

Журнал эволюционной биохимии и физиологии, 2022, T. 58, № 6, стр. 549-557
ПРОСТРАНСТВЕННО-ВРЕМЕННЫЕ ПАТТЕРНЫ МЕЖМЫШЕЧНОГО ВЗАИМОДЕЙСТВИЯ ПРИ ЛОКОМОЦИЯХ, ВЫЗВАННЫХ ЧРЕСКОЖНОЙ ЭЛЕКТРИЧЕСКОЙ СТИМУЛЯЦИЕЙ СПИННОГО МОЗГА
С. А. Моисеев *
Великолукская государственная академия физической культуры и спорта
Великие Луки, Россия
* E-mail: sergey_moiseev@vlgafc.ru
Поступила в редакцию 15.07.2022
После доработки 16.09.2022
Принята к публикации 27.09.2022
- EDN: QKHBEI
- DOI: 10.31857/S0044452922060080
Аннотация
В работе рассматривается пространственно-временная структура мышечных синергий при произвольных локомоциях, выполняемых в условиях горизонтальной вывески нижних конечностей, и при шагоподобных движениях, модулируемых чрескожной электрической стимуляцией спинного мозга человека. Синергии извлекали разложением матриц с помощью метода главных компонент (PCA). Выявлено меньшее количество синергий при локомоциях, инициируемых чрескожной электрической стимуляцией спинного мозга. Установлено, что временнáя структура извлекаемых мышечных синергий при локомоциях в условиях электрической стимуляции спинного мозга имеет явно выраженные пики активности и высокую воспроизводимость паттернов активации. В ряде случаев они реализуются в разные временные периоды локомоторного цикла при их многократной реализации. Мышечные нагрузки в структуре выявленных синергетических модулей существенно различаются, однако векторы синергий оказываются в высокой степени схожими в разных экспериментальных условиях. Различия в параметрах пространственно-временной структуры мышечных синергий произвольных локомоций и модулируемых чрескожной электрической стимуляцией спинного мозга, вероятно, связаны с реорганизацией ритмогенерирующей части нейрональной сети, осуществляющей управление структурой локомоторного цикла.
В последнее десятилетие был достигнут существенный прогресс в исследованиях синергетических эффектов при управлении движением, однако остается большое количество нерешенных вопросов, начиная от методических аспектов их извлечения и заканчивая концептуальными вопросами о природе изучаемых синергетических феноменов. Одной из наиболее значимых для понимания природы выявляемых синергетических эффектов является область взаимодействия разных уровней ЦНС в генерации и модуляции синергий. В процессе локомоторной активности такое взаимодействие может быть достигнуто путем введения в систему управления реципрокной и коактивационной команд. Представления об этих командах в ЦНС, как об основных режимах взаимодействия пулов мотонейронов, иннервирующих пары мышц в системе агонист-антагонист, известно довольно давно, а в последнее время понятие таких команд было введено для описания контроля произвольными многосуставными движениями [1, 2]. Эти данные развивают представление об иерархически организованном управлении автоматизированными движениями посредством центральных генераторов паттернов, инициируемых простой командой вышележащих центров [3, 4].
Ранее нами было показано, что применение чрескожной электрической стимуляции спинного мозга (ЧЭССМ), вероятно, воздействуя на моторные проявления таких команд, меняет реципрокные синергетические взаимоотношения мышц нижних конечностей в структуре синергетических модулей [5]. Однако синергетические эффекты были рассмотрены в условиях произвольно выполняемых движений при ходьбе по тредбану, что накладывает определенные ограничения на работу синергетических центров в структуре спинальных нейрональных сетей, регулирующих локомоторную активность. Помимо биомеханических ограничений имеют место и другие факторы, влияющие на работу синергетических центров, такие как афферентация от опорных зон стопы, центральные команды, определяющие произвольный компонент локомоторных движений и ряд других факторов, в результате чего параметры моторного выхода оказываются довольно вариативны даже при их внутрииндивидуальном рассмотрении. В ряде работ предлагаются способы ее минимизации, однако они, преимущественно, связаны с вычислительными аспектами извлечения синергий [6]. Все это затрудняет получение прямых доказательств синергетического контроля мышечной активности со стороны центральной нервной системы.
С помощью стимуляции структур спинного и головного мозга животных разными раздражителями удавалось получить пространственно-временные профили мышечных активаций, характерные для синергии и схожие по структуре с реальными движениями [7, 8]. Несмотря на это природа наблюдаемых синергетических эффектов во многом остается неясной. Применение стимуляционных методов воздействия на структуры ЦНС в условиях, минимизирующих влияние названных выше факторов на работу синергетических центров, может дать более точную информацию о том, как параметры двигательной задачи кодируются в структуре мышечных синергий. В связи с этим целью работы явилось изучение синергетических эффектов межмышечного взаимодействия при непроизвольных локомоциях, вызываемых ЧЭССМ в условиях горизонтальной вывески нижних конечностей.
МЕТОДЫ ИССЛЕДОВАНИЯ
Эксперименты выполнены на 8 испытуемых мужского пола в возрасте от 21 до 35 лет. Исследования проведены на базе Научно-исследовательского института проблем спорта и оздоровительной физической культуры Великолукской государственной академии физической культуры и спорта в лаборатории физиологии нервной и мышечной систем. Все исследования проведены с соблюдением требований и принципов биомедицинской этики, сформулированными в Хельсинкской декларации 1964 г., и одобрены локальным биоэтическим комитетом. Каждый участник представил добровольное письменное информированное согласие на участие в исследованиях. Испытуемые располагались в горизонтальной вывеске нижних конечностей для минимизации гравитационного воздействия и облегчения возникающих в результате электростимуляционного воздействия на структуры спинного мозга шагоподобных движений [9, 10]. Протокол исследований предполагал выполнение произвольных шагательных движений в горизонтальной вывеске нижних конечностей и в условиях, минимизирующих произвольный компонент. В последнем случае испытуемым давали инструкцию не двигаться произвольно, не препятствовать и не концентрироваться на движениях нижних конечностей в случае их возникновения. Контроль отсутствия произвольного компонента в структуре вызванных локомоций осуществлялся, исходя из оценки амплитуды движений в суставах, электромиографической активности скелетных мышц и координации между нижними конечностями (рис. 1). Во всех экспериментальных условиях анализировали не менее 8 полных циклов шага. Граничные моменты полного цикла шага определяли по крайним положениям плюсневой антропометрической точки правой ноги в сагиттальной плоскости.
Рис. 1.
Расположение испытуемого в горизонтальной вывеске нижних конечностей (a), траектории перемещения антропометрических точек и образцы электромиограммы мышц нижних конечностей при произвольной ходьбе (b) и в условиях чрескожной электрической стимуляции спинного мозга (c).

Регистрировали электромиограммы (ЭМГ) билатеральных мышц нижних конечностей: передней большеберцовой (TA), медиальной головки икроножной (GM), латеральной широкой (VL), двуглавой бедра (BF), прямой бедра (RF). ЭМГ регистрировали при помощи беспроводного 16-канального биомонитора МЕ6000 (Финляндия) с частотой дискретизации 2000 Гц. Для отведения ЭМГ применяли накожные одноразовые самоклеющиеся электроды с токопроводящим гелем и активной площадью контакта 2.5 см2, 36 × 45 мм (Swaromed, Австрия). Электроды накладывались биполярно, при этом активный располагался в области проекции двигательной точки исследуемой мышцы, а референтный прикреплялся по ходу ее волокон с межэлектродным расстоянием 2 см, предусматривался дополнительный электрод заземления на каждом канале. Все зарегистрированные ЭМГ фильтровали с помощью оригинального программного обеспечения. Принцип работы фильтра состоит в поиске паттерна, характерного для артефакта стимуляции, и замены фрагмента ЭМГ, содержащего наводку, на динамическое среднее. Далее ЭМГ фильтровались полосовым фильтром с полосой пропускания 30–450 Гц, силой подавления 60 дБ с нулевой задержкой, затем усреднялись в интервалах 0.002 с и повторно применяли фильтр низких частот 15 Гц, используя программное обеспечение MegaWin. Все ЭМГ нормировались к пиковой амплитуде каждой мышцы в определенном экспериментальном условии.
ЧЭССМ осуществлялась посредством стимулятора Биокин ЭС-5 (ООО “Косима”, Россия). Катод помещали между позвонками Т11 и Т12, два анода – симметрично над гребнями подвздошных костей, частота стимуляции составляла 30 Гц. Интенсивность стимуляции подбирали индивидуально, увеличивая ток до величины, инициирующей двигательную реакцию, но не вызывающей болезненных ощущений – в диапазоне 30–90 мА. Основываясь на предыдущих результатах, показывающих, что непроизвольные шагательные движения могут быть вызваны у здоровых людей с помощью ЧЭССМ частотой 30 Гц, применяемой между позвонками T11 и T12, мы использовали те же параметры стимуляции [4]. Синхронно осуществляли видеозахват движений основных антропометрических точек сегментов тела обеих нижних конечностей: плюсневой, нижнеберцовой, верхнеберцовой, вертельной с частотой дискретизации 500 Гц. Применяли систему 3D-видеоанализа Qualisys (Швеция), включающей 8 высокоскоростных камер Oqus.
Зарегистрированные интерференционные ЭМГ, координаты антропометрических точек в системе 3D экспортировали в Statistica (StatSoft, Inc., version 10) и формировали матрицу исходных данных (X), размерностью (I × J), где I – число точек (измерений в момент времени – 500 для каждого экспериментального условия, шага, испытуемого), а J – число независимых переменных (вариационных рядов ЭМГ – 8). Помимо вариационных рядов ЭМГ в матрице создавали дополнительные переменные, позволяющие идентифицировать периоды шагательного цикла и принадлежность данных к определенному испытуемому и шагу [11, 12]. Все вариационные ряды были интерполированы относительно единой точки отсчета и стандартизированы к единице стандартного отклонения.
Из матриц извлекали компоненты (синергии) с помощью метода главных компонент (PCA). Рассматривали компоненты, имеющие собственные значения (eigenvalues) больше единицы и учитывающие не менее 10% общей дисперсии. Анализировали следующие параметры: количество извлекаемых компонент (синергий), процент общей дисперсии, учитываемый каждым фактором в общем наборе данных (VAF), матрицы нагрузок (loadings) и матрицы счетов (scores). Исходная матрица X разлагалась на произведение двух матриц: X = T × P + E, где T – матрица счетов, P – матрица нагрузок, E – матрица остатков. Матрица нагрузок несет информацию о взаимосвязи или независимости переменных относительно новых, формальных переменных, полученных в процессе разложения матриц – “векторы синергии”.
При сравнении “векторов синергий” в качестве сравниваемых вариационных рядов данных выступали значения нагрузок мышц в установленном порядке в разных экспериментальных условиях, в этом случае для сравнения вариационных рядов применяли анализ косинусного сходства (сos θ), где 1 – полное подобие, 0 – отсутствие подобия. Матрица нагрузок также включает весовые коэффициенты каждой мышцы, дающие информацию о степени вовлечения их в синергию, чем выше коэффициент, тем больше связь с новой компонентой. Рассчитывали внутрииндивидуальные средние значения весовых коэффициентов каждой мышцы для каждого извлеченного компонента при многократных реализациях полного цикла шага. Для оценки достоверности различий при сравнении среднегрупповых весовых коэффициентов применяли однофакторный дисперсионный анализ ANOVA c post-hoc анализом по критерию Newman–Keuls. Статистически значимыми различиями считали значения критерия p < 0.05.
Матрица счетов определяет временную организацию выявленных синергий и представляет собой проекции исходных данных на подпространство главных компонент – “коэффициенты активации” синергий. Коэффициенты активации представляют собой динамический процесс, отражающий изменение активности синергий во времени. Коэффициенты активации синергий сравнивали при помощи анализа максимальных значений кросскорреляционных функций (r) с учетом смещения относительно нуля, где 1 – полное соответствие, 0 – отсутствие взаимосвязи. К вариационным рядам, содержащим коэффициенты активации синергий, перед расчетом кросскорреляционных функций применяли простое экспоненциальное сглаживание (α = 0.01).
Математико-статистическая обработка данных выполнена в Statistica 10.0 и включала расчет среднего арифметического (M), ошибки среднего арифметического (SE), стандартного отклонения (SD), коэффициентов вариативности (CV). Разложение матриц осуществляли в среде Statistica, используя стандартный модуль “Advanced/Multivariate – PCA”.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
В среднем по группе испытуемых при произвольных локомоциях было установлено 4.4 ± 0.2 компонента – синергии, с долей объясняемой дисперсии 78.0 ± 1.4%. При воздействии чрескожной электрической стимуляции спинного мозга количество синергий достоверно снижалось на 22.7% и достигало 3.4 ± 0.2.
При рассмотрении пространственной структуры мышечных синергий произвольных и вызванных ЧЭССМ локомоций были установлены следующие закономерности. Наибольшие нагрузки первой синергии при произвольных движениях приходились на следующие мышцы: переднюю большеберцовую, прямую бедра правой ноги, а также на икроножную и двуглавую бедра левой нижней конечности (рис. 2). В условиях ЧЭССМ наряду с этими мышцами имели высокие коэффициенты икроножная правая и прямая мышца бедра левой ноги. Обнаружена тенденция, заключающаяся в снижении роли латеральной широкой мышцы правой стороны и икроножной левой в первой синергии при стимуляции. При этом существенно возрастала роль икроножной правой мышцы, двуглавой и прямой мышцы бедра левой нижней конечности (p < 0.05). Анализ сходства векторов первой синергии показал их высокое соответствие в обоих экспериментальных условиях, где коэффициент оказывался не менее 0.85.
Рис. 2.
Мышечные нагрузки и векторы синергий (сплошная и пунктирная линии) при произвольной ходьбе в условиях горизонтальной вывески нижних конечностей и при локомоциях, вызванных чрескожной электрической стимуляцией спинного мозга. По оси абсцисс – скелетные мышцы, по оси ординат – коэффициенты. VOL – произвольная ходьба, TCES – ходьба, вызванная стимуляцией. Заливкой выделены достоверные различия мышечных нагрузок при p < 0.05. S1, 2, 3, 4 – номер синергии.

Во второй синергии при произвольной ходьбе наибольшие нагрузки были выявлены в икроножной мышце правой ноги, а при стимуляции – в латеральной широкой левой мышце нижней конечности. Векторы второй синергии демонстрировали также высокую степень сходства – 0.75 (рис. 2). При ЧЭССМ отмечено снижение вовлечения во вторую синергию икроножной правой и прямой мышцы бедра левой ноги, а также возрастание роли латеральной широкой (р < 0.05). В третьей выявленной синергии нагрузки были невысокими и не превышали 0.5, однако векторы синергии демонстрировали высокое сходство. В четвертой синергии отмечено существенное возрастание роли двуглавой мышцы бедра правой нижней конечности при стимуляции (рис. 2).
Установлено, что коэффициенты активации первой синергии при произвольных локомоциях в условиях вывески нижних конечностей имели выраженный пик активности в первой четверти локомоторного цикла (рис. 3). Внутрииндивидуальные паттерны временнόй активации синергии демонстрировали высокое сходство – 0.79 ± 0.03, а CV не превышали 9.17%. Профили активации первой синергии при вызванных ЧЭССМ шагоподобных движениях оказались менее схожими, коэффициенты кросскорреляционных функций не превышали в среднем 0.33 ± 0.01, однако оказывались низковариативными (CV – 9.93%). Анализ соответствия временных профилей произвольных локомоторных движений и вызываемых ЧЭССМ показал среднее их соответствие – 0.41 ± 0.07, а вариативность оценивалась как средняя, CV составляли в среднем 49.91%.
Рис. 3.
Коэффициенты активации мышечных синергий при произвольной ходьбе в условиях горизонтальной вывески нижних конечностей и при локомоциях, вызванных чрескожной электрической стимуляцией спинного мозга. По оси абсцисс – прогресс цикла шага, по оси ординат – у.е. VOL – произвольная ходьба, TCES – ходьба, вызванная стимуляцией. Заливкой показаны крайние внутрииндивидуальные профили коэффициентов активации. Значения r представлены в виде M ± SD ± SE. S1, 2, 3, 4 – номер синергии.
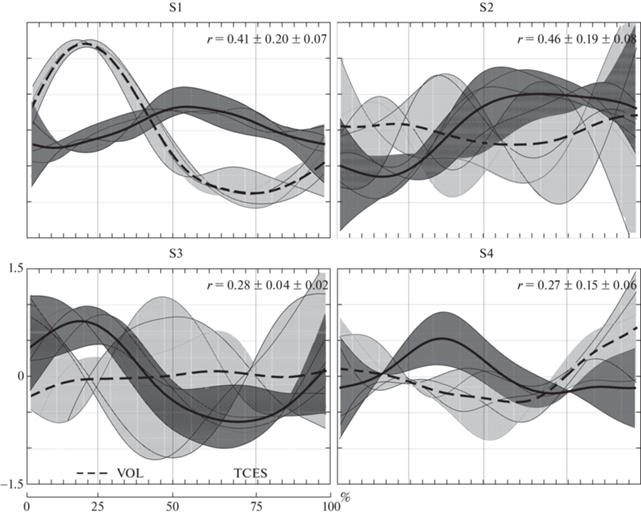
Временнáя структура второй мышечной синергии при произвольной ходьбе в горизонтальной вывеске нижних конечностей отличалась средней воспроизводимостью циклов шага и низкой их вариативностью. Схожая структура и внутрииндивидуальная вариативность была отмечена и при непроизвольных локомоциях. Так, коэффициенты кросскорреляционных функций составляли 0.48 ± ± 0.06, а коэффициенты вариативности не превышали 23.81%. При сравнении структуры второй синергии, полученной в разных экспериментальных условиях, были отмечены среднее их соответствие и средняя вариативность паттернов временнόй активации (рис. 3). Следует отметить, что во время произвольной ходьбы при внутрииндивидуальном рассмотрении отмечались пики активности синергии в разные периоды цикла шага, а при вызванных локомоциях, преимущественно, наблюдался один выраженный пик, приходящийся на третью четверть движения.
При произвольной ходьбе третья мышечная синергия характеризовалась одним пиком активности, однако внутрииндивидуальные профили структуры циклов шага были существенно смещены во времени, на что указывают средние значения кросскорреляционных функций со смещением относительно нуля – 0.50 ± 0.03 и низкие коэффициенты вариативности, не превышающие в среднем по группе 16.51%. При локомоциях, инициируемых ЧЭССМ, наблюдалась схожая картина, но практически без смещения профилей активации (рис. 3). Отмечалось низкое сходство коэффициентов активации при произвольных и вызванных локомоциях – 0.28 ± 0.02, CV – 17.45%.
Профили временнόй структуры четвертой установленной синергии демонстрировали возрастание активности к окончанию цикла шага, а при движениях, вызываемых ЧЭССМ, пик активности синергии приходился на вторую четверть цикла шага. Следует отметить низкую воспроизводимость структуры циклов при внутрииндивидуальном рассмотрении, коэффициенты кросскорреляционных функций в обоих рассматриваемых условиях локомоций оценивались как низкие – 0.29 ± 0.05 и 0.21 ± 0.01 соответственно. Сравнительный анализ временнόй структуры вызванных и произвольных локомоций показал также низкое ее сходство – 0.27 ± ± 0.06, а CV достигали 54.86%, что оценивается как средняя вариативность.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
При анализе количества извлекаемых компонент (мышечных синергий) и доли дисперсии, описываемой каждым из них, мы наблюдали меньшие значения обоих параметров при локомоциях, инициируемых ЧЭССМ. В целом такие результаты ожидаемы, поскольку даже биомеханическая структура цикла шага при локомоциях, вызываемых искусственно, несколько отличается от произвольных и сами условия их выполнения не типичны для ходьбы в вертикальном положении. Поэтому логично было бы ожидать, что и механизм управления локомоциями в таких условиях будет иметь некоторые особенности. Первое, на что хотелось бы обратить внимание – это снижение качества реконструкции исходных данных с применением PCA в условиях стимуляционного воздействия на спинной мозг. В обычных условиях синергетические взаимоотношения мышечных групп нижних конечностей регулируются спинальными генераторами паттернов, посредством реципрокных и коактивационных команд, а моторным выходом являются низковариативные стереотипные кинематические паттерны и характеристики мышечной активности [1, 2, 9]. Методами факторизации данных в таких условиях удается учесть большую часть наблюдаемой дисперсии в параметрах мышечной активности, это было показано во многих исследованиях, где VAF как критерий эффективности извлечения синергий достигает 80% и более [6, 13, 14].
Электрическая стимуляция, воздействуя на спинальные нейрональные сети, вносит изменения в их организацию (частотный код синергий), что проявляется в снижении эффективности извлечения синергий. Причем эти изменения не являются сбивающим фактором в работе спинальных нейрональных структур, а являются средством целенаправленного на них воздействия. В пользу этого утверждения свидетельствуют наши результаты, демонстрирующие четкие очертания основных пиков активности синергий и высокую воспроизводимость временнόй структуры извлекаемых мышечных синергий при локомоциях, вызываемых ЧЭССМ. При произвольных локомоциях такое наблюдается только в первой синергии. Таким образом, искусственно вызванные локомоции в условиях горизонтальной вывески нижних конечностей имеют так называемые базовые (фундаментальные) профили временнόй активации, которые можно наблюдать при вертикальной ходьбе в обычных условиях, а произвольные шагательные движения в таких условиях чаще отличаются высокой вариативностью.
Концепция двигательного синергизма предполагает снижение вычислительной нагрузки на структуры нервной системы посредством объединения элементов системы в модули с меньшей размерностью. Исходя из этого, можно утверждать, что чем больше количество модулей, тем выше сложность управляющей системы. При ходьбе в обычных условиях регистрируют от четырех до пяти мышечных синергий. Это наблюдается при анализе активности мышц одной конечности или мышц одной стороны тела при беге, ходьбе, педалировании на велоэргометре [13, 15]. Вполне вероятно, что структура мышечных синергий при локомоторных движениях, так или иначе, может включать большинство поверхностных мышц нижних конечностей, однако участие каждой из них в структуре синергетических модулей в разных условиях реализации локомоций может различаться. Нами было показано, что при сравнении локомоций, вызываемых ЧЭССМ, и произвольных, существенно различаются мышечные нагрузки в структуре первой синергии, а именно, при вызванных движениях возрастает роль GM правой, BF и RB левой ноги, снижается VL правой и GM левой. При этом векторы синергий оказываются в высокой степени схожими в разных условиях. Это свидетельствует о том, что пространственная структура естественных и вызванных локомоций имеет единый механизм управления, вероятно, реализуемый нейрональными сетями, локализованными на уровне Т11 – Т12 позвонков. Кроме того, формирование четкого очертания основных пиков активности во временнόй структуре мышечных синергий при движениях, вызываемых ЧЭССМ, свидетельствует в пользу этого утверждения.
Различия в компонентном составе мышечных синергий отмечаются даже при многократном повторении стереотипных движений, как дискретных, так и циклических. В этой связи вполне ожидаемо, что мышечные нагрузки в структуре извлекаемых нами синергий при произвольных и вызванных движениях оказались различными. Кроме того, “шагоподобная локомоторная активность” может быть представлена разными паттернами, в различной степени схожими по биомеханической структуре с реальными движениями [16]. Важными установленными факторами в нашем исследовании явилось высокое соответствие векторов синергий и низкая вариативность с четким очертанием основных пиков активности коэффициентов активации синергий при вызванных локомоциях. Такие результаты могут быть связаны со следующими положениями. Одна из теорий, описывающая сложное взаимодействие управляющих сигналов в ЦНС при локомоциях, предполагает наличие в структуре спинальных локомоторных генераторов двух независимых нейрональных сетей, осуществляющих управление ритмической активностью мышц сгибателей и разгибателей, включающих ритмогенерирующую часть и сеть, формирующую локомоторные паттерны [17–19]. Проводя аналогию с положениями концепции пространственно-временной архитектуры синергизма, коэффициенты активации мышечных синергий являются эквивалентом ритмогенерирующей части нейронной сети, управляющей локомоциями, а векторы синергий и мышечные нагрузки, определяющие соотношение активации мышечных групп, будут являться аналогией сети формирования паттернов. Таким образом, электрическая стимуляция спинного мозга на уровне Т11–Т12 позвонков, по меньшей мере, воздействует на ритмогенерирующую часть нейронной сети, что проявляется в формировании основных (фундаментальных) локомоторных временных паттернов.
Наблюдаемая нами вариативность временнόй структуры синергий при произвольной ходьбе в горизонтальной вывеске в ряде случаев связана со смещением коэффициентов активации, выявляемым при помощи кросскорреляционного анализа. Такие синергии обозначаются как “синергии, изменяющиеся во времени”, т.е. синергетические паттерны реализуются в разные временные периоды локомоторного цикла при их многократной реализации [14, 20, 21].
ЗАКЛЮЧЕНИЕ
Таким образом, временнáя структура извлекаемых мышечных синергий при локомоциях в условиях электрической стимуляции спинного мозга имеет явно выраженные пики активности и высокую воспроизводимость паттернов активации. В ряде случаев они реализуются в разные временные периоды локомоторного цикла при их многократной реализации. Мышечные нагрузки в структуре выявленных синергетических модулей существенно различаются, однако векторы синергий оказываются в высокой степени схожими в разных экспериментальных условиях. Различия в параметрах пространственно-временной структуры мышечных синергий произвольных локомоций и модулируемых ЧЭССМ, могут быть связаны с реорганизацией ритмогенерирующей части нейрональной сети, осуществляющей управление ритмической активностью мышц сгибателей и разгибателей в структуре полного цикла шага.
Список литературы
Feldman AG, Levin MF, Garofolini A, Piscitelli D, Zhang L (2021) Central pattern generator and human locomotion in the context of referent control of motor actions. Clin Neurophysiol 132 (11): 2870–2889. https://doi.org/10.1016/j.clinph.2021.08.016
Madarshahian S, Latash ML (2022) Reciprocal and coactivation commands at the level of individual motor units in an extrinsic finger flexor-extensor muscle pair. Exp Brain Res 240 (1): 321–340. https://doi.org/10.1007/s00221-021-06255-w
Аршавский ЮИ, Делягина ТГ, Орловский ГН (2015) Центральные генераторы: механизм работы и их роль в управлении автоматизированными движениями. Журн высш нерв деят им ИП Павлова 65 (2): 156–187. [Arshavsky YI, Deliagina TG, Orlovsky GN (2015) Central Pattern Generators: Mechanisms of the Activity and Their Role in the Control of “Automatic” Movements. IP Pavlov J Higher Nerv Activit 65 (2): 156–187. (In Russ)]. https://doi.org/10.7868/S0044467715020033
Gorodnichev RM, Pivovarova EA, Puhov A, Moiseev SA, Gerasimenko YP, Savochin AA, Moshonkina TR, Chsherbakova NA, Kilimnik VA, Selionov VA, Kozlovskaya IB, Edgerton VR (2012) Transcutaneous electrical stimulation of the spinal cord: a noninvasive tool for the activation of stepping pattern generators in humans. Human Physiol 38 (2): 158–167.
Moiseev SA, Gorodnichev RM (2017) Motor synergies structure variability in different intensity locomotions. Human Physiol 48 (4): 22–33. https://doi.org/10.31857/S0131164622040087
Turpin N, Uriac S, Dalleau G (2021) How to improve the muscle synergy analysis methodology? Eur J Appl Physiol 121 (4): 1009–1025. https://doi.org/10.1007/s00421-021-04604-9
Overduin S, d’Avella A, Carmena J, Bizzi E (2014) Muscle synergies evoked by microstimulation are preferentially encoded during behavior. Front Comput Neurosci 8: 20. https://doi.org/10.3389/fncom.2014.00020
Cheung VC, d’Avella A, Bizzi E (2009) Adjustments of motor pattern for load compensation via modulated activations of muscle synergies during natural behaviors. J Neurophysiol 101 (3): 1235–1257. https://doi.org/10.1152/jn.01387.2007
Гурфинкель ВС, Левик ЮС, Козенников ОВ, Селионов ВА (1998) Существует ли генератор шагательных движений у человека? Физиология человека 24 (3): 42–50. [Gurfinkel VS, Levik US, Kozennikov OV, Selionov VA (1998) Is there a locomotor circuitry in humans? Fiziol chelov 24 (3): 42–50. (In Russ)].
Minyaeva AV, Moiseev SA, Pukhov AM, Savokhin AA, Gerasimenko YP, Moshonkina TR (2017) Response of external inspiration to the movements induced by transcutaneous spinal cord stimulation. Human Physiol 43 (5): 43–51. https://doi.org/10.7868/S0131164617050113
Moiseev SA, Ivanov SM, Gorodnichev RM (2022) The Motor Synergies’ Organization Features at Different Levels of Motor Control during High Coordinated Human’s Movement. J Evol Biochem Phys 58: 610–622. https://doi.org/10.1134/S0022093022020272
Moiseev S, Pukhov A, Mikhailova E (2022) Methodological and computational aspects of extracting extensive muscle synergies in moderate-intensity locomotions. J Evol Biochem Phys 58: 88–97. https://doi.org/10.1134/S0022093022010094
Ivanenko Y, Poppele R, Lacquaniti F (2006) Motor control programs and walking. Neuroscientist 12 (4): 339–348. https://doi.org/10.1177/1073858406287987
Hogan N, Sternad D (2013) Dynamic primitives in the control of locomotion. Front Comput Neurosci 7: 71. https://doi.org/10.3389/fncom.2013.00071
De Marchis C, Schmid M, Bibbo D (2013) Inter-individual variability of forces and modular muscle coordination in cycling: a study on untrained subjects. Hum Mov Sci 32 (6): 1480–1494. https://doi.org/10.1016/j.humov.2013.07.018
Шапкова ЕЮ (2004) Локализация и свойства спинальных генераторов локомоций (СЛГ), выявленные электростимуляцией спинного мозга у человека. Рос физиол журн им ИМ Сеченова 90 (8): 420–432. [Shapkova EY (2004) Localization and properties of spinal locomotion generators (SLG) revealed by electrical stimulation of the spinal cord in humans. Russ J Physiol 90 (8): 420–432. (In Russ)].
Rybak I, Shevtsova N, Lafreniere-Roula M (2006) Modelling spinal circuitry involved in locomotor pattern generation: insights from deletions during fictive locomotion. J Physiol 577 (2): 617–639. https://doi.org/10.1113/jphysiol.2006.118703
Churchland M, Cunningham J, Kaufman M (2012) Neural population dynamics during reaching. Nature 487 (7405): 51–56. https://doi.org/10.1038/nature11129
Hogan N, Sternad D (2007) On rhythmic and discrete movements: reflections, definitions and implications for motor control. Exp Brain Res 181 (1): 13–30. https://doi.org/10.1007/s00221-007-0899-y
Sternad D, Marino H, Charles S, Duarte M, Dipietro L, Hogan N (2013) Transitions between discrete and rhythmic primitives in a unimanual task. Front Comput Neurosci 7: 90.https://doi.org/10.3389/fncom.2013.00090
Overduin SA, d’Avella A, Roh J, Carmena JM, Bizzi E (2015) Representation of Muscle Synergies in the Primate Brain. J Neurosci 35 (37): 12615–12624. https://doi.org/10.1523/JNEUROSCI.4302-14.2015
Дополнительные материалы отсутствуют.
Инструменты
Журнал эволюционной биохимии и физиологии