

Известия РАН. Физика атмосферы и океана, 2023, T. 59, № 1, стр. 71-87
Оценка баланса углерода степных экосистем России
Л. Л. Голубятников a, *, И. Н. Курганова b, В. О. Лопес де Гереню b
a Институт физики атмосферы им. А.М. Обухова РАН
119017 Москва, Пыжевский пер., 3, Россия
b Институт физико-химических и биологических проблем почвоведения РАН
142290 Пущино, Институтская ул., 2, Россия
* E-mail: golub@ifaran.ru
Поступила в редакцию 11.07.2022
После доработки 04.10.2022
Принята к публикации 12.10.2022
- EDN: EHIJUF
- DOI: 10.31857/S0002351523010042
Аннотация
Степные экосистемы, занимая около 8% поверхности суши, являются существенным звеном глобального круговорота углерода в системе атмосфера–растительность–почва. На основе геоинформационно-аналитического метода с использованием базы данных эмпирически измеренных значений первичной биологической продукции и регрессионной модели, позволяющей оценить интенсивность поступления диоксида углерода из почвы в атмосферу, получена оценка углеродного (C–СО2) баланса естественных степных экосистем России. Показано, что рассматриваемые степные экосистемы на территории России являются стоком атмосферного углерода. Средняя интенсивность этого потока составляет 231 ± 202 гС/(м2 год). Естественные степные экосистемы России ежегодно депонируют 111 ± 97 МтС. Согласно полученным оценкам, естественные степные экосистемы могут обеспечить от 8 до 19% стока атмосферного углерода в наземные экосистемы России.
ВВЕДЕНИЕ
Экосистемы суши играют ключевую роль в глобальном круговороте углерода, оказывают существенное влияние на концентрацию парниковых газов в атмосфере. Растительный покров наземных экосистем ежегодно ассимилирует 150–175 ГтС атмосферного диоксида углерода (СО2) в результате фотосинтеза [1–3], создавая общую первичную биологическую продукцию растительности (GPP, gross primary production). Около 50–60% абсорбированного растениями углерода возвращается в атмосферу в результате дыхания растений – автотрофного дыхания (RA, autotrophic respiration) [4, 5]. Показателем интенсивности процесса аккумуляции атмосферного углерода растительным покровом является первичная биологическая продукция (NPP, net primary production), которая определяется количеством органического вещества, накапливающимся за определенный интервал времени (обычно это год) на единице площади в надземной и подземной частях растительного сообщества [6], т.е. NPP = GPP-RA. По данным [7, 8], ежегодно растительность наземных экосистем депонирует 29–37% СО2, поступающего в атмосферу в результате хозяйственной деятельности человека. Поток СО2 из наземных экосистем в атмосферу, образующийся в результате разложения микроорганизмами почвенного органического вещества и растительного детрита, называется гетеротрофным дыханием почвы (RH, soil heterotrophic respiration). Гетеротрофное дыхание почвы и дыхание корней растений являются основными естественными источниками СО2, поступающего в атмосферу с поверхности почвы. Этот поток получил название почвенного дыхания (RS, soil respiration) [9, 10]. По данным [11–15], оценки RS для наземных экосистем лежат в пределах 68–100 ГтС/год. Почвенное дыхание является самым интенсивным потоком СО2 из экосистем суши в атмосферу и незначительное изменение его величины в глобальном масштабе может привести к существенному изменению концентрации СО2 в атмосфере [12, 16]. Согласно оценкам [12, 14, 17, 18], глобальное дыхание почв в настоящее время на порядок превышает антропогенные выбросы СО2 в результате сжигания ископаемого топлива.
Функционирование экосистемы в качестве стока или источника атмосферного СО2 определяется знаком величины углеродного (C–СО2) баланса рассматриваемой экосистемы. Значение углеродного баланса природной экосистемы или чистой экосистемной продукции (NEP, net ecosystem production) равно разности между поступлением углерода в экосистему в результате продукционных процессов и эмиссией углерода (в виде CО2) в результате процессов деструкции органического вещества накопленного в экосистеме [16, 19–21]. Значительная неопределенность оценок глобального баланса углерода связана с разной степенью исследования интенсивности потоков углерода как на уровне отдельных экосистем, так и регионов в целом.
Одной из природных зон с достаточно слабо изученными звеньями круговорота углерода является зона степей. Степи относятся к засушливым экосистемам, в которых растительный покров образован преимущественно разными видами трав с доминированием многолетних злаков. Степные экосистемы занимают 8% непокрытой льдом поверхности суши и являются существенным углеродным резервуаром из-за высокого содержания органического вещества в почве, запасы которого в метровом слое почвы составляют 22.9–35.7 кгС/м2 [22–26]. Степную зону образуют степи Евразии, прерии Северной Америки, близкие к прериям травяные сообщества юго-востока Австралии, пампы Южной Америки, вельды Южной Африки, туссоки Новой Зеландии [27]. Самая обширная часть степной зоны расположена на территории Евразии. Евразийские степи занимают около 8 млн. км2 и образуют полосу шириной от 150 до 600 км [28] вытянутую в широтном направлении от Среднедунайской низменности на западе до Маньчжурской равнины на востоке (между 41°–57° N и 26°–127° E).
Для степных экосистем характерен широкий диапазон изменения климатических характеристик, которые определяют зональность и контрастность почвенно-растительного покрова степной зоны [27, 29]. Согласно климатической классификации В. Кёппена [30] в степной зоне представлены степной, пустынный, теплый морской, умеренный морской, теплый континентальный и умеренный континентальный типы климата.
Отличительной чертой растительного покрова степей является мощная корневая система – доля подземной фитомассы может достигать 87% общей фитомассы растений [31]. В степных экосистемах сформировались наиболее плодородные типы почв: чернозёмы и каштановые в степях Евразии, чернозёмовидные в прериях Северной Америки и пампах Южной Америки. Степная территория является основным земледельческим регионом мира и характеризуется высокой степенью антропогенной деградации природных экосистем. Значительная часть территории степей распахана и используется для выращивания сельскохозяйственных культур. Большие степные площади используются в качестве пастбищ и сенокосов [29].
Почвенно-растительный покров, климатические условия, интенсивность антропогенного влияния определяют основные параметры углеродного цикла степных экосистем и их способность к поглощению и эмиссии парниковых газов. Из всех типов наземных экосистем на территории России роль степных экосистем в биогенном цикле углерода наименее определена. В настоящей работе для степных экосистем России получены оценки углеродного (C–СО2) баланса и его компонентов (NPP, RH), определен вклад этих экосистем в углеродный бюджет страны.
ОБЪЕКТЫ ИССЛЕДОВАНИЯ
Согласно карте растительности [32], степные экосистемы на территории России занимают около 1.7 млн. км2, т.е. менее 10% территории страны. Степная зона в России охватывает обширную территорию южной части Восточно-Европейской равнины, равнинные территории Предкавказья и Крыма, южные районы Западно-Сибирской равнины, межгорные котловины северной части Центральной Азии (республик Бурятия, Хакассия, Тыва, Иркутской области, Забайкальского края и южной части Красноярского края) (рис. 1). Степные экосистемы России подразделяются на луговые, настоящие, сухие и опустыненные. Подзона луговых степей расположена вдоль южной границы лесной зоны. На территории России луговые степи образуют наиболее обширную по площади степную подзону (710 тыс. км2). Под экосистемами настоящих степей в России находится 470 тыс. км2. Эти экосистемы занимают значительные территории на юго-западе Европейской части России, в Азиатской части встречаются фрагментарно. Сухие степи распространены преимущественно в Ставропольском крае, республиках Крым, Дагестан, Тыва. Площадь этой степной подзоны на территории России составляет 320 тыс. км2. Экосистемы опустыненных степей распространены на площади 230 тыс. км2, занимают значительную территорию на юго-востоке Европейской части России и в межгорных котловинах центральной и южной Тывы.
Рис. 1.
Картосхема размещения степей разных типов на территории России. Типы степей: I – луговые, II – настоящие, III – сухие, IV – опустыненные.
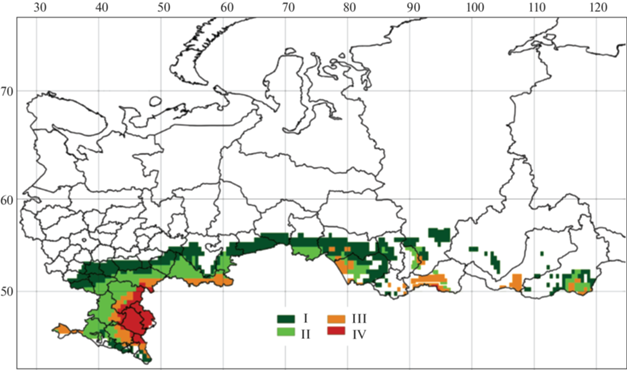
В степных экосистемах наблюдается дефицит влаги, который нарастает от луговых степей к опустыненным степям. Большая часть атмосферных осадков (75–85%) на степной территории России выпадает в теплый период года. Годовое количество осадков изменяется от 430 мм в луговых степях до 150 мм в опустыненных. Испаряемость с открытой водной поверхности составляет от 650 мм/год в луговых степях до 800 мм/год в опустыненных. В степном регионе России годовые суммы температур приземного воздуха >10°С находятся в диапазоне от 1900 до 2600°С [27].
Значительную часть степной территории России занимают сельскохозяйственные угодья (пашни, пастбища, сенокосы) [33]. В степной зоне расположены полностью или частично территории сорока двух субъектов Российской Федерации (РФ). Природные степные экосистемы сохранились на относительно небольших участках. Согласно инвентаризации сохранившихся степных массивов [34], общая площадь природных степных экосистем на территории России оценивается в 200 тыс. км2, что составляет менее 12% общей площади степной территории РФ.
В 90-х годах XX века из-за произошедших в России социально-экономических изменений значительные площади пахотных земель были выведены из сельскохозяйственного оборота. Необрабатываемые пахотные угодья пополнили пул залежных земель. Из-за сокращения поголовья скота снизилась антропогенная нагрузка на пастбища, сократилась площадь пастбищ и сенокосов. В результате этих событий на части сельскохозяйственных угодий в степной зоне начался процесс восстановления природных степных экосистем. При благоприятных условиях восстановления за 15–30 лет в этих экосистемах формируется травянистый покров, который по видовому составу близок к степным сообществам, существовавшим до распашки [35, 36]. В процессе формирования степного растительного покрова возрастает количество поступающего в почву органического вещества и происходит постепенное восстановление естественных физико-химических свойств и микробиологических характеристик почв бывших сельскохозяйственных угодий [37]. В частности, в почвенных горизонтах залежных земель изменяется структура, плотность, водно-воздушный и гидротермический режимы, возрастает содержание углерода и элементов минерального питания растений [38–40]. В данной работе рассматриваются природные и восстанавливающиеся (вторичные) степные экосистемы. Эти экосистемы объединены общим названием естественные экосистемы степной зоны.
В течение вегетационного периода в природных степных экосистемах интенсивность аккумуляции атмосферного углерода в процессе фотосинтеза превосходит интенсивность выделения углерода в атмосферу в результате деструкционных процессов [16, 41]. В холодный период года в результате увядания и отмирания надземной части травянистого покрова и формирования снежного покрова поглощение атмосферного углерода растительностью степных экосистем прекращается, в то же время продолжается разложение почвенного детрита микроорганизмами и дыхание подземных частей растений, т.е. продолжается поступление CО2 из почв этих экосистем в атмосферу. Следует отметить, что в холодный период года в результате низких температур воздуха и промерзания верхних слоев почвы интенсивность деструкционных процессов в степных экосистемах значительно уменьшается по сравнению с теплым периодом. Таким образом, в теплый период года природные степные экосистемы являются стоком атмосферного углерода, в холодный период года эти экосистемы функционируют как источник поступления СО2 в атмосферу. Поглощаемый степными растениями атмосферный углерод накапливается, в основном, в органическом веществе почвенного пула экосистем (корневой системе растений, фитодетрите, гумусе) [14, 42]. По данным [31, 36, 42, 43], подземная корневая часть растений в степных экосистемах содержит 80–95% углерода общей фитомассы растений.
МЕТОДЫ ИССЛЕДОВАНИЯ
Площади вторичных степных экосистем рассчитаны на основе данных статистических отчетов о наличии и распределении земель в субъектах РФ на 1 января 2020 года Федеральной службы государственной регистрации, кадастра и картографии (Росреестр) и бюллетеней о состоянии сельского хозяйства РФ в 2020 году Федеральной службы государственной статистики (Росстат). В данных по земельной статистике Росреестра [44] к группе залежей сельскохозяйственных угодий субъектов РФ отнесены земельные участки, которые официально переведены из пашни в залежи. Однако часть необрабатываемых пахотных земель в субъектах РФ не переводилась в другие угодья и формально продолжает считаться пашней. По этой причине оценка реальной площади залежей выполнена на основе статистических данных Росреестра о площадях пахотных угодий и декларируемых залежей [44] с учетом данных ежегодных отчетов Росстата о посевных площадях и площадях чистых паров [45]. Восстановление степных экосистем происходит и на территориях, которые перестали использоваться как пастбища и сенокосы. Оценка площадей вторичных степей на месте бывших пастбищ и сенокосов сделана на основе статистических данных Росреестра о площадях этих типов сельскохозяйственных угодий [44] и данных ежегодных отчетов Росстата о площадях сенокосов и пастбищ, используемых хозяйствами и гражданами [45].
В ряде рассматриваемых субъектов РФ заброшенные пахотные земли, неиспользуемые пастбища и сенокосы расположены не только в степной зоне. Данные о площадях участков сельскохозяйственных угодий, на которых прекращено антропогенное влияние и которые находятся вне степной зоны, отсутствуют. В качестве верхней оценки для площадей указанных участков в работе [35] предложено принять 20% территории, которая перестала использоваться в качестве сельскохозяйственных угодий в рассматриваемом субъекте РФ.
Оценки первичной биологической продукции естественных степных экосистем в субъектах РФ получены на основе геоинформационно-аналитического метода с использованием базы данных эмпирически измеренных значений этой величины. В данном исследовании использованы данные о NPP степных экосистем из работ [28, 31, 32, 36, 43, 46–52]. Для перевода значений NPP, выраженных в единицах сухого вещества, в углеродные единицы использованы коэффициенты, полученные на основе аналитических данных по содержанию азота (N) и соотношению С/N в фитомассе рассматриваемых типов экосистем [53].
Оценки почвенного дыхания выполнены на основе регрессионной T&P-модели [54]. Эта модель позволяет оценивать среднемесячную интенсивность выделения CO2 из почв (RSd, гС/(м2 сут)) на основе среднемесячной приземной температуры воздуха (T, °C) и количества осадков за месяц (P, см/мес):
где F (гC/(м2 сут)) – почвенное дыхание при 0°C и достаточном увлажнении почвы; Q (° 1/C) – коэффициент температурной чувствительности почвенного дыхания, К (см/мес) – константа полунасыщения гиперболической зависимости между RSd и P. Расчет параметров для T&P-модели выполнен нелинейным методом наименьших квадратов в программной среде для статистической обработки данных R (функция nlm) [55]. Месячные потоки CО2 из почв степных экосистем получены умножением значений RSd на количество дней соответствующего месяца. Годовой поток CО2 из почв экосистем RS вычислен суммированием расчетных значений месячных величин почвенного дыхания. Расчет почвенного дыхания степных экосистем выполнен на основе метеорологических данных за период с 1970 по 2000 годы из базы WorldClim (www.worldclim.org).На основе экспериментальных данных для разных природно-климатических зон установлено, что доля гетеротрофного дыхания в общем потоке CО2 из почв травяных экосистем составляет 55% [56], т.е. для значений RH и RS справедливо соотношение
Углеродный (C–СО2) баланс естественных степных экосистем рассчитан по формуле:
Если величина NEP для экосистемы положительная, то рассматриваемая экосистема является стоком атмосферного углерода. В случае если величина NEP отрицательная, то экосистема представляет собой источник атмосферного углерода.Для оценки углеродного баланса естественных степных экосистем в теплый и холодный периоды года полагалось, что теплый период года составляют месяцы с T ≥ +1°C, холодный период – месяцы с T < +1°C. Продолжительность теплого и холодного периодов года в степных районах определялась для каждого из рассматриваемых субъектов РФ.
Используемые данные земельной статистики Росреестра и Росстата сгруппированы по субъектам РФ. По этой причине площади степных экосистем и значения потоков NPP, RH, NEP для этих экосистем рассчитаны для каждого субъекта РФ, имеющего степные территории. Расчеты выполнены на пространственной сетке с разрешением 0.5° × 0.5° географической широты и долготы. На основе полученных результатов определены средние значения NPP, RH, NEP для естественных степных экосистем как для России в целом, так и для Европейского, Сибирского и Центральноазиатского регионов РФ. Расчеты средних значений указанных потоков выполнены с учетом площадей занимаемых естественными степными экосистемами в каждом субъекте РФ.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Среднегодовые приповерхностные температуры воздуха и годовые количества атмосферных осадков. Для степной территории России характерно существенное изменение климатических показателей как в широтном, так и в долготном направлениях. Следует отметить, что в степном регионе России диапазоны изменения климатических характеристик значительно превосходят аналогичные диапазоны свойственные лесному региону умеренных широт, который занимает более обширную территорию в РФ по сравнению со степями [27].
Согласно используемым метеорологическим данным значения среднегодовой приповерхностной температуры воздуха (Tyr) для степных территорий субъектов РФ изменяются от –5.0…–4.8°С (республики Бурятия и Тыва) до 10.5–10.9°С (Краснодарский край, Республика Крым). Значения годовых осадков (Pyr) в степях России варьируют от 237–284 мм/год (Астраханская область, Республика Калмыкия) до 765–769 мм/год (Краснодарский край, Карачаево-Черкесская Республика). Наибольший размах значений Tyr (8.4°С) и Pyr (532 мм/год) наблюдается в степных экосистемах Европейского региона России. Статистические характеристики величин Tyr и Pyr для степных регионов РФ представлены в табл. 1.
Таблица 1.
Статистические характеристики среднегодовой приповерхностной температуры воздуха (Tyr) и годового количества осадков (Pyr) для степных регионов России
| Степной регион | Тyr, °С | Рyr, мм/год | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| xmin | xmax | A | $\bar {x}$ | σ | Cv, % | xmin | xmax | A | $\bar {x}$ | σ | Cv, % | |
| Европейский | 2.5 | 10.9 | 8.4 | 7.3 | 2.5 | 34 | 237 | 769 | 532 | 420 | 121 | 29 |
| Сибирский | –2.9 | 2.1 | 5.1 | 0.9 | 1.6 | 184 | 395 | 578 | 183 | 444 | 48 | 11 |
| Центральноазиатский | –5.0 | –0.6 | 4.4 | –3.6 | 1.8 | 50 | 372 | 583 | 211 | 437 | 71 | 16 |
| Степные регионы России в целом | –5.0 | 10.9 | 15.9 | 3.1 | 5.2 | 170 | 237 | 769 | 532 | 430 | 98 | 23 |
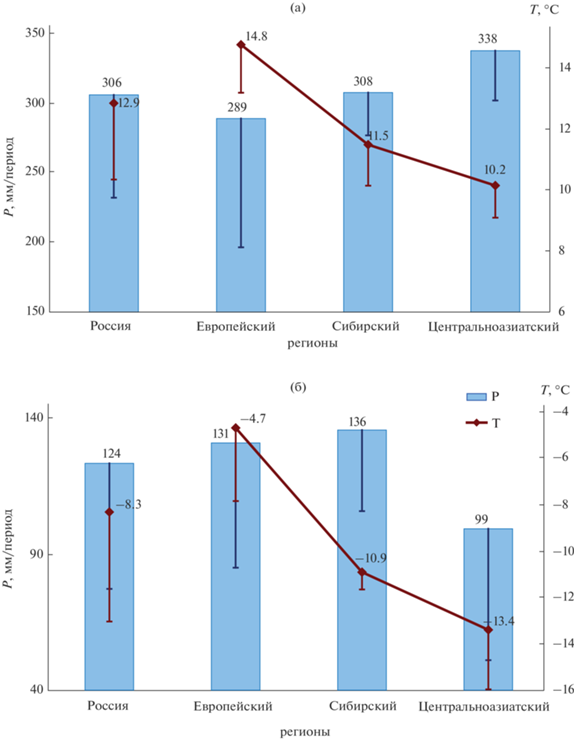
Средние приповерхностные температуры воздуха теплого/холодного периода года в степных экосистемах России изменяются от 10.2°С/–13.4°С в Центральноазиатском регионе до 14.8°С/–4.7°С в Европейском регионе (рис. 2). Количество атмосферных осадков теплого периода года в степных регионах России в 2.2–3.4 раза превосходит количество осадков холодного периода. В степях Центральноазиатского региона по сравнению с другими степными регионами России выпадает наибольшее (338 мм/период) количество осадков в теплый период года (рис. 2а) и наименьшее (99 мм/период) количество осадков в холодный период года (рис. 2б).
Рис. 2.
Приповерхностная температура воздуха (T) и количество осадков (P) в степных регионах России (средние значения за период и стандартные отклонения): а – теплый период года, б – холодный период года. Вертикальными линиями изображены отрицательные части стандартных отклонений.
Оценка площадей природных и вторичных степных экосистем. В [34] на основе полевых исследований и космических снимков среднего разрешения были выявлены сохранившиеся относительно крупные массивы природных степных экосистем на территории России. Значительная часть (131 тыс. км2) сохранившихся степных экосистем находится в Европейском регионе. В Сибирском и Центральноазиатском регионах на до-лю сохранившихся степей приходится 18 и 53 тыс. км2, соответственно (рис. 3 а).
Рис. 3.
Распределение площадей природных и вторичных степных экосистем в регионах РФ и на территории России в целом. Степные экосистемы: I – на необрабатываемых пашнях (залежные), II – на неиспользуемых пастбищах и сенокосах, III – природные.
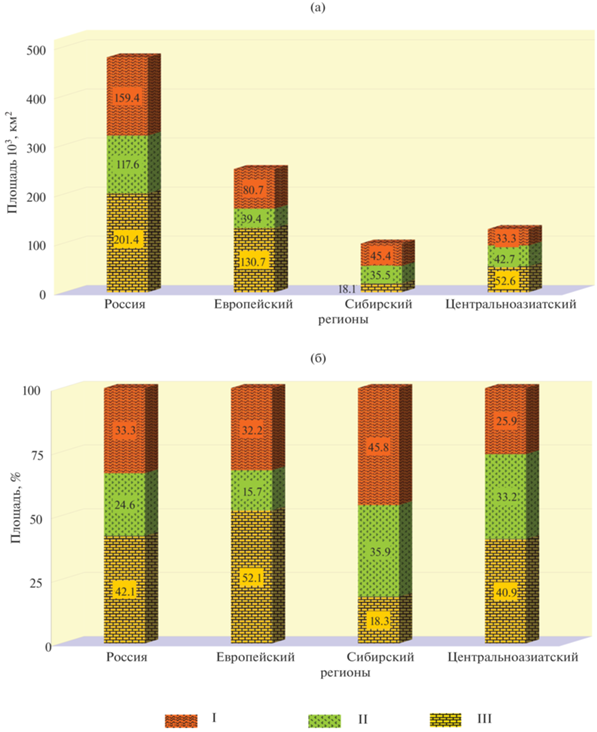
После прекращения использования некоторых сельскохозяйственных угодий в степной зоне на площадях бывших пахотных земель, пастбищ и сенокосов начался процесс восстановления степных экосистем. Согласно выполненным расчетам, на начало 2020 г. вторичные степные экосистемы в России занимали 277 тыс. км2, т.е. их площадь на 38% превышала площадь природных степных участков.
Большая часть (57%) вторичных степных экосистем расположена в Азиатской части России. Площади степных экосистем, которые появились на месте заброшенных пахотных земель, пастбищ и сенокосов, в Азиатской и Европейской частях России практически одинаковы и составляют около 80 тыс. км2 (рис. 3а). Площадь вторичных степных экосистем, которые появились на месте неиспользуемых пастбищ и сенокосов, в Азиатской части России в 2 раза превосходит площадь аналогичных экосистем в Европейской части России (рис. 3 б).
Идентификация T&P-модели. Многолетние инструментальные исследования внутригодовой динамики эмиссии CO2 из почв естественных степных экосистем в России не проводились. По этой причине идентификация T&P-модели (1) была выполнена на основе данных многолетних полевых измерений потоков CО2 в атмосферу из почв двух луговых экосистем, расположенных в южной части Московской области. Еженедельные измерения эмиссии CО2 из почвы проводились круглогодично с 1998 по 2017 года методом закрытых камер на злаково-разнотравном лугу Приокско-Террасного биосферного заповедника и с 2004 по 2018 года на разнотравно-злаковом лугу вблизи г. Пущино. При измерениях фиксировался поток CО2 в атмосферу из луговых почв, который включал в себя CО2, образовавшейся как в результате дыхания корней растений, так и разложения микроорганизмами напочвенного детрита и органического вещества почв. Луговые экосистемы различались по видовому составу растительности, типовой принадлежности и текстуре почв. Для рассматриваемых луговых экосистем характерны значительное видовое разнообразие травянистого покрова, сформированная корнями злаков мощная дернина, значения фитомассы и NPP близкие к аналогичным показателям луговых степей. По этим причинам, с некоторыми допущениями, рассматриваемые луговые экосистемы можно рассматривать как экосистемы близкие к экосистемам луговых степей. В расчетах использовались метеорологические данные станции фонового мониторинга (пос. Данки, Серпуховской район, Московская область), которая расположена на одной из площадок экспериментальных исследований почвенного дыхания.
На основе экспериментальных данных для каждой из рассматриваемых луговых экосистем определены модельные параметры F, Q и K (табл. 2). Проведенные расчеты показали, что полученные две T&P-модели (T&P-G1, T&P-G2) адекватно описывают внутригодовую динамику среднемесячной эмиссии СО2 из почв исследуемых экосистем (табл. 3). Коэффициенты несоответствия Тейла (U) модельных рядов среднемесячных значений почвенного дыхания и данных инструментальных измерений этой величины не превышают значения 0.3 (табл. 2), которое является пороговым значением для природных процессов [57, 58]. Полученные значения коэффициента U свидетельствует в пользу того, что результаты моделирования имеют высокую степень совпадения с данными инструментальных измерений. Значения коэффициента детерминации R2 (табл. 2) также указывают на высокую корреляционную связь расчетных и экспериментальных значений среднемесячных потоков СО2 в атмосферу из почв рассматриваемых экосистем.
Таблица 2.
Параметры и оценки точности T&P-моделей для расчета потоков С–СО2 из почв луговых экосистем
| Объект исследования | Параметры модели | Оценки точности модели | ||||||
|---|---|---|---|---|---|---|---|---|
| модель | F, гС/(м2 сут) |
Q, ° 1/C | К, см/мес | R2 | U | |||
| T&P-G1 | T&P-G2 | T&P-G1 | T&P-G2 | |||||
| Экосистема 1 | T&P-G1 | 1.687 | 0.0569 | 2.203 | 0.62 | 0.64 | 0.19 | 0.24 |
| Экосистема 2 | T&P-G2 | 1.843 | 0.0654 | 2.745 | 0.71 | 0.70 | 0.20 | 0.17 |
Таблица 3.
Средние значения ($\bar {x}$) и стандартные отклонения (σ) экспериментальных (FieldData) и модельных (T&P-G1, T&P-G2) значений почвенного дыхания (гС/(м2‧сут)) для двух луговых экосистем
| Месяц | Экосистема 1 | Экосистема 2 | ||||||
|---|---|---|---|---|---|---|---|---|
| FieldData | T&P-G1 | FieldData | T&P-G2 | |||||
| $\bar {x}$ | σ | $\bar {x}$ | σ | $\bar {x}$ | σ | $\bar {x}$ | σ | |
| Январь | 0.66 | 0.41 | 0.72 | 0.20 | 0.63 | 0.33 | 0.67 | 0.24 |
| Февраль | 0.74 | 0.48 | 0.73 | 0.22 | 0.51 | 0.27 | 0.67 | 0.21 |
| Март | 0.74 | 0.42 | 0.97 | 0.21 | 0.55 | 0.26 | 0.97 | 0.26 |
| Апрель | 1.34 | 0.57 | 1.50 | 0.35 | 1.46 | 0.58 | 1.57 | 0.47 |
| Май | 2.69 | 0.88 | 2.46 | 0.53 | 3.32 | 0.89 | 3.05 | 0.53 |
| Июнь | 3.54 | 1.23 | 3.18 | 0.43 | 4.73 | 1.59 | 3.78 | 0.63 |
| Июль | 3.86 | 1.52 | 3.77 | 0.60 | 4.45 | 1.64 | 4.40 | 0.80 |
| Август | 3.00 | 1.44 | 3.19 | 0.88 | 3.55 | 1.46 | 4.04 | 1.01 |
| Сентябрь | 2.44 | 0.88 | 2.26 | 0.38 | 2.82 | 1.03 | 2.46 | 0.52 |
| Октябрь | 1.61 | 0.65 | 1.62 | 0.33 | 1.69 | 0.65 | 1.73 | 0.45 |
| Ноябрь | 0.96 | 0.47 | 1.05 | 0.22 | 1.13 | 0.34 | 1.10 | 0.26 |
На основе модельных и экспериментальных данных о месячных потоках СО2 в атмосферу из почв рассматриваемых луговых экосистем оценены годовые величины RS этих экосистем. Полученные результаты показали, что отклонение модельных значений годового RS от экспериментальных оценок этой величины в луговых экосистемах лежит в диапазоне от 1 до 26% в зависимости от используемой T&P-модели. В тоже время отклонение средних значений годового RS, полученных на основе модельных оценок этого потока в каждой луговой экосистеме, от годовых экспериментальных значений RS составляет 6–11%. В рамках настоящего исследования значение годового потока СО2 в атмосферу из почв степных экосистем рассчитано как среднее значение соответствующих годовых потоков СО2 рассчитанных по данным моделей T&P-G1, T&P-G2.
Поглощение атмосферного углерода растительным покровом степных экосистем. Величина первичной биологической продукции степных экосистем снижается от луговых степей к опустыненным, подчиняясь фактору аридности [31, 47]. Для луговых степей России величина NPP составляет 755 ± 64 гС/(м2 год) (среднее ± стандартное отклонение) В рассматриваемых опустыненных степях значение NPP уменьшается до 311 ± ± 36 гС/(м2 год). Средняя величина NPP для естественных экосистем степных регионов России составляет 594 ± 148 гС/(м2 год). Следует отметить, что полученное среднее значение NPP для изучаемых степных экосистем в 1.9 раз превышает среднее значение NPP для лесных экосистем на территории России, оцененное в 319 ± 19 гС/(м2 год) [59].
Согласно полученным оценкам значения NPP естественных степей Европейского региона России изменяется от 282 до 779 гС/(м2 год) со средним значением 494 гС/(м2 год). В степных экосистемах Сибирского региона значения NPP варьируют от 656 до 790 гС/(м2 год) со средним значением 730 гС/(м2 год). Первичная продуктивность степных растительных сообществ межгорных котловин Центральноазиатского региона России изменяется от 435 до 741 гС/(м2 год) со средним значением 613 гС/(м2 год). Статистические характеристики значений NPP рассматриваемых регионов России приведены в табл. 4. Полученные оценки показывают, что на территории Европейского региона России продуктивность естественных степей незначительно превышает продуктивность лесов (454 гС/(м2·год) [59]) этого региона. Средняя продуктивность степных экосистем Сибирского региона в 1.5–1.8 раза выше продуктивности лесных экосистем южной тайги Центральной Сибири (405–475 гС/(м2 год) [60]).
Таблица 4.
Статистические характеристики значений первичной биологической продукции, гетеротрофного дыхания почвы и углеродного баланса в степных регионах России
| Степной регион | xmin | xmax | A | $\bar {x}$ | σ | Сv, % |
|---|---|---|---|---|---|---|
| Первичная биологическая продукция, NPP | ||||||
| Европейский | 282 | 779 | 497 | 494 | 160 | 32 |
| Сибирский | 656 | 790 | 134 | 730 | 39 | 5 |
| Центральноазиатский | 435 | 741 | 306 | 613 | 108 | 18 |
| Степные регионы России в целом | 282 | 790 | 509 | 575 | 160 | 28 |
| Почвенное гетеротрофное дыхание, RH | ||||||
| Европейский | 348 | 554 | 206 | 405 | 43 | 11 |
| Сибирский | 240 | 337 | 98 | 312 | 30 | 10 |
| Центральноазиатский | 216 | 296 | 79 | 247 | 26 | 11 |
| Степные регионы России в целом | 216 | 554 | 338 | 343 | 78 | 23 |
| С–СО2 баланс, NEP | ||||||
| Европейский | –134 | 394 | 527 | 89 | 173 | 195 |
| Сибирский | 370 | 463 | 93 | 418 | 35 | 8 |
| Центральноазиатский | 218 | 465 | 247 | 366 | 93 | 25 |
| Степные регионы России в целом | –134 | 465 | 599 | 231 | 202 | 87 |
Согласно полученным результатам растительный покров естественных степных экосистем Европейского региона России депонируют 124 ± 5 МтС/год, Центральноазиатского региона – 79 ± 6 МтС/год, Сибирского региона – 72 ± ± 4 МтС/год. Полученные оценки депонирования атмосферного углерода естественными степными экосистемами регионов России составляют 45, 29 и 26% суммарной NPP степей РФ соответственно.
Поступление C–СО2 в атмосферу из почв степных экосистем. Согласно расчетам средний годовой поток двуокиси углерода из почв степной зоны на территории России составляет 624 ± ± 142 гС/(м2 год). Полученная оценка годового почвенного дыхания степных экосистем сравнима со значениями RS (319–735 гС/(м2 год) [61–63]), которые наблюдались в лесных экосистемах умеренных широт России. Немногочисленные эмпирические оценки интенсивности поступления СО2 в атмосферу из степных почв на территории России основаны на данных нескольких измерений, проведенных в течение одного–двух вегетационных сезонов. Согласно опубликованным данным инструментальных измерений потоки СО2 в атмосферу из почв отдельных естественных степных экосистем в России фиксировались в диапазоне от 194 до 3269 гС/(м2 год) [64, 65]. Существенный разброс величин RS для степных экосистем предопределен в первую очередь разнообразием почвенного и растительного покрова степей, погодными условиями в период проведения исследования, различиями в методах, которые использовались для измерения почвенного дыхания [66].
Опубликованные оценки вклада RH в эмиссию СО2 с поверхности почв травяных экосистем превышают 50%. Согласно [67] модельные оценки доли гетеротрофного дыхания в почвенном дыхании степных экосистем России изменяются от 59 до 71% со средним значением в 68%. В [68, 69] сообщается, что для североамериканских прерий соотношение RH/RS составляет 60–75%. В [70] вклад RH в почвенное дыхание травяных экосистем центральной Италии оценен в 64–72%. Используемая в данной работе статистическая оценка доли RH в почвенном дыхании травяных экосистем (55%) близка к нижней границе приведенного в [71] интервала изменения этой величины для травяных экосистем умеренных широт (58–72%) и согласуется с аналогичной оценкой (52%) полученной на основе инструментальных измерений на луговых и степных участках Центрально-Черноземного биосферного заповедника (Курская область, Россия) [72].
Согласно полученным оценкам, почвенное гетеротрофное дыхание естественных степей Европейского региона России изменяется от 348 до 554 гС/(м2 год) со средним значением 405 гС/(м2 год). В степных экосистемах Сибирского региона значения RH варьируют от 240 до 337 гС/(м2 год) со средним значением 312 гС/(м2 год). Для степных экосистем межгорных котловин Центральноазиатского региона России получены значения RH от 216 до 296 гС/(м2 год) со средним значением 247 гС/(м2 год). Среднее значение RH естественных степных экосистем на территории России оценено в 343 ± 78 гС/(м2 год). Статистические характеристики значений RH рассматриваемых регионов приведены в табл. 4. Полученные модельные оценки гетеротрофного дыхания естественных степных экосистем превышают аналогичные оценки для степей Европейской части России (250–285 гС/(м2 год)) и сравнимы с оценками для степей Азиатской части России (219–246 гС/(м2 год)), которые приведены в [67]. Оценка среднего значения RH естественных степных экосистем России согласуется с полученной в [73] оценкой этой величины – 390 гС/(м2 год).
Выполненные расчеты показали, что в результате гетеротрофного дыхания из почв естественных степных экосистем России в атмосферу ежегодно поступает 164 ± 37 МтС. Почвы рассматриваемых степных экосистем Европейского региона в результате гетеротрофного дыхания эмитируют 102 ± 11 МтС/год или 62% годового количества RH степных регионов России. Для Сибирского и Центральноазиатского регионов величины RH естественных степных экосистем равны 31 ± 3 и 32 ± 3 МтС/год, соответственно, что составляет по 19% годового количества гетеротрофного дыхания почв степных регионов России. Согласно полученным оценкам, годовое гетеротрофное дыхание естественных степных экосистем России составляет 60% годовой NPP этих экосистем.
Оценка C–СО2 баланса в естественных степных экосистемах. В зависимости от периода года степные экосистемы могут быть как источниками, так и стоками атмосферного углерода. Следует отметить, что интенсивность продукционного процесса в степных экосистемах в течение вегетационного периода определяет знак годового углеродного баланса этих экосистем. Инструментальные измерения потоков углерода в экосистемах северных районов Казахстана показали, что природные степи и залежи в вегетационный период депонируют атмосферный углерод (170 и 301 гС/м2 соответственно), в холодный период года из этих экосистем происходит эмиссия СО2 с интенсивностями 128 и 155 гС/м2 соответственно [74]. Разнотравная прерия (Северная Дакота, США) в теплый период года аккумулирует углерод из атмосферы с интенсивностью 129 гС/м2, в холодный период эмитирует углерод в атмосферу с интенсивностью 84 гС/м2 [75]. Согласно проведенным расчетам, в течение теплого периода года природные степи разных типов, разновозрастные залежи и неиспользуемые пастбища Европейского, Сибирского и Центральноазиатского регионов России депонируют 185 ± 168 гС/м2, 501 ± 58 гС/м2 и 438 ± 113 гС/м2 соответственно, в холодный период года из степных экосистем этих регионов в атмосферу поступает 96 ± 21 гС/м2, 83 ± 21 гС/м2, 72 ± 26 гС/м2 соответственно. Высокая углерододепонирующая способность естественных степей Сибирского и Центральноазиатского регионов в теплый период года определяется тем, что в этих регионах, соответственно, 82 и 59% рассматриваемой территории (рис. 3б) занимают вторичные степные экосистемы, т.е. экосистемы активно депонирующие атмосферный углерод. Следует отметить так же, что растительный покров степных экосистем Сибирского региона отличает высокая продуктивность: значения NPP в 1.5 раз превышают продуктивность степей Европейского региона и в 1.2 раза выше продуктивности степей Центральноазиатского региона (табл. 4).
Годовой углеродный баланс степных экосистем, как экосистем аридных регионов, существенно зависит от количества осадков за год и сезонного распределения их выпадения. В годы с режимом выпадения атмосферных осадков благоприятным для развития травянистого покрова естественные степные экосистемы могут быть значительным стоком атмосферного углерода с диапазоном изменения значений NEP от 184 до 236 гС/(м2 год) [76–78]. В засушливые годы степные экосистемы могут как поддерживать свой углеродный баланс в состоянии близком к равновесному со значениями NEP от –32 до 45 гС/(м2 год), так и быть источниками поступление СО2 в атмосферу с варьированием NEP в пределах от –253 до ‒187 гС/(м2 год) [75, 76]. Согласно проведенным расчетам, наиболее интенсивный сток углерода (461–463 гС/(м2 год)) характерен для естественных экосистем луговых степей Сибирского региона России. Климатические условия, в которых функционируют эти экосистемы, благоприятны для развития травянистого покрова в течение вегетационного периода (NPP достигает значений 741–772 гС/(м2 год)). Вместе с тем относительно суровые зимы в этом регионе существенно замедляют гетеротрофное дыхание почв в холодный период года, величина которого варьирует в зимний период от 50 до 120 гС/м2. Выявлено, что некоторые экосистемы сухих и опустыненных степей юго-востока Европейского региона России имеют отрицательные значения NEP в диапазоне от –134 гС/(м2 год) до –35 гС/(м2 год). Эти экосистемы функционируют в неблагоприятных для развития травянистого покрова климатических условиях. В указанных экосистемах интенсивность гетеротрофного почвенного дыхания (RH = 360–486 гС/(м2 год)) превышает первичную биологическую продукцию (NPP = 282–386 гС/(м2 год)).
Антропогенные воздействия на степные экосистемы (выпас животных, распашка, скашивание травостоя, внесение удобрений, весенний пал) существенно влияют на углеродный баланс этих экосистем. Инструментальные измерения потоков углерода на экспериментальных участках настоящей степи северного Казахстана [74] показали, что в агроэкосистемах (поля с зерновыми культурами, сенокосы) значения углеродного баланса уменьшаются по сравнению с участками естественных степных экосистем на 30–60% за вегетационный период и на 10–11% за холодный период года. Из опубликованных результатов исследований следует, что естественные степи северного Казахстана являются стоком атмосферного углерода с интенсивностью 42 гС/(м2 год), а зерновые поля и сеяные сенокосы этого региона выступают источниками атмосферного углерода с интенсивностью 4–52 гС/(м2 год) [74]. Результаты экспериментальных измерений потоков углерода в естественной разнотравной прерии и двух типов пастбищ на её территории [75, 79] показали, что участки прерии без антропогенного влияния являются стоком атмосферного углерода с интенсивностью 46 гС/(м2 год), на пастбищных участках прерии величина стока углерода уменьшается более чем на 20% (табл. 5). Проведенные расчеты показали, что заброшенные пастбища в сухих степях юго-востока Европейского региона России, которые многие годы находились под сильной пастбищной нагрузкой и подверглись значительной деградации, являются источниками поступления СО2 в атмосферу со значениями NEP от –125 до –65 гС/(м2 год).
Таблица 5.
Оценки С–СО2 баланса в степных экосистемах различных регионов мира по данным инструментальных измерений
| Месторасположение, тип степи, экосистема, годы измерений | С–СО2 баланс, гС/(м2 год) | Источник |
|---|---|---|
| Россия, Хакассия, настоящая степь: | [78] | |
| природная степь, 2002–2004 | 99–130 | |
| залежь 5 лет, 2003–2004 | 216 | |
| залежь 10 лет, 2004 | 143 | |
| Северный Казахстан, настоящая степь: | [74] | |
| природная степь, 2002 | 42 | |
| залежь, 2002 | 146 | |
| зерновое поле, 2002 | –52 | |
| сенокос сеяный, 2002 | –4 | |
| Монголия, настоящая степь, пастбище, 2002–2003 | 41 | [90] |
| Венгрия, сухая степь, природная степь, 2003–2004 | –80–180 | [91] |
| США, Северная Дакота, разнотравная прерия: | ||
| природная прерия, 1996–1999 | 45 | [75] |
| пастбище, 1996–1998 | –19–52 (36) | [79] |
| пастбище сеяное, 1996–1998 | –36–35 (–14) | [79] |
| США, Южная Дакота, разнотравная прерия, зерновое поле (кукуруза), 2008–2011 | 121–374 (230) | [92] |
| США, Оклахома, разнотравная прерия: | ||
| зимнее пастбище, 1997 | 142 | [93] |
| зерновое поле (озимая пшеница), 2003–2007 | –74–128 (37) | [92] |
| США, Оклахома, высокотравная прерия: | ||
| природная прерия, 1997–1998 | 268 | [94] |
| природная прерия, после весеннего пала, 1997 | 343 | [93] |
| США, Висконсин, высокотравная прерия: | [25] | |
| природная прерия, 2001–2004 | –144–194 (38) | |
| залежь 65 лет, 2001–2004 | –228–129 (–73) | |
| США, Колорадо, низкотравная прерия: | [76] | |
| природная прерия, 2000–2003 | –49–95 (3) | |
| природная прерия, 2004–2007 | –187–78 (–69) | |
| пастбище с умеренной нагрузкой, 2004–2006 | –32–128 (73) | |
| пастбище с чрезмерной нагрузкой, 2004–2006 | –26–57 (29) | |
| пастбище с умеренной нагрузкой весной и осенью, 2004–2006 |
–102…–61 (–85) | |
| США, Иллинойс, высокотравная прерия, зерновое поле (кукуруза), 1997, 1999, 2001, 2003, 2006 | 295–575 (412) | [92] |
| США, Техас, низкотравная степь, пастбище, 2010–2011 | –353–70 | [76] |
| США, Вайоминг, низкотравная степь, природная степь, 1997–1998 | 39–109 | [76] |
| Канада, Альберта, разнотравная прерия, зерновое поле (яровая пшеница), 2007 | –37 | [92] |
| Канада, Манитоба, высокотравная прерия, зерновое поле (яровая пшеница), 2008 | 229 | [92] |
Заброшенные пашни (залежи) в степной зоне, на которых происходит восстановление степных экосистем, являются существенным стоком атмосферного углерода. Согласно [74] интенсивность годового стока атмосферного углерода в залежную экосистему настоящей степи в 3.5 раза превышает скорость депонирования углерода в экосистеме природной степи (табл. 5). В [78] отмечается, что сток углерода в естественную степную экосистему на территории Хакассии не превышает 130 гС/(м2 год), в то же время залежи 5 и 10 лет абсорбируют 216 и 143 гС/(м2 год) соответственно. Учитывая статистические данные о распределении площадей разных типов естественных экосистем в степной зоне на территории России (рис. 3 ), можно предположить, что около 50% ежегодного стока атмосферного углерода в эти экосистемы приходится на залежные земли.
Согласно полученным оценкам среднее значение NEP в степных экосистемах России составляет 231 ± 202 гС/(м2 год), варьируя от 89 ± 173 гС/(м2 год) в Европейском регионе до 418 ± 35 гС/(м2 год) в Сибирском регионе (табл. 3). Для естественных степных экосистем Европейского региона России характерен наибольший размах варьирования величины NEP (527 гС/(м2 год)), т.к. в этом регионе присутствуют как экосистемы являющиеся стоком атмосферного углерода большой мощности (более 250 гС/(м2 год)), так и экосистемы, функционирующие как источник атмосферного углерода средней мощности (125–134 гС/(м2 год)). Полученная оценка С–СО2 баланса в естественных степных экосистемах превышает в ~3.5 раза среднее значение стока диоксида углерода на лесных территориях России, оцениваемое в 66 ± 15 гС/(м2 год) [59], в 5 раз сток углерода в бореальных лесах Северного полушария, который составляет 46 гС/(м2 год) [80], в 2.9–5.6 раз значения стока атмосферного углерода в экосистемы субарктической тундры северо-востока Европейской части России, который оценивается от 41 гС/(м2 год) до 79 гС/(м2 год) [81].
Выполненные расчеты показали, что естественные степные экосистемы России ежегодно поглощают 111 ± 97 МтС. Полученное значение стока углерода атмосферы в естественные степные экосистемы России согласуется с диапазоном изменения этой величины от 92 до 121 МтС/год, опубликованным в [66]. Сток атмосферного углерода в природные и вторичные степные экосистемы Европейского, Сибирского и Центральноазиатского регионов России составляет, соответственно, 20, 37 и 43% годового депонирования углерода этими экосистемами и 18, 57 и 60% годовой NPP рассматриваемых экосистем.
Результаты исследования показали, что естественные экосистемы степной зоны на территории России демонстрируют высокий стоковый потенциал атмосферного углерода, в этих экосистемах наблюдается нарушение баланса углеродного цикла. Одна из причин углеродного дисбаланса в рассматриваемых экосистемах состоит в том, что около 58% территории естественных степей занимают залежные земли и бывшие пастбища (рис. 3б). На этой территории в настоящее время продолжаются процессы восстановления видовой структуры растительности и почвенного покрова, которые свойственны природным степным экосистемам. На всех стадиях восстановительного процесса во вторичных степных экосистемах существенно возрастает продуктивность растительных сообществ и содержание почвенного органического вещества [37, 52, 82, 83]. При восстановлении степных экосистем наблюдается увеличение интенсивности почвенного гетеротрофного дыхания, однако существенный рост величины RH в этих экосистемах сдерживается аридностью климата и увеличением термоизоляции верхних горизонтов почвы в процессе накопления надпочвенного слоя детрита и пополнения запасов органического вещества в почвах степей [84].
ЗАКЛЮЧЕНИЕ
Зона степей занимает 10% территории России. Степные экосистемы России подразделяются на луговые, настоящие, сухие и опустыненные, на долю которых приходится, соответственно, 41, 27, 19 и 13% рассматриваемой степной территории. Природные и вторичные степные экосистемы на территории России занимают 0.5 млн. км2, что составляет 28% общей площади степной зоны на территории РФ и 3% площади всех наземных экосистем страны. Рассматриваемые экосистемы расположены отдельными степными массивами в южных регионах России.
Согласно данным инвентаризации степных территорий, в Европейском регионе России находится 65% сохранившихся степных экосистем страны, в Сибирском и Центральноазиатском регионах – 9 и 26%, соответственно. Анализ данных о состоянии и использовании земель РФ показал, что современные площади, занимаемые вторичными степными экосистемами, в 1.4 раза превосходят площади природных степных экосистем, которые были выявлены в ходе инвентаризации степных территорий. В Европейском регионе России площадь вторичных степных экосистем в 1.1 раза меньше площади занятой природными степями. В Сибирском и Центральноазиатском регионах площади вторичных степных экосистем превышают площади природных степей в 4.5 и 1.4 раза, соответственно.
Проведенные расчеты показали, что естественные степные экосистемы России в настоящее время являются существенным стоком атмосферного СО2. Ежегодно эти экосистемы поглощают 111 ± 97 МтС (407 ± 355 МтСО2). Сток углерода в естественные степные экосистемы составляет 40% от продукционной составляющей углеродного цикла этих экосистем. Следует отметить, что углерод, депонируемый степными экосистемами, в основном аккумулируется в подземной части растительного сообщества, в фитодетрите (главным образом подземном) и гумусовых веществах почвы.
Количество атмосферного углерода, ежегодно депонируемое рассматриваемыми степными экосистемами, сравнимо со стоком углерода в управляемые леса РФ (96 МтС/год [85]) и составляет ~20% современного стока углерода в лесные экосистемы России, который оценивается в 530–595 МтС/год [86].
Наземные экосистемы России ежегодно депонируют 0.6–1.3 Гт атмосферного углерода [14, 87–89]. Согласно расчетам, выполненным в рамках настоящего исследования, природные и вторичные степные экосистемы РФ обеспечивают от 8% до 19% стока атмосферного углерода в наземные экосистемы России, что составляет существенную часть годового бюджета углерода на территории страны.
Проведенное исследование показало, что степные регионы обладают высоким углерододепонирующим потенциалом. Полученные оценки указывают, что естественные степные экосистемы способны оказывать значительное влияние на смягчение последствий климатических изменений и выступать в качестве существенных климатостабилизирующих систем. Антропогенная нагрузка значительно изменяет интенсивность продукционных и деструкционных потоков углерода в степных экосистемах и тем самым существенно влияет на углеродный газообмен между степными территориями и атмосферой.
Работа выполнена в рамках Государственного задания ИФА им. А.М. Обухова РАН (рег. № 1021032424681-6), Государственного задания ФИЦ ПНЦБИ РАН (рег. № 122040500037-6), при финансовой поддержке проекта “Развитие научно-образовательного Центра мониторинга климатически-активных веществ (Углерод в экосистемах: мониторинг)” в рамках ФНТП в области экологического развития Российской Федерации и климатических изменений на 2021–2030 годы.
Список литературы
Anav A., Friedlingstein P., Beer C. et al. Spatiotemporal patterns of terrestrial gross primary production: a review // Reviews of Geophysics. 2015. V. 53. P. 785–818.
Joiner J., Yoshida Y., Zhang Y. et al. Estimation of terrestrial global gross primary production (GPP) with satellite data-driven models and eddy covariance flux data // Remote Sensing. 2018. V. 10. 1346.
Badgley G., Anderegg L.D.L., Berry J.A., Field C.B. Terrestrial gross primary production: using NIRV to scale from site to globe // Global Change Biology. 2019. V. 25. P. 3731–3740.
Ciais, P., Sabine C., Bala G. et al. Carbon and Other Biogeochemical Cycles // Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge: Cambridge University Press., 2013. P. 465–570.
Randerson J.T., Chapin III F.S., Harden J.W. et al. Net ecosystem production: a comprehensive measure of net carbon accumulation by ecosystems // Ecological Applications. 2002. V. 12(4). P. 937–947.
Голубятников Л.Л., Денисенко Е.А. Модельные оценки влияния изменений климата на ареалы зональной растительности равнинных территорий России // Изв. РАН. Сер. биологическая. 2007. № 2. С. 212–228.
Friedlingstein P., Jones M.W., O’Sullivan M. et al. Global carbon budget 2021 // Earth Syst. Sci. Data. 2022. V. 14. P. 1917–2005.
Harris N.L., Gibbs D.A., Baccini A. et al. Global maps of twenty-first century forest carbon fluxes // Nature Climate Change. 2021. V. 11. P. 234–240.
Кобак К.И. Биотические компоненты углеродного цикла. Л.: Гидрометеоиздат, 1988. 248 с.
Почвенные процессы и пространственно-временная организация почв / Под ред. Кудеярова В.Н. М.: Наука, 2006. 568 с.
Raich J.W., Schlesinger W.H. The global carbon dioxide flux in soil respiration and its relationship to vegetation and climate // Tellus. 1992. V. B44(2). P. 81–99.
Rustad L.E., Huntington T.G., Boone R.D. Controls on soil respiration: implications for climate change // Biogeochemistry. 2000. V. 48. P. 1–6.
Xu M., Shang H. Contribution of soil respiration to the global carbon equation // J. Plant Physiology. 2016. V. 203. P. 16–28.
Кудеяров В.Н. Дыхание почв и биогенный сток углекислого газа на территории России (аналитический обзор) // Почвоведение. 2018. № 6. С. 643–658.
Bond-Lamberty B. New techniques and data for understanding the global soil respiration flux // Earth’s Future. 2018. V. 6. P. 1176–1180.
Кудеяров В.Н., Заварзин Г.А., Благодатский С.А. и др. Пулы и потоки углерода в наземных экосистемах России. М.: Наука, 2007. 315 с.
Reichstein M., Beer C. Soil respiration across scales: the importance of a model-data integration framework for data interpretation // J. Plant Nutrition and Soil Science. 2008. V. 171(3). P. 344–354.
Bond-Lamberty B., Thomson A.M. A global database of soil respiration data // Biogeosciences. 2010. V. 7(6). P. 1915–1926.
Заварзин Г.А. Лекции по природоведческой микробиологии. М.: Наука, 2004. 348 с.
Verburg P.S.J., Arnone III J.A., Obrist D. et al. Net ecosystem carbon exchange in two experimental grassland ecosystems // Global Change Biology. 2004. V. 10. P. 498–508.
Kim D., Lee M.-I., Seo E. Improvement of soil respiration parameterization in a dynamic global vegetation model and its impact on the simulation of terrestrial carbon fluxes // J. Climate. 2019. V. 32. P. 127–143.
Орлов Д.С., Бирюкова О.Н., Суханова Н.И. Органическое вещество почв Российской Федерации. М.: Наука, 1996. 256 с.
Щепащенко Д.Г., Мухортова Л.В., Швиденко А.З., Ведрова Э.Ф. Запасы органического углерода в почвах России // Почвоведение. 2013. № 2. С. 123–132.
Mikhailova E.A., Post C.J. Organic carbon stocks in the Russian Chernozem // European Journal of Soil Science. 2006. V. 57. P. 330–336.
Kucharik C.J., Fayram N.J., Cahill K.N. A paired study of prairie carbon stocks, fluxes, and phenology: comparing the world’s oldest prairie restoration with an adjacent remnant // Global Change Biology. 2006. V. 12. P. 122–139.
Berhongaray G., Alvarez R., De Paepe J. et al. Land use effects on soil carbon in the Argentine Pampas // Geoderma. 2013. V. 192. P. 97–110.
Мордкович В.Г. Степные экосистемы. Новосибирск: Академическое изд-во Гео, 2014. 170 с.
Базилевич Н.И., Титлянова А.А. Биотический круговорот на пяти континентах: азот и зольные элементы в природных наземных экосистемах. Новосибирск: Изд-во СО РАН, 2008. 381 с.
Чибилёв А.А. Степная Евразия: региональный обзор природного разнообразия. М., Оренбург: Институт степи РАН/РГО, 2016. 324 с.
Кёппен В. Основы климатологии. Климаты земного шара. М.: Учпедгиз наркомпроса РСФСР, 1938. 375 с.
Титлянова А.А., Базилевич Н.И., Шмакова Е.И. и др. Биологическая продуктивность травяных экосистем. Географические закономерности и экологические особенности. Новосибирск: ИПА СО РАН, 2018. 110 с.
Базилевич Н.И. Биологическая продуктивность экосистем северной Евразии. М.: Наука, 1993. 294 с.
Чибилёв А.А. Степи Северной Евразии. Екатеринбург: Уральское отд. РАН Институт степи, 1998. 102 с.
Инвентаризация сохранившихся степных экосистем (степных массивов) России. 2016. http://savesteppe.org.
Смелянский И. Сколько в степном регионе России залежей? // Степной бюллетень. 2012. № 36. С. 4–7.
Титлянова А.А., Самбуу А.Д. Сукцессии в травяных экосистемах. Новосибирск: Изд-во СО РАН, 2016. 191 с.
Люри Д.И., Горячкин С.В., Караваева Н.А. и др. Динамика сельскохозяйственных земель России в ХХ веке и постагрогенное восстановление растительности и почв. М.: ГЕОС, 2010. 416 с.
Kurganova I., Lopes de Gerenyu V., Kuzyakov Y. Large-scale carbon sequestration in post-agrogenic ecosystems in Russia and Kazakhstan // Catena. 2015. V. 133. P. 461–466.
Караваева Н.А., Денисенко Е.А. Постагрогенные миграционно-мицеллярные черноземы разновозрастных залежей южной лесостепи ЕТР // Почвоведение. 2009. № 10. С. 1165–1176.
Русанов А.М., Тесля А.В. Изменение основных свойств степных черноземов как результат их постагрогенной трансформации // Вестник Оренбургского ГУ. 2012. № 6(142). С. 98–102.
Титлянова А.А., Тесаржова М. Режимы биологического круговорота. Новосибирск: Наука, 1991. 150 с.
Сарула, Чэнь Х., Хоу С. и др. Запасы углерода в типичной степи при различном управлении выпасом // Почвоведение. 2014. № 11. С. 1365–1374.
Титлянова А.А., Косых Н.П., Кыргыс Ч.С. и др. Продуктивность травяных экосистем Тувы // Почвы и окружающая среда. 2020. Т. 3. № 2. e110.
Государственный (национальный) доклад о состоянии и использовании земель в Российской Федерации в 2019 году. М.: Федеральная служба государственной регистрации, кадастра и картографии Росреестр, 2020. 206 с.
Посевные площади Российской Федерации в 2020 году. Информационно-аналитические материалы Федеральной службы государственной статистики. http://rosstat.gov.ru/.
Титлянова А.А. Биологический круговорот углерода в травяных биогеоценозах. Новосибирск: Наука, 1977. 219 с.
Гаджиев И.М., Королюк А.Ю., Титлянова А.А. и др. Степи Центральной Азии. Новосибирск: Изд-во СО РАН, 2002. 299 с.
Титлянова А.А., Косых Н.П., Миронычева-Токарева Н.П., Романова И.П. Подземные органы растений в травяных экосистемах. Новосибирск: Наука, 1996. 128 с.
Титлянова А.А. Чистая первичная продукция травяных и болотных экосистем // Сибирский экологический журнал. 2007. № 5. С. 763–770.
Титлянова А.А., Шибарева С.В. Новые оценки запасов фитомассы и чистая первичная продукция степных экосистем Сибири и Казахстана // Изв. РАН. Сер. географическая. 2017. № 4. С. 43–55.
Чимитдоржиева Г.Д., Егорова Р.А., Мильхеев Е.Ю., Цыбенов Ю.Б. Потоки углерода в степных экосистемах (на примере Южного Забайкалья) // Растительный мир Азиатской России. 2010. № 2(6). С. 33–39.
Самбуу А.Д. Изменение чистой первичной продукции экосистем Тувы в связи с различным использованием земель // Вестник ТГУ. 2010. № 339. С. 172–177.
Golubyatnikov L.L., Denisenko E.A., Svirezhev Yu.M. Model of the total exchange carbon flux for terrestrial ecosystems // Ecological Modelling. 1998. V. 108. P. 265–276.
Raich J.W., Potter C.S., Bhagawatti D. Interannual variability in global soil respiration, 1980-94 // Global Change Biology. 2002. V. 8. P. 800–812.
Team, R Core. R: A language and environment for statistical computing. Vienna, Austria: R Foundation for Statistical Computing, 2020. https://www.R-project.org/.
Kurganova I.N. Carbon dioxide emission from soils of Russian terrestrial ecosystems. Interim Report, IR-02-070. Laxenburg, Austria: IIASA, 2003. 64 p.
Murray-Smith D.J. Methods for the external validation of continuous system simulation models: a review // Mathematical and Computer Modelling of Dynamical Systems. 1998. V. 4. P. 5–31.
Decostere B., De Craene J., Van Hoey S. et al. Validation of a microalgal growth model accounting with inorganic carbon and nutrient kinetics for wastewater treatment // Chemical Engineering Journal. 2016. V. 285. P. 189–197.
Швиденко А.З., Щепащенко Д.Г. Углеродный бюджет лесов России // Сибирский лесной журнал. 2014. № 1. С. 69–92.
Ведрова Э.Ф. Биогенные потоки углерода в бореальных лесах Центральной Сибири // Изв. РАН. Сер. биологическая. 2011. № 1. С. 77–89.
Осипов А.Ф. Влияние межгодовых различий метеорологических характеристик вегетационного периода на эмиссию СО2 с поверхности почвы среднетаежного сосняка бруснично-лишайникового (Республика Коми) // Почвоведение. 2018. № 12. С. 1455–1463.
Махныкина А.В., Прокушкин А.С., Меняйло О.В. и др. Влияние климатических факторов на эмиссию СО2 из почв в среднетаежных лесах Центральной Сибири: эмиссия как функция температуры и влажности почвы // Экология. 2020. № 1. С. 51–61.
Алферов А.М., Блинов В.Г., Гитарский М.Л. и др. Мониторинг потоков парниковых газов в природных экосистемах. Саратов: Амирит, 2017. 279 с.
Егоров В.П., Кривонос Л.A. Биологическая активность черноземов в агроценозах Курганской области // Почвы Западной Сибири и повышение их биологической активности. Омск: Изд-во ОмСХИ, 1983. С. 8–14.
Sarzhanov D.A., Vasenev V.I., Vasenev I.I. et al. Carbon stocks and CO2 emissions of urban and natural soils in Central Chernozemic region of Russia // Catena. 2017. V. 158. P. 131–141.
Курганова И.Н., Лопес де Гереню В.О., Жиенгалиев А.Т., Кудеяров В.Н. Углеродный бюджет степных экосистем России // Доклады АН. 2019. Т. 485. № 6. С. 732–735.
Mukhortova L., Schepaschenko D., Moltchanova E. et al. Respiration of Russian soils: climatic drivers and response to climate change // Science of the Total Environment. 2021. V. 785. 147314.
Craine F.M., Wedin D.A., Chapin III F.S. Predominance of ecophysiological controls on soil CO2 flux in a Minnesota grassland // Plant Soil. 1999. V. 207. P. 77–86.
Buyanovsky G.A., Kucera C.L., Wagner G.H. Comparative analyses of carbon dynamics in native and cultivated ecosystems // Ecology. 1987. V. 68. P. 2023–2031.
Gavrichkova O. Drivers of soil respiration of root and microbial origin in grasslands. Ph.D. thesis. Viterbo, Italy: University of Tuscia, 2009. 166 p.
Subke J.-A., Inglima I., Cotrufo M.F. Trends and methodological impacts in soil CO2 efflux partitioning: a metaanalytical review // Global Change Biology. 2006. V. 12. P. 921–943.
Ананьева Н.Д., Сушко С.В., Иващенко К.В., Васенев В.И. Микробное дыхание почв подтайги и лесостепи европейской части России: полевой и лабораторный подходы // Почвоведение. 2020. № 10. С. 1276–1286.
Stolbovoi V. Soil respiration and its role in Russia’s terrestrial C flux balance for the Kyoto baseline year // Tellus. 2003. V. B55. P. 258–269.
Perez-Quezada J.F., Saliendra N.Z., Akshalov K. et al. Land use influences carbon fluxes in northern Kazakhstan // Rangeland Ecology and Management. 2010. V. 63. P. 82–93.
Frank A.B., Dugas W.A. Carbon dioxide fluxes over a northern semiarid, mixed-grass prairie // Agriculture Forestry Meteorology. 2001. V. 108. P. 317–326.
Gilmanov T.G., Morgan J.F., Hanan N.P. et al. Productivity and CO2 exchange of Great Plains Ecoregions. I. Shortgrass steppe: flux tower estimates // Rangeland Ecology and Management. 2017. V. 70. P. 700–717.
Gilmanov T.G., Svejcar T.J., Johnson D.A. et al. Long-term dynamics of production, respiration, and net CO2 exchange in two sagebrush-steppe ecosystems // Rangeland Ecology and Management. 2006. V. 59. P. 585–599.
Belelli-Marchesini L. Analysis of the carbon cycle of steppe and old field ecosystems of Central Asia. Ph.D. thesis. Viterbo, Italy: University of Tuscia, 2007. 212 p.
Frank A.B. Carbon dioxide fluxes over a grazed prairie and seeded pasture in the northern Great Plains // Environmental Pollution. 2002. V. 116. P. 397–403.
Virkkala A.-M., Aalto J., Rogers B.M. et al. Statistical upscaling of ecosystem CO2 fluxes across the terrestrial tundra and boreal domain: regional patterns and uncertainties // Global Change Biology. 2021. V. 27. P. 4040–4059.
Marushchak M.E., Kiepe I., Biasi C. et al. Carbon dioxide balance of subarctic tundra from plot to regional scales // Biogeosciences. 2013. V. 10(1). P. 437–452.
Русанов А.М. Естественное восстановление агроландшафтов степной и лесостепной зон Оренбургской области // Степной бюллетень. 2012. № 36. С. 8–12.
Титлянова А.А., Шибарева С.В. Изменение чистой первичной продукции и восстановление запасов углерода в почвах залежей // Почвоведение. 2022. № 4. С. 500–510.
Карелин Д.В., Люри Д.И., Горячкин С.В. и др. Изменение почвенной эмиссии диоксида углерода в ходе постагрогенной сукцессии в черноземной лесостепи // Почвоведение. 2015. № 11. С. 1354–1366.
Замолодчиков Д.Г., Грабовский В.И., Коровин Г.Н. и др. Бюджет углерода управляемых лесов Российской Федерации в 1990–2050 гг.: ретроспективная оценка и прогноз // Метеорология и гидрология. 2013. № 10. С. 73–94.
Филипчук А.Н., Малышева Н.В., Золина Т.А., Югов А.Н. Бореальные леса России: возможности для смягчения изменения климата // Лесохозяйственная информация: электронный сетевой журнал. 2020. № 1. С. 92–114. http://lhi.vniilm.ru.
Ciais P., Canadell J.G., Luyssaert S. et al. Can we reconcile atmospheric estimates of Northern terrestrial carbon sink with land-based accounting? // Curr. Opin. Environ. Sustain. 2010. V. 2. P. 225–230.
Kurganova I.N., Kudeayrov V.N., Lopes de Gerenyu V.O. Updated estimate of carbon balance on Russian territory // Tellus. 2010. V. B62. P. 497–505.
Dolman H., Shvidenko A.Z., Schepaschenko D.G. et al. An estimate of the terrestrial carbon budget of Russia using inventory based, eddy covariance and inversion methods // Biogeosciences. 2012. V. 9. P. 5323–5340.
Li S.G., Asanuma J., Eugster W. et al. Net ecosystem carbon dioxide exchange over grazed steppe in central Mongolia // Global Change Biology. 2005. V. 11. P. 1941–1955.
Nagy Z., Pinter K., Czobel Sz. et al. The carbon budget of semi-arid grassland in a wet and a dry year in Hungary // Agriculture, Ecosystems and Environment. 2007. V. 121. P. 21–29.
Gilmanov T.G., Wylie B.K., Tieszen L.L. et al. CO2 uptake and ecophysiological parameters of the grain crops of midcontinent North America: estimates from flux tower measurements // Agriculture, Ecosystems and Environment. 2013. V. 164. P. 162–175.
Gilmanov T.G., Verma S.B., Sims P.L. et al. Gross primary production and light response parameters of four Southern Plains ecosystems estimated using long-term CO2-flux tower measurements // Global Biogeochemical Cycles. 2003. V. 17(2). 1071.
Suyker A.E., Verma S.B. Year-round observations of the net ecosystem exchange of carbon dioxide in a native tallgrass prairie // Global Change Biology. 2001. V. 7. P. 279–289.
Дополнительные материалы отсутствуют.
Инструменты
Известия РАН. Физика атмосферы и океана