

Физиология растений, 2019, T. 66, № 3, стр. 189-197
Распространенность С4-растений семейства маревых по профилю засоления почвы пустыни Кызылкум
П. Ю. Воронин a, Е. В. Шуйская a, К. Н. Тодерич b, c, Т. Ф. Ражабов d, Д. А. Ронжина e, f, Л. А. Иванова e, f
a Федеральное государственное бюджетное учреждение науки Институт физиологии растений
им. К.А. Тимирязева Российской академии наук
Москва, Россия
b Международная платформа по исследованию засушливых земель и образованию
(International Platform for Dryland Research and Education), Тоттори университет
Тоттори, Япония
c Международный центр по биоземледелию на засоленных почвах для Центральной Азии
и Закавказья
Ташкент, Узбекистан
d Самаркандский государственный университет
Самарканд, Узбекистан
e Федеральное государственное бюджетное учреждение науки Ботанический сад Уральского отделения
Российской академии наук
Екатеринбург, Россия
f Федеральное государственное автономное образовательное учреждение высшего образования
Тюменский государственный университет
Тюмень, Россия
Поступила в редакцию 27.03.2018
После доработки 13.06.2018
Принята к публикации 16.07.2018
Аннотация
На основании ботанико-географических описаний выявлены особенности распространенности С4-видов по рельефу пустыни Кызылкум. В низинах (0–50 м над уровнем моря) обнаружено преобладание С3-галофитов, на средних возвышенностях (50–200 м над уровнем моря) начинают преобладать галофиты с С4-фотосинтезом. Выше 200 м над уровнем моря отмечено резкое снижение вклада галофильной флоры в биоразнообразие местной растительности. Таким образом, проникновение галофитов выше 200 м ограничено наличием у них С4-фотосинтеза как адаптивного признака приспособления к рассолению почвы. При снижении засоления почвы у эдификаторных многолетних растений пустыни Кызылкум (тамарикс, саксаул и верблюжья колючка) отмечено снижение проявления суккулентности морфологии листа. В летний засушливый период С4-фотосинтез дает конкурентное преимущество по сравнению с С3-галофитами по выживанию на пересыхающих в летний сезон рассоленных почвах возвышенных частей рельефа местности. Сделан вывод о том, что приобретение C4-фотосинтеза галофильными видами представляет собой адаптацию фотосинтеза галофильной флоры к сезонному рассолению и иссушению почвы возвышенных частей рельефа пустыни Кызылкум.
ВВЕДЕНИЕ
Для морфологического строения листа С4-растения в большинстве случаев характерны черты кранц-анатомии, которые вероятно послужили структурной основой для формирования кооперативного механизма С4-фотосинтеза [1]. Общепринято, что конкурентное преимущество С4-видов обусловлено большей продукционной эффективностью С4-фотосинтеза в условиях пониженной атмосферной концентрации СО2, высокой инсоляции и температуры за счет использования особого кооперативного механизма концентрирования СО2 вблизи центров карбоксилирования. Эта биохимическая гипотеза также хорошо объясняет закономерную современную широтную распространенность С4-однодольных видов растений [2, 3]. Однако некоторые данные указывают на ограниченный характер биохимической гипотезы. Так, с позиций биохимической гипотезы не удается объяснить видовую специфику корреляции климатических факторов с распространенностью C4- одно- и двудольных растений. Широтное распространение C4-однодольных коррелирует наиболее тесно с минимумом температуры в середине вегетационного сезона, тогда как распространение C4-двудольных наиболее тесно коррелирует с сезонным эвапо-транспирационным режимом [2–4]. Кроме того, представительство C4-видов в составе местной флоры сильнее коррелирует с эдафическими факторами (засолением почвы, нарушением почвы), чем с широкомасштабным широтным фактором [5–7]. Поэтому некоторые исследователи связывают возникновение C4-синдрома не столько с глобальным снижением концентрации СО2 в атмосфере, сколько с формированием открытых ландшафтов и становлением сезонного, устойчивого чередования влажных и засушливых периодов климата [8].
Неполное согласие накопленных данных с биохимической гипотезой свидетельствует в пользу множественности причин, приводящих к множественному и независимому становлению C4-синдрома в различных растительных таксонах [9–12]. В связи с этим, проблему возникновения и представительства C4-видов в специфических флористических сообществах следует рассматривать в контексте конкретных обстоятельств их эволюционной истории [13].
Повышенная засоленность почвы является одним из факторов роста и развития C4-растений [14, 15]. Таким образом, зависимость почвенного засоления от топографии местности может непосредственно влиять на распространенность видов растений с C4-фотосинтезом среди растений галофильной флоры пустыни Кызылкум. В этом случае особенности топографии местного ландшафта могли послужить ведущим фактором распространенности C4-синдрома у галофитов поскольку все остальные климатические показатели пустыни Кызылкум в целом однородны.
Цель данной работы состояла в том, чтобы на основе анализа анатомических изменений листьев растений, представляющих основные эдификаторные флористические группы пустыни Кызылкум, по профилю естественного засоления почвы и распространенности С4-видов семейства маревые (Chenopodiaceae) в биоразнообразии местных растительных сообществ оценить значение ландшафтно-топографического фактора на распространение С4-фотосинтеза у галофитов пустыни Кызылкум.
МАТЕРИАЛЫ И МЕТОДЫ
Описание местности проведения исследования. Местность исследования представляла собой пустынные: в подгорной части щебнистые и в равнинной части песчано-глинистые – засоленные территории (рис. 1). Кызылкум простирается между двумя крупнейшими реками Центральной Азии – Сырдарьей на востоке и Амударьей на юге и юго-западе. В целом рельеф местности понижается с подгорных равнин на юго-востоке к побережью Аральского моря на северо-западе. Для реконструкции распространенности видов маревых использовали ботанико-географическую карту пустыни Кызылкум [16], топографические карты местности, навигатор и альтиметр GPS (“Magellan”, США). Характеристика мест взятия проб почвы и растительного материала представлена в табл. 1.
Рис. 1.
Карта-схема географического расположения пустыни Кызылкум (a) и ботанико-географического распространения С4-видов растений сем. Chenopodiaceae по пустыне Кызылкум (б). Вклад С4-маревых в видовое разнообразие растительных сообществ на территории отмечен разным тоном: бесцветный тон соответствует 0%, светлый тон соответствует 30%, точечное тонирование соответствует 30–60%, темный тон соответствует (60%).

Таблица 1.
Характеристика мест взятия проб почвы и растительного материала вдоль высотного градиента топографии местности (подгорные равнины песчаной пустыни Кызылкум)
| № | Название, координаты и тип рельефа местности | Высота над уровнем моря, м |
|---|---|---|
| 1 | Нурата, 40°27′ с.ш., 66°09′ в.д. | |
| cклон | 900 | |
| 820 | ||
| равнина | 669 | |
| 668 | ||
| 2 | Айдаркуль, 40°58′ с.ш., 65°56′ в.д. | |
| склон | 290 | |
| 251 | ||
| равнина | 247 | |
| 3 | Зафарабад, 40°32′ с.ш., 65°01′ в.д. | |
| склон | 378 | |
| 348 | ||
| 4 | Кызылкесек, 41°02′ с.ш., 64°54′ в.д. | |
| склон | 132 | |
| 121 | ||
| 115 | ||
| равнина | 115 | |
| 108 | ||
| 105 |
Растительный материал. Изучали морфологическое строение листа и фотосинтезирующих побегов видов-представителей основных эдификаторных групп – галофильной и ксерофильной флоры Кызылкума. В этом ряду древовидный влаголюбивый галофит Tamarix hispida (Tamaricaceae) и многолетний травянистый ксерофит Alhagi pseudalhagi (Fabaceae) представляли эдификаторные С3-виды пустыни Кызылкум. С4-виды представлял эдификаторный древовидный суккулентный, соле- и засухоустойчивый кустарник Haloxylon aphyllum (Chenopodiaceae).
Содержание соли в почве и растительном материале. Ландшафтный профиль засоления характеризовали содержанием катионов Na+, Ca2+, K+ в пробах почвы. Почвенные пробы брали с глубины 40 см. Содержание ионов определяли в водных экстрактах из почвенных проб и растительного материала (100 мг материала на пробу) с помощью атомно-абсорбционного спектрофотометра (Hitachi 207, Япония). Пробы почвы высушивали до постоянно сухого веса при температуре 100°C для определения в них содержания воды.
Показатели морфологического строения листа определяли с помощью системы цифрового анализа изображений Simagis Mesoplant (ООО “СИАМС”, Россия). Для анатомических исследований собирали неповрежденные, полностью сформированные листья среднего яруса со здоровых, развитых растений (3 растения для каждого вида). Из средней части листьев делали высечки и фиксировали их в 3.5%-растворе глутарового альдегида в фосфатном буфере (рН = 7.4). Далее делали поперечные срезы толщиной 15 мкм на криомикротоме MicromHM 525 (“Сarl Zeiss”, Германия). Толщину тканей определяли на поперечных срезах в 10 аналитических повторностях, используя световой микроскоп ZeissAxiostar (“Сarl Zeiss”, Германия) при увеличении ×200 и систему цифрового анализа изображений Simagis Mesoplant (ООО “СИАМС”, Россия).
На графиках приведены средние значения показателей со стандартной ошибкой.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Предполагают, что усиление аридности и сезонности климата с начала миоцена послужило одной из главных экологических предпосылок широкого распространения видов с С4-фотосинтезом на открытых ландшафтах [8].
Сезонно-топографическая детерминанта режима рассоления и иссушения почвы пустыни Кызылкум
По своему характеру рельеф Кызылкум представляет собой слегка холмистую, местами сильно расчлененную равнину. Для Кызылкума характерен резко континентальный климат. Само возникновение открытого ландшафта пустыни Кызылкум в ряду других Туранских пустынь связано с высыханием в эоцене древнего эпиконтинентального моря. Об этом свидетельствует близость пропорции основных катионов (Na : Ca : K) в морской воде к их соотношению в почве солончаков пустыни Кызылкум (табл. 2). Отмечено усиление засоленности (рис. 2) и увлажнения (рис. 3) почвы от возвышенной подгорной равнины на юго-востоке к болотистому побережью Аральского моря на северо-западе Кызылкума. Сезонность климата пустыни Кызылкум проявляется в устойчивом чередовании засухи в летние месяцы и выпадении дождей осенью и зимой. Дождевые осадки промывают верхний слой почвы местных возвышенностей и смывают соли в низинные части рельефа. В историческом масштабе времени сезонная цикличность температурно-влажностного режима формирует высотное снижение засоления и влажности почвы от понижений к возвышенным частям рельефа местности. Возвышенные части рельефа характеризуются минимальным засолением и влажностью почвы (рис. 2 и 3). При этом отмечена однозначная прямо пропорциональная зависимость влажности почвы от ее засоления (рис. 4). Уменьшение засоления почвы возвышенных частей ландшафта увеличивает водный потенциал почвы, а значит, способствует ее иссушению.
Таблица 2.
Содержание (мкмоль/г) наиболее распространенных катионов в морской воде и в мокрых солончаках пустыни Кызылкум
| Субстрат | Концентрация, мкмоль/г (соотношение, %) | Источник данных | ||
|---|---|---|---|---|
| Na+ | Ca2+ | K+ | ||
| Солончак | 650 (100) | 34 (5) | 4 (1) | Настоящая работа |
| Море | 469 (100) | 10 (2) | 10 (2) | [17] |
Рис. 2.
Зависимость засоления почвы от топографической высоты местности. Почвенное засоление определяли по содержанию в почве основных катионов Na+ (a, б), Ca2+ (в, г) и K+ (д, е) в почве равнинных ландшафтов (a, в, д) и склонов холмов (б, г, е). Цифрами на графиках отмечены места взятия проб, описанные в таблице 1. Оси ординат представлены в логарифмическом масштабе. Точки на графиках представляют средние значения ± стандартные отклонения (n = 9).
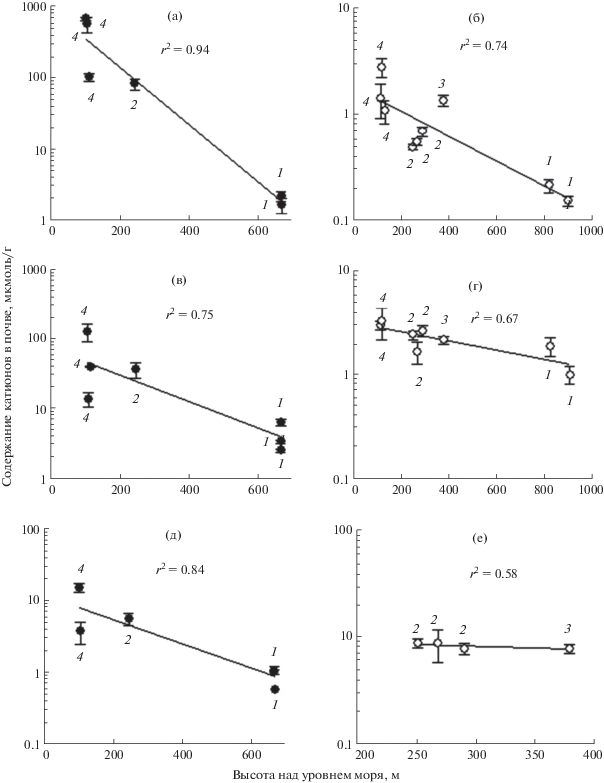
Рис. 3.
Зависимость относительной влажности почвы от топографической высоты равнинных ландшафтов местности (а) и склонов холмов (б) пустыни Кызылкум. Цифрами на графиках отмечены места взятия проб, описанные в таблице 1. Точки на графиках представляют средние значения ± стандартные отклонения от среднего (n = 9).


Распространенность С4-галофитов по ландшафту пустыни Кызылкум
Низинные территории пустынного ландшафта являются местообитанием галофильной флоры. Сохранение влаги в засоленных почвах в низинных частях рельефа местности обеспечивает выживание многолетних галофильных растений во время засушливого летнего сезона. В Кызылкуме растительные сообщества, в составе которых присутствуют С4-виды, наиболее часто встречаются на высотах от 50 до 200 м над уровнем моря. Так растительные сообщества с заметным присутствием С4-видов (>30%) так же, как и с их доминированием (>60%) обнаруживали на высоте 50–100 м над уровнем моря. Это означает, что наибольшее число С4-видов встречается локально по рельефу с высотой около 100 м над уровнем моря (табл. 3), соответствующей умеренному засолению и слабому увлажнению почвы (рис. 2 и 3).
Таблица 3.
Распространенность C4-видов (Chenopodiaceae) среди растительных сообществ пустыни Кызылкум по рельефу местности (n = 192)
| Высота над уровнем моря, м | Классовые интервалы вклада C4-маревых в биоразнообразие растительных сообществ, % | Растительные сообщества с С4-маревыми в своем составе, % | ||
|---|---|---|---|---|
| 0–30 | 30–60 | 60–100 | ||
| 0–50 | 2.6 | 1.0 | 0.0 | 3.6 |
| 50–100 | 7.8 | 15.6 | 11.5 | 34.9 |
| 100–200 | 20.3 | 17.2 | 2.1 | 39.6 |
| 200–300 | 15.1 | 5.2 | 1.6 | 21.9 |
| Всего, % | 45.8 | 39.0 | 15.2 | 100 |
Поскольку растения из семейства маревых доминируют в галофильной флоре Центральной Азии [18–20], их представительство в растительном биоразнообразии является показателем представительства С4-синдрома у местной галофильной флоры. Увеличение доли С4-галофитов из семейства маревых в местном биоразнообразии по сравнению с С3-видами вплоть до определенного предела следовало за общим рассолением почвы (рис. 5). При этом в возвышенных сезонно-пересыхающих местообитаниях с относительно рассоленными (умеренно засоленными) почвами преобладала С3-ксерофильная флора, а представительство С4-растений снижалось (табл. 3). Согласно нашим данным, засоленность почвы на склонах холмов пустыни Кызылкум на несколько порядков меньше, чем в ее низинах (рис. 2). Эта особенность является причиной исчезновения большинства С4-маревых из местной галофильной флоры на высотах больше 200 м над уровнем моря. Таким образом, в результате сезонного выпадения дождей, возникает устойчивая тенденция к рассолению почвы локально возвышенных частей рельефа, и присущие С4-маревым галофильные свойства определяют предел их местного высотного распространения. В результате, распространение галофильной флоры пустыни Кызылкум ограничено высотой рельефа около 100 м над уровнем моря, на которой в местных растительных сообществах доля галофитов с С4-фотосинтезом максимальна (табл. 3).
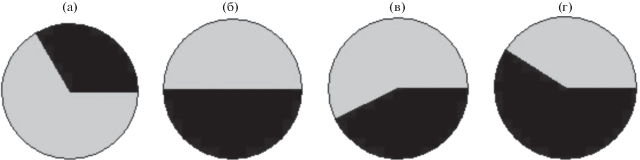
Рис. 5.
Соотношение между C3-видами (серый цвет) и C4-видами сем. Chenopodiaceae (черный цвет) в Кызылкесеке (см. табл. 1) по градиенту концентрации Na+ в почве: а – 0.3 мкмоль/г, б – 20 мкмоль/г, в – 60 мкмоль/г, г – 80 мкмоль/г.
Снижение суккулентности листа сопровождает продвижение галофильной флоры от низин к возвышенностям местного рельефа пустыни Кызылкум
По сравнению с рассоленными почвами возвышенных ландшафтов в местах наибольшего засоления у трех эдификаторных видов пустыни Кызылкум проявлялась тенденция к укрупнению размеров клеток покровных тканей листа (табл. 4). Известно, что суккулентность листа является чертой галофильности С3- и С4-растений [21, 22]. Соленакопление в листьях обеспечивает привлечение воды в листья за счет снижения в них водного потенциала. Усиление оводненности тканей листа в свою очередь приводит к укрупнению клеток покровных тканей, являющихся конечными пунктами на пути движения воды по растению из почвы в атмосферу. Укрупнение клеток покровных тканей листа отчетливо наблюдали у галофильных видов: тамарикса (С3-растение) и саксаула (С4-растение) (табл. 4). Так в Кызылкуме, по сравнению с С3-ксерофитом A. pseudalhagi (0.05–0.50 ммоль Na+/г), суккулентные галофильные виды H. aphyllum (3–5 ммоль Na+/г), T. hispida (1–4 ммоль Na+/г) в разы больше накапливали соль в листьях. Поэтому единообразное проявление С4-видами сем. Chenopodiaceae галофильности отражено в морфологии их листьев наличием черт суккулентности и кранц-анатомии [23, 24]. Большую долю (50–80%) видов с С4-фотосинтезом в Центральной Азии составляют виды семейства маревых [25]. Большинство С3- и С4-видов коренной флоры Кызылкума являются галофитами. Поэтому закономерное рассоление и иссушение почвы от низин к возвышенным частям рельефа, а также снижение суккулентности листьев галофильных С3- и С4-растений (табл. 4) и снижение вклада С3-галофильных видов в растительное биоразнообразие, позволяют предполагать, что эволюционным ответом галофильной флоры на рассоление и иссушение почвы в возвышенностях рельефа пустыни Кызылкум был переход маревых С3-галофитов на С4-тип фотосинтеза (рис. 5).
Таблица 4.
Количественная характеристика тканей листа эдификаторных многолетних растений пустыни Кызылкум (n = 30)
| Вид растения | Ткань | Места сбора образцов* | |||
|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | ||
| Haloxylon aphyllum | Эпидерма | 19.2 ± 1.7a | 18.8 ± 1.3a | 23.1 ± 2.1b | 26.5 ± 3.6b |
| – кутикула | 2.0 ± 0.2a | 2.2 ± 0.2a | 2.1 ± 0.2a | 2.3 ± 0.2a | |
| Гиподерма | 15.0 ± 1.5a | 18.6 ± 1.0b | 21.0 ± 0.6c | 19.6 ± 0.7bc | |
| Мезофилл | 27.7 ± 3.7a | 35.9 ± 3.5b | 39.3 ± 5.9b | 39.1 ± 2.1b | |
| Обкладка сосудистых пучков | 15.1 ± 1.2a | 22.5 ± 1.7c | 19.2 ± 1.3b | 19.1 ± 0.7b | |
| Tamarix hispida | Эпидерма | – | 20.0 ± 1.1a | 22.5 ± 1.9a | 17.4 ± 0.6b |
| – кутикула | – | 2.4 ± 0.2a | 4.3 ± 0.5b | 4.3 ± 0.3b | |
| Палисадный мезофилл | – | 55.7 ± 1.6a | 58.4 ± 1.6a | 60.6 ± 3.7a | |
| Alhagi pseudalhagi | Эпидерма | 21.7 ± 0.5a | 22.8 ± 0.5b | 25.1 ± 0.6c | 26.7 ± 0.6d |
| – кутикула | 3.1 ± 0.1a | 2.7 ± 0.1b | 2.3 ± 0.1c | 2.7 ± 0.1b | |
| Гиподерма | 43.5 ± 0.9a | 44.9 ± 0.9ab | 46.6 ± 1.0b | 45.6 ± 0.9b | |
| Мезофилл | 37.5 ± 0.4a | 43.2 ± 0.4d | 38.5 ± 0.4b | 41.7 ± 0.4c | |
Примечание. *В местностях 1, 2 и 4 (см. табл. 1) образцы были собраны на равнинных участках. Разными латинскими буквами отмечены достоверные различия на уровне p < 0.05. Толщина тканей указана в мкм.
Реконструкция становления С4-фотосинтеза среди галофитов семейства маревых в пустыне Кызылкум
Главным климатическим трендом в пустыне Кызылкум является усиление аридности климата с сопутствующим понижением грунтовых вод. Сезонное пересыхание солончаков и конечное уменьшение их площади являются в исторической перспективе существенным фактором эволюционного отбора галофильной флоры на устойчивость к засухе. Локальное разнообразие видового состава растений с С3- и С4-типами фотосинтеза и различной степенью выраженности ксероморфизма и суккулентности листа обусловлено в основном мозаичным характером сложения почв с разными физико-химическими свойствами (загипсованность, лессы, глинистость и щебнистость), их минеральным составом, увлажнением, а также особенностями микрорельефа. По нашему мнению, относительно недавнее освоение этой экологической ниши видами прибрежно-средиземноморской зоны, в которой маревые составляют традиционно высокую долю, обусловило быстрый процесс эволюции (5–6 млн лет) различных вариантов фотосинтеза (С3, С3–С4, С4), а также богатство морфотипов и жизненных форм.
Для галофитов сем. Chenopodiaceae способность переносить летнюю почвенную засуху в активном состоянии за счет С4-фотосинтеза несомненно представляет собой ценный адаптивный признак. В свою очередь сезонное рассоление и пересыхание почвы возвышенных частей микрорельефа местности и снижение суккулентных черт строения листа галофитов в этих местообитаниях в новом свете представляют известную проблему географической идентификации “анцестрального С3-центра маревых” и становления в этом семействе С4-фотосинтеза [25–27]. Можно предположить, что движение анцестральных С3-видов сем. Chenopodiaceae за отступающей кромкой древнего высыхающего эпиконтинентального моря завершилось их современной локализацией вблизи береговой линии Средиземного, Каспийского и Аральского морей. Виды сем. Chenopodiaceaе остаточных популяций на пути этого движения подвергались совместному сезонному воздействию пересыхания-рассоления почвы на возвышенных частях микрорельефа. В этих новых условиях они были ограничены в своих возможностях, как галофиты, привлекать воду в лист за счет соленакопления. Это отражалось в снижении суккулентности их листьев. Потребность в увеличении эффективности использования воды на базе предсуществующей кранц-анатомии листа вероятно привела к формированию у них С4-фотосинтеза.
Список литературы
Laetsch W.M. The C4-syndrome: a structural analysis // Ann. Rev. Plant Physiol. 1974. V. 25. P. 27–52.
Teeri J.A., Stowe L.G. Climatic patterns and the distribution of C4 grasses in North America // Oecologia. 1976. V. 23. P. 1–12.
Hattersley P.W. The distribution of C3 and C4 grasses in Australia in relation to climate // Oecologia. 1983. V. 57. P. 113–128.
Stowe L.G., Teeri J.A. The geographic distribution of C4 species of the Dicotyledoneae in relation to climate // Am. Naturalist. 1978. V. 112. P. 609–623.
Collins R.P., Jones M.B. The influence of climatic factors on the distribution of C4 species in Europe // Vegetatio. 1985. V. 64. P. 121–129.
Perelman S.B., Len R.J.C., Oesterheld M. Cross-scale vegetation patterns of Flooding Pampa grasslands // J. Ecol. 2001. V. 89. P. 562–577.
Feldman S.R., Bisaro V., Biani N.B., Prado D.E. Soil salinity determines the relative abundance of C3/C4 species in Argentinean grasslands // Glob. Ecol. Biogeogr. 2008. V. 17. P. 708–714.
Osborne C.P., Fleckleton R.P. Ecological selection pressures for C4 photosynthesis in the grasses // Proc. R. Soc. B. 2008. V. 276. P. 1753–1760.
Akhani H., Edwards G., Roalson E.H. Diversification of the old world Salsoleae s.l. (Chenopodiaceae): molecular phylogenetic analysis of nuclear and chloroplast data sets and a revised classification // Int. J. Plant. Sci. 2007. V. 168. P. 931–956.
Гамалей Ю.В., Шереметьев С.Н. С4-травы Старого и Нового света // Ботан. журн. 2009. Т. 94. С. 1753–1770.
Edwards E.J., Smith S.A. Phylogenetic analysis reveal the shady history of C4 grasses // Proc. Natl. Acad. Sci. 2010. V. 107. P. 2532–2537.
Edwards E.J., Osborne C.P., Stromberg C.A.E., Smith S.A., Bond W.J., Christin P.A., Cousins A.B., Duvall M.R., Fox D.L., Freckleton R.P., Ghannoum O., Hartwell J., Huang Y., Janis C.M., Keeley J.E., Kellogg E.A., Knapp A.K., Leakey A.D., Nelson D.M., Saarela J.M., Sage R.F., Sala O.E., Salamin N., Still C.J., Tipple B. The origins of the C4 grasslands: integrating evolutionary and ecosystem science // Science. 2010. V. 328. P. 587–591.
Taub D.R. Climate and the US distribution of C4 grass subfamilies and decarboxylation variants of C4 photosynthesis // Am. J. Botany. 2000. V. 87. P. 1211–1215.
Brownell P.F., Crossland C.J. The requirement for sodium as a micronutrient by species having the C4 dicarboxylic photosynthetic pathway // Plant Physiol. 1972. V. 49. P. 794–797.
Fisher D.D., Schenk H.J., Thorsch J.A., Ferren W.R. Leaf anatomy and subgeneric affiliation of C3 and C4 species of Suaeda (Chenopodiaceae) in North America // Am. J. Bot. 1997. V. 84. P. 1198-1210.
Карта растительности Казахстана и Средней Азии в пределах пустынной области (масштаб 1 : 2 000 500) / Ред. Е.И. Рачковская. М.: ТОО “Экор”, 1995.
Horn R.A. Marine chemistry. The structure of water and the chemistry of the hydrosphere. Wiley Interscience New York, London, Sydney, Toronto. 1969.
Пьянков В.И., Мокроносов А.Т. Основные тенденции изменения растительности Земли в связи с глобальным потеплением климата // Физиология растений. 1993. Т. 40. С. 515–530.
Akhani H., Trimborn P., Ziegler H. Photosynthetic pathways in Chenopodiaceae from Africa, Asia and Europe with their ecological, phytogeographical and taxonomical importance // Pl. Syst. Evol. 1997. V. 206. P. 187–221.
Pyankov V.I., Gunin P.D., Tsoog Sh., Black C.C. C4 plants in the vegetation of Mongolia: their natural occurance and geographical distribution in relation to climate // Oecologia. 2000. V. 123. P. 15–31.
Воронин П.Ю., Манжулин А.В., Мясоедов Н.А., Балнокин Ю.В., Терентьева Е.И. Морфологические типы и фотосинтез листьев С4-растений при долговременном засолении почвы // Физиология растений. 1995. Т. 42. С. 356–367.
Балнокин Ю.В., Мясоедов Н.А., Шамсутдинов З.Ш., Шамсутдинов Н.З. Роль Na+ и K+ в поддержании оводненности тканей органов у галофитов сем. Chenopodiaceae различных экологических групп // Физиология растений. 2005. Т. 52. С. 779–787.
von Willert D.J., Eller B.M., Werger M.J.A., Brinckmann E. Desert succulents and their life strategies // Vegetation. 1990. V. 90. P. 133–143.
Akhani H., Ghasemkhani M. Diversity of photosynthetic organs in Chenopodiaceae from Golestan National Park (NE Iran) based on carbon isotope composition and anatomy of leaves and cotyledons // Nova Hedwig Beih. 2007. V. 131. P. 265–277.
Pyankov V., Ziegler H., Kuz’min A., Edwards G. Origin and evolution of C4 photosyntesis in the tribe Salsoleae (Chenopodiaceae) based on anatomical and biochemical types in leaves and cotyledons // Plant System. and Evol. 2001. V. 230. P. 43–74.
Pyankov V., Black C., Stichler W., Ziegler H. Photosynthesis in Salsola species (Chenopodiaceae) from Southern Africa relative to their C4 syndrome origin and their African-Asian arid zone migration pathways // Plant Biol. 2002. V. 4. P. 62-69.
Камелин Р.В. Древнексерофильное семейство Chenopodiaceae во флоре Турана и Центральной Азии // Ботан. журн. 2011. Т. 96. С. 441–464.
Дополнительные материалы отсутствуют.
Инструменты
Физиология растений