 1
1 Институт проблем экологии и эволюции им. А.Н. Северцова РАН
Москва, Россия
* E-mail: kovalzon@sevin.ru
Поступила в редакцию 09.06.2022
После доработки 22.07.2022
Принята к публикации 24.07.2022
- EDN: YUPWJA
- DOI: 10.31857/S0044452922050059
Аннотация
У трех дальневосточных лесных котов (Prionailurus bengalensis euptilura) и четырех домашних котов (Felis catus) с предварительно вживленными (под общим наркозом) датчиками регистрировали глубокую подкожную температуру спины и двигательную активность в условиях вольерного содержания в течение двух месяцев в осенне-зимний период. Обнаружено, что осенью при положительной дневной температуре циркадианный ритм температуры тела отсутствует у дальневосточных лесных котов и слабо выражен у домашних котов. Однако он возникает в период зимних холодов в виде синхронных у всех животных колебаний с амплитудой 3–4°C, акрофазой в середине светлого периода суток и минимумом в середине ночи. При этом реальной связи с двигательной активностью у лесных котов не просматривается. Проведенная для сравнения регистрация активности и температуры “сердцевины” тела у двух домашних кошек, находящихся в это же время года в условиях лаборатории, не выявила никаких сопоставимых ритмов. Сделан вывод о том, что циркадианные биоритмы активности-покоя и подкожной температуры не являются постоянными характеристиками организма дальневосточных лесных и домашних котов, а могут возникать, исчезать и радикально изменяться при изменении окружающей температуры.
Дальневосточный (амурский) лесной кот (Prionailurus bengalensis euptilura) является северным подвидом бенгальской кошки. Этот малоизученный дикий вид обладает рядом интересных эколого-физиологических адаптаций, позволяющих ему переносить длительную и морозную зиму, хотя это животное не обладает способностью перемещаться по рыхлому снегу и не может охотиться в этот сезон [1–4]. Мы впервые предприняли попытку зарегистрировать с помощью вживленных датчиков циркадианные ритмы активности-покоя и температуры тела дальневосточных лесных котов в условиях вольерного содержания в осенне-зимний период.
Для сравнения аналогичная регистрация была параллельно проведена у домашних котов (Felis catus) в тех же условиях. Хотя кошки обоего пола были одним из самых распространенных лабораторных животных весь ХХ век и основным объектом сомнологических экспериментов, эколого-физиологические исследования на них никогда не проводились. Неизвестно, как ведет себя организм не только дальневосточных лесных, но и домашних котов при постоянном пребывании в естественных или полуестественных условиях. Есть только несколько довольно старых работ, где сопоставлялись циркадианные ритмы активности-покоя и температуры тела у кошек в лабораторных условиях, результаты которых весьма противоречивы и не очень убедительны [5–7]. Некоторые авторы полагают, что у домашних кошек вообще полностью отсутствуют закономерные суточные колебания цикла бодрствование-сон и температуры тела [8]. Чтобы убедиться в наличии (или отсутствии) циркадианной ритмики у этих животных в домашних условиях, результаты были сопоставлены с данными, полученными у двух лабораторных кошек.
МЕТОДЫ ИССЛЕДОВАНИЯ
Все экспериментальные процедуры, выполненные в данном исследовании, соответствовали этическим стандартам, утвержденным правовыми актами РФ и принципам Базельской декларации, и одобрены биоэтической комиссией Института проблем экологии и эволюции им. А.Н. Северцова РАН.
Несколько дальневосточных лесных и домашних котов с момента рождения находятся в условиях постоянного содержания в просторных индивидуальных вольерах в лесном массиве в Центре коллективного пользования “Живая коллекция диких видов млекопитающих” на территории научно-экспериментальной базы “Черноголовка” Института проблем экологии и эволюции им. А.Н. Северцова РАН. В каждой вольере имеется небольшой домик-укрытие с подстилкой из сена. Вольеры находятся при естественном освещении; ни в самих вольерах, ни на прилегающей территории дополнительной подсветки нет. Животные получают нормированный корм (200 г куриного фарша) раз в день около 18 ч 6 дней в неделю. Трем взрослым дальневосточным лесным котам (возраст 2–3 года, масса тела 5–6 кг) и четырем взрослым домашним котам (возраст 2–3 года, масса тела 4 кг) под наркозом (для дальневосточных лесных котов – 0.1 мг золетила + 0.7/0.8 мг рометара на животное; для домашних котов – 0.08 мг золетила + 0.6/0.7 мг рометара на животное; в/м) вживили через небольшой разрез на холке экологгеры (миниатюрные автономные термодатчики, снабженные акселерометрами, производства ООО “Эмби Ресерч”, Новосибирск [9]) между мышцами межлопаточной области спины, на глубину 2–3 см от поверхности тела. Разрез зашили, животным ввели антибиотик и поместили обратно в свои вольеры.
Глубокую подкожную температуру тела измеряли в °C, а двигательную активность – в условных единицах (долях G, где dG – результирующее ускорение по трем направлениям). Датчики температуры и двигательной активности производят измерения каждую минуту, усредняют их за период, установленный пользователем, и хранят на внутреннем накопителе. В данном исследовании этот период составлял 10 мин. В таком режиме заряда батареи экологгера хватает на 9–10 нед. Регистрация продолжалась 65 сут в осенне-зимний период, с 28.10.2021 по 05.01.2022. По окончании опыта животные были вновь наркотизированы таким же образом, датчики извлечены, разрез зашит, животным введен антибиотик, и они помещены обратно в свои вольеры.
Для сравнения у двух лабораторных кошек с хронически вживленными (для целей другого эксперимента) интрагастральными фистулами по И.П. Павлову была проведена аналогичная регистрация путем помещения экологгеров внутрь фистулы. У одной кошки (№ 1) регистрация была проведена в летне-осенний (22.08–01.10.2019), у другой (№ 2) – в осенне-зимний (22.10–28.12.2021) период. Все это время кошки находились в лабораторном помещении при комнатной температуре и, главным образом, естественном освещении (верхний свет не включался), вода и пища были доступны постоянно.
Данные с накопителей датчиков были считаны специальным устройством и обработаны в программе Excel с определением средних значений и стандартной ошибки (M ± SEM).
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
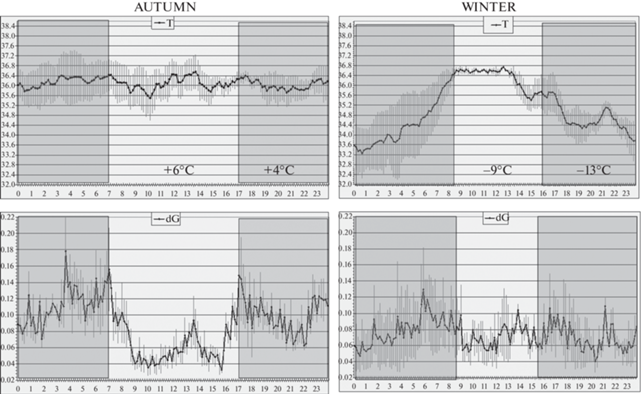
Как видно из фрагментов записей дальневосточных лесных котов (рис. 1), динамика регистрируемых показателей в осенний (31.10–10.11.2021) и зимний (26.12.2021–03.01.2022) периоды сильно различалась. Посуточное усреднение данных (рис. 2, левая часть) это наблюдение подтверждает. В осенний период, когда средняя температура воздуха составляла +6°C днем и +4°C ночью, никакого циркадианного ритма температуры тела не наблюдалось. Глубокая подкожная температура спины демонстрировала случайные колебания около “установленной точки” 36.1°С. Двигательная активность, напротив, претерпевала закономерный циркадианный сдвиг с более низкими значениями в светлое время суток, что естественно для ночных хищников. Однако с наступлением холодов (–9°C днем и –13°C ночью) динамика глубокой подкожной температуры спины значительно изменялась (рис. 2, правая часть). Ночью, с ежесуточным понижением окружающей температуры, подкожная температура дальневосточных лесных котов опускалась до 33.2°С. Примерно с полуночи начинался быстрый рост температуры тела, достигавший максимума (36.5°С) с наступлением рассвета и сохранявшийся в виде плато в течение пяти наименее холодных дневных часов. Затем с началом похолодания начинался ее постепенный спад, завершавшийся в период между полуночью и часом ночи. Среднее значение глубокой подкожной температуры спины у дальневосточных лесных котов в зимние время составляло 35.1°C, на 1°C ниже, чем в осенний период. Эти колебания температуры тела происходили синхронно и синфазно у всех трех животных, отражая, в целом, динамику температуры окружающего воздуха в зимнее время – максимальную после полудня и минимальную в ночные предутренние часы. При этом ритм двигательной активности животных сглаживался, и различие между уровнем дневной и ночной активности исчезало.
Рис. 1.
Коты дальневосточные лесные. Фрагменты 2-месячной записи. Вверху: температура тела (T, верхние кривые) и двигательная активность (dG, нижние кривые) в осенний период (с 31.10 по 10.11. 2021). Усредненные данные по трем животным (M ± SEM). По оси абсцисс – время суток в часах. По левой ординате – глубокая подкожная температура межлопаточной области спины (масштаб 10°C/шкалу). По правой ординате – показания акселерометра (масштаб 0.8 dG/шкалу). Затемненные участки – ночные периоды.
Внизу: то же в зимний период, с 26.12.2021 по 04.01.2022.
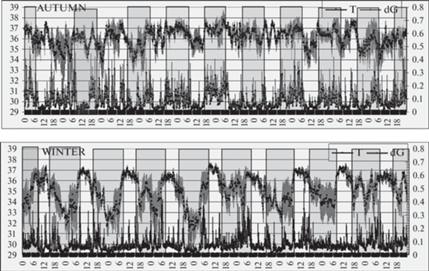
Рис. 2.
Коты дальневосточные лесные. Внутрисуточная динамика температуры тела (вверху) и двигательной активности (внизу), усреднение по 3 животным × 10 сут (M ± SEM) осенью (слева, среднесуточная температура +6°C днем и +4°C ночью) и зимой (справа, среднесуточная температура –9°C днем и –13°C ночью). По осям абсцисс – время суток в часах. По осям ординат, вверху – глубокая подкожная температура спины (масштаб 6.4°C/шкалу), внизу – показания акселерометра (масштаб 0.2 dG/шкалу). Затемненные участки – ночные периоды.
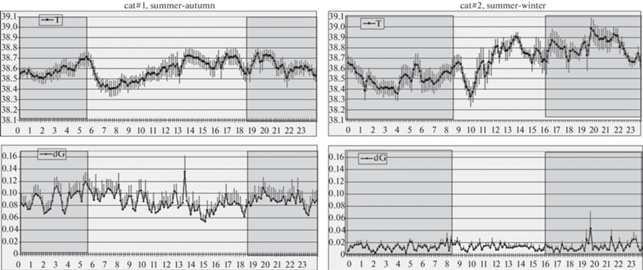
У домашних котов (рис. 3 и 4) в осенний период глубокая подкожная температура спины колебалась около “установленной точки” 36.9°C. При этом ночью, с понижением температуры воздуха, она немного опускалась, достигая минимума (36.2°С) к 5 ч утра, и затем быстро поднималась, достигая к рассвету (6:30–7 ч) 37.5°С. После рассвета подкожная температура резко падала до 36.4°C и вновь возрастала до 38°C к 13 ч (полдень по местному солнечному времени). Затем подкожная температура домашних котов претерпевала постепенное волнообразное снижение. Двигательная активность существенно не менялась, однако заметно, что оба подъема температуры тела (в 6 и 13 ч) сочетались с небольшим повышением значения dG.
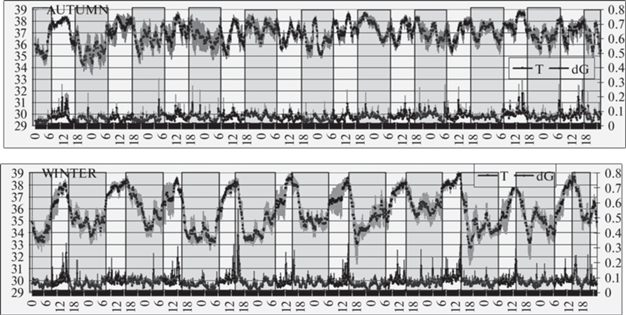
В зимнее время подкожная температура спины ночью опускалась до 34.4°С. Примерно с 3 ч ночи, несмотря на предельно низкую окружающую температуру, начинался подъем подкожной температуры, достигавший акрофазы (38.3°С) к 14 ч (когда отмечалось максимальное значение температуры окружающего воздуха), после чего происходил резкий спад, завершавшийся около 19 ч. С 19 до 3 ч температура тела домашних котов сохранялась на низком уровне 34.4°–35°С. Среднее значение глубокой подкожной температуры спины у домашних котов в зимние время составляло 35.8°С, на 1.1°С ниже, чем в осенний период. Что касается двигательной активности, то отмечался ее выраженный пик, максимум которого совпадал с акрофазой температуры тела. Все эти колебания происходили синхронно и синфазно у всех четырех котов.
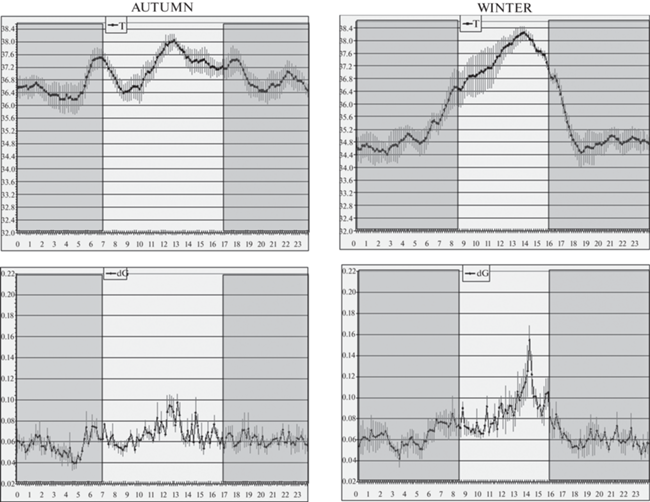
У лабораторных кошек, содержавшихся при комнатной температуре и естественном освещении, значительных и закономерных колебаний температуры датчика, находящегося в фистуле желудка, не отмечалось, за исключением небольшого (на 0.3°С) спада температуры тела в момент восхода солнца (рис. 5 и 6). В целом температура в полости желудка была гораздо выше, чем между мышцами спины (в среднем 38.5°–38.6° против 36.9°С) и более стабильна, отражая, очевидно, терморегуляцию “сердцевины” тела. Показатель двигательной активности у кошки № 1 демонстрировал небольшие апериодические подъемы, но никакой закономерной связи со световым циклом не проявлялось.
Рис. 5.
Вверху: кошка домашняя лабораторная № 1. Фрагмент 1.5-месячной записи в летне-осенний период (30.08–11.09.2019) в масштабе 10°C/шкалу (как на рис. 1 и 3). Внизу: то же, кошка № 2, зимний период (17–26.12.2021). Прочие обозначения – как на рис. 1.
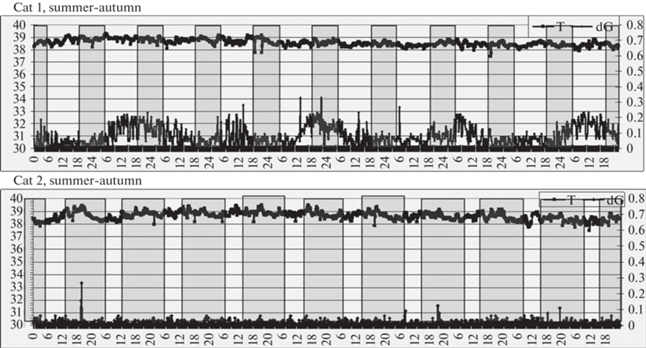
Рис. 6.
Слева: кошка домашняя лабораторная № 1, летне-осенний период.
Справа: кошка домашняя лабораторная № 2, осенне-зимний период. Масштаб: 1°С/шкалу и 0.17 dG/шкалу для верхней и нижней оси ординат соответственно.
Прочие обозначения – как на рис. 2.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Циркадианные ритмы активности и температуры тела находятся под воздействием внешних факторов – окружающей температуры и освещенности. Однако лишь у представителей нескольких видов млекопитающих проводились сезонные исследования активности – покоя и температуры тела – у овцы [10] и лошади [11], у гигантской африканской крысы [12], росомахи [13] и антилопы орикс [14]. У кошачьих подобные исследования в литературе не представлены. Настоящее исследование показывает, что циркадианные биоритмы активности-покоя и глубокой подкожной температуры тела не являются постоянными характеристиками организма дальневосточных лесных и домашних котов, а могут возникать, исчезать и радикально изменяться при изменении окружающей температуры. При положительных значениях температуры окружающего воздуха циркадианная ритмика подкожной температуры спины дальневосточных и домашних котов и кошек слабо выражена или полностью отсутствует. Обитающие в аналогичных условиях на научно-экспериментальной базе “Черноголовка” мелкие хищники – соболя и хорьки – также демонстрируют в осенний период слабо выраженную циркадианную ритмику активности – покоя и температуры брюшной полости, нечетко связанную с изменением освещенности [15]. Однако при отрицательных окружающих температурах у котов возникают ритмические колебания подкожной температуры. При этом синхронность и синфазность возникающих биоритмов у всех животных указывают на то, что они подчиняются определенному внешнему ритмоводителю (ритмоводителям). Судя по полученным нами данным, такими ритмоводителями являются суточные колебания внешней температуры и смена дня и ночи. Очевидно, что с наступлением морозной погоды организм животного, живущего в вольерных условиях, вынужден приспосабливаться, сохраняя относительное постоянство температуры “сердцевины” тела. Для этого животное предельно снижает теплоотдачу, сокращая периферический кровоток, что отражается в динамике глубокой подкожной температуры. Эти циклические колебания, в свою очередь, отражают температуру окружающего воздуха (максимальную в послеполуденное время и минимальную – в ночные предутренние часы). Интересно, что аналогичная динамика подкожной температуры, отражающая температуру окружающей среды, отмечается и при адаптации к высоким температурам у орикса [14].
Что касается двигательной активности, то дальневосточные лесные коты, накопившие к зиме толстый подкожный жировой слой, не демонстрируют никакой потребности в дополнительном мышечном разогреве. Их двигательная активность зимой в ночное время несколько снижается, а в дневное – повышается, так что в целом она зимой не выше, чем осенью. Домашние же коты, не имеющие столь мощной “термоизоляции”, вынуждены перемещаться по вольеру для дополнительного разогрева. Неожиданным при этом оказалось повышение двигательной активности в дневное время в осенний и, в особенности, зимний период у “ночных” хищников – домашних котов.
Наши опыты с регистрацией температуры “сердцевины” тела у пары домашних кошек подтверждают точку зрения Jouvet и других авторов об отсутствии циркадианных ритмов активности-покоя и температуры тела у этих животных при постоянном пребывании в условиях комнатной температуры [5, 8]. Эти ритмы формируются, очевидно, под воздействием внешних факторов, главным из которых является низкая окружающая температура. Обращает на себя внимание четкий температурный спад, возникающий с рассветом у домашних кошек. По усредненным данным этот спад оказывался весьма сходен с таковым у домашних котов, живущих в вольерных условиях при значительно более низких осенних температурах, но в 5 раз меньше по амплитуде (ср. рис. 6 и рис. 4). Этот спад отмечался при усреднении у всех шести домашних животных, участвовавших в настоящем исследовании, независимо от пола, окружающих условий и локализации датчика, и различался лишь по амплитуде. Это позволяет считать такое колебание реальным маркером циркадианной ритмики температуры тела домашних кошек.
Сопоставление настоящих результатов с нашими предыдущими [15] и литературными данными [16–19] показывает, что у млекопитающих существуют, по крайней мере, три различные стратегии адаптации к холоду.
(1) Гибернация и торпор [16–19].
(2) Повышение поведенческой активности, направленное на дополнительный разогрев тела, в 12‑часовой активный период суток, с достижением максимальной температуры тела (брюшной полости) в середине субъективной ночи, что позволяет животному (факультативным гибернаторам – монгольским хомячкам) сохранять внутреннее тепло в 12-часовые периоды поведенческого покоя (субъективно – дневные). Отражается в повышении амплитуды синхронных циркадианных ритмов температуры тела (в три раза, от 0.5 до 1.6°С) и двигательной активности (в 2 раза) [15].
(3) Использование дрожательного и недрожательного термогенеза [18] без выраженной поведенческой активации в ночной и утренний периоды с достижением максимальной подкожной температуры в середине дня (дальневосточные лесные и домашние коты). Отражается в формировании четкого циркадианного ритма подкожной температуры (с амплитудой колебаний 3–4°С) без значительных изменений (по крайней мере, у дальневосточных лесных котов) показателя двигательной активности.
Таким образом, настоящее исследование представляет собой первую попытку зафиксировать такие фундаментальные характеристики, как циркадианные ритмы температуры тела и уровня двигательной активности при различной окружающей температуре, у дальневосточных лесных котов в сопоставлении с их домашними сородичами.
Список литературы
Павлова ЕВ, Найденко СВ (2008) Неинвазивный мониторинг глюкокортикоидов в экскрементах дальневосточного лесного кота (Prionailurus bengalensis euptilurus). Зоол журн 87 (11): 1375–1381. [Pavlova EV, Naidenko SV (2008) Noninvasive monitoring of glucocorticoids in feces of the Bengal cat (Prionailurus bengalensis euptilurus). Zool J 87 (11): 1375–1381. (In Russ)].
Antonevich AL, Alekseeva GS, Vasilieva NA, Pavlova EV, Loshchagina JA., Duplyakina SYu, Naidenko SV (2019) Social play changes reflect differences in biology and development of three felids. Rus J Theriol 18 (2): 80–90. https://doi.org/10.15298/rusjtheriol.18.2.02
Seryodkin IV, Burkovskiy OA (2019) Food habit analysis of the Amur leopard cat Prionailurus bengalensis euptilurus in the Russian Far East. Biol Bull 46 (6): 648–653. https://doi.org/10.1134/S1062359019660038
Naidenko S, Chistopolova M, Hernandez-Blanco JA, Erofeeva M, Rozhnov V (2021) The effect of highway on spatial distribution and daily activity of mammals. Transp Res Part D 94: 102808. https://doi.org/10.1016/j.trd.2021.102808
Hawking F, Lobban MC, Gammage K, Worms MJ (1971) Circadian rhythms (activity, temperature, urine and microfilariae) in dog, cat, hen, duck, thamnomys and gerbillus. J Interdiscipl Cycle Res 2 (4): 455–473. https://doi.org/10.1080/09291017109359289
Kuwabara N, Seki K, Aoki K (1986) Circadian, sleep and brain temperature rhythms in cats under sustained daily light-dark cycles and constant darkness. Physiol Behav 38 (2): 283–289.
Randall W, Cunningham JT, Randall S, Liittschwager J, Johnson RF (1987) A two-peak circadian system in body temperature and activity in the domestic cat, Felis catus. J Therm Biol 12 (1): 27–37.
Jouvet M (2016) Le sommeil, la conscience et l’éveil. Paris, Odile Jacob. [Рус. пер.: Жуве М (2021) Наука о сне. Кто познает тайну сна – познает тайну мозга! М. АСТ].
Петровский ДВ, Ромащенко АВ, Троицкий СЮ, Мошкин МП (2015) Межлинейные различия мышей по температурной реакции на интраназальное введение наночастиц оксида платины. Вавиловск журн ген сел 19 (4): 439–444. [Petrovskii DV, Romashchen-ko AV, Troitskii SYu, Moshkin MP (2015) Between-strain differences in hypothermic response in mice after intranasal administration of Pto nanoparticles. Vavilovsk Zhurn Genetiki Selektsii – Vavilov J Gen Breed 19 (4): 439–444. (In Russ)]. https://doi.org/10.18699/VJ15.058
Fuchs B, Sørheim KM, Chincarini M, Brunberg E, Stubsjøen SM, Bratbergsengen K, Hvasshovd SO, Zimmermann B, Lande US, Grøva L (2019) Heart rate sensor validation and seasonal and diurnal variation of body temperature and heart rate in domestic sheep. Vet Anim Sci 8: 100075https://doi.org/10.1016/j.vas.2019.100075
Giannetto C, Aragona F, Arfuso F, Piccione G, De Caro S, Fazio F (2022) Diurnal variation in rectal and cutaneous temperatures of horses housed under different management conditions. Int J Biometeor 66 (8): 1601–1611. https://doi.org/10.1007/s00484-022-02304-3
Dzenda T, Ayo JO, Lakpini CAM, Adelaiye AB (2011) Diurnal, seasonal and sex variations in rectal temperature of African giant rats (Cricetomys gambianus, Waterhouse). J Therm Biol 36: 255–263. https://doi.org/10.1016/j.jtherbio.2011.03.010
Thiel1 A, Evans AL, Fuchs B, Arnemo JM, Aronsson M, Persson J (2019) Effects of reproduction and environmental factors on body temperature and activity patterns of wolverines. Front Zool 16: 21. https://doi.org/10.1186/s12983-019-0319-8
Davimes JG, Alagaili AN, Bhagwandin A, Bertelsen MF, Mohammed OB, Bennett NC, Manger PR, Gravett N (2018) Seasonal variations in sleep of free-ranging Arabian oryx (Oryx leucoryx) under natural hyperarid conditions. Sleep 41: 5. https://doi.org/10.1093/sleep/zsy038
Kovalzon VM, Averina OA, Minkov VA, Petrin AA, Vysokikh MYu (2020) Unusual correlation between rest–activity and body temperature rhythms in the naked mole rat (Heterocephalus glaber) as compared to five other mammalian species. J Evol Biochem Physiol 56 (5): 451–458. https://doi.org/10.1134/S0022093020050087
Пастухов ЮФ, Максимов АЛ, Хаскин ВВ (2003) Адаптация к холоду и условиям субарктики: проблемы термофизиологии, Т. 1. Магадан. МНИЦ “Арктика”. [Pastukhov YuF, Maksimov AL, Haskin VV (2003) Adaptation to cold and subarctic conditions: the problems of thermophysiology, V. 1. Magadan. ISRC “Arktika”. (In Russ)].
Mohr SM, Bagriantsev SN, Gracheva EO (2020) Cellular, molecular, and physiological adaptations of hibernation: The solution to environmental challenges. Annu Rev Cell Dev Biol 36: 13.1–13.24. https://doi.org/10.1146/annurev-cellbio-012820-09594
Frare C, Williams CT, Drew KL (2021) Thermoregulation in hibernating mammals: The role of the “thyroid hormones system”. Mol Cell Endocrinol 519: 111054. https://doi.org/10.1016/j.mce.2020.111054
Junkins MS, Bagriantsev SN, Gracheva EO (2022) Towards understanding the neural origins of hibernation. J Exp Biol 225 (1): jeb229542. https://doi.org/10.1242/jeb.229542
Дополнительные материалы отсутствуют.
Инструменты
Журнал эволюционной биохимии и физиологии