

Физиология растений, 2022, T. 69, № 6, стр. 563-571
Дыхание растений: классические и современные представления
Т. К. Головко a, *, Е. В. Гармаш a, **
a Институт биологии Коми научного центра Уральского отделения Российской академии наук
Сыктывкар, Россия
* E-mail: golovko@ib.komisc.ru
** E-mail: garmash@ib.komisc.ru
Поступила в редакцию 07.04.2022
После доработки 14.04.2022
Принята к публикации 14.04.2022
- EDN: YVCSDD
- DOI: 10.31857/S0015330322060070
Аннотация
В кратком обзоре изложены современные представления о дыхании растений, его роли в жизнедеятельности, устойчивости и продуктивности. Приведены общие сведения об организации и регуляции дыхательного метаболизма, путях генерации и диссипации энергии, влиянии факторов среды на метаболизм и энергетику дыхания. Намечены направления дальнейших исследований.
ВВЕДЕНИЕ
В эволюционном ряду развития жизни на Земле дыхание занимает особое место как процесс, обеспечивающий функционирование организмов энергией и метаболитами. О его масштабах свидетельствует тот факт, что за сутки в процессе дыхания растения окисляют до 70% продуктов фотосинтеза [1, 2], а растительность надземных экосистем возвращает в атмосферу по меньшей мере половину всего ассимилированного за год СО2 [3].
Основу клеточного дыхания составляют гликолиз, цикл трикарбоновых кислот (ЦТК) и митохондриальная электрон-транспортная цепь (мЭТЦ), основная функция которой состоит в генерации АТФ (рис. 1). Митохондрии взаимодействуют с ядром и хлоропластами, участвуют в клеточном сигналинге и поддержании редокс-баланса [4]. Митохондриальная ретроградная регуляция играет важную роль в реакции растений на стресс, активирует целевые гены, сообщает об изменении энергетической эффективности и в тесной коммуникации с другими сигнальными путями клетки направляет метаболизм с роста на адаптацию к неблагоприятным воздействиям [5]. Наличие множества альтернативных дыхательных путей и ферментов придает растениям поразительную пластичность, способность программировать метаболизм и энергетику клеток в соответствии с этапом онтогенетического развития и изменениями условий среды. По мнению О.А. Семихатовой [6] эффективность обмена энергии при дыхании (ЭЭД) в норме и при стрессе является одним из кардинальных вопросов биологии.
Рис. 1.
Схема ключевых этапов дыхания и использования дыхательной энергии в растении. Вовлечение альтернативного и других нефосфорилирующих путей в ЭТЦ митохондрий неизбежно приводит к диссипации энергии, но необходимо для поддержания гомеостаза растительной клетки. Обсуждение в тексте. Сокращения: ОА – оксалоацетат; 2‑ОГ – 2-оксоглутарат; Гл-6-Ф – глюкозо-6-фосфат; Рибулозо-5-Ф – рибулозо-5-фосфат; ОПФП – окислительный пентозофосфатный путь; ПВК – пировиноградная кислота (пируват); ФД –фотодыхание; ФЕП – фосфоенолпируват; ЭЭД – энергетическая эффективность дыхания.
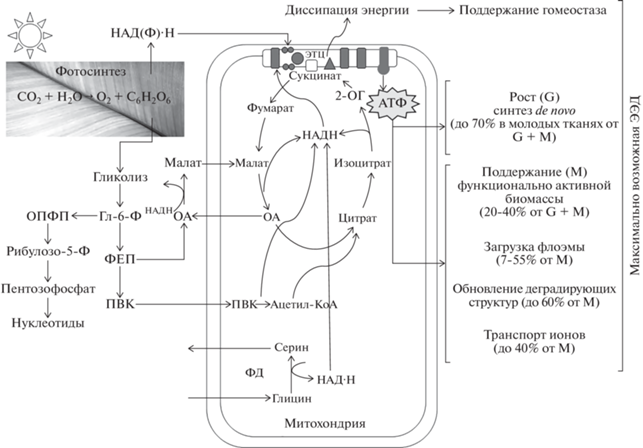
Этапы дыхания подробно описаны О.А. Семихатовой и Т.В. Чирковой [7]. Отметим ключевые моменты и остановимся на некоторых новых сведениях о химизме и физиологии дыхательного процесса. Начальный этап дыхания – гликолиз – процесс анаэробного распада гексозы до пировиноградной кислоты (ПВК) с образованием АТФ, НАД·Н и различных предшественников для анаболизма, локализован в цитозоле и пластидах. Пластидный и цитозольный гликолитические пути взаимодействуют через высокоселективные транспортеры, локализованные во внутренней мембране пластид. В цитозоле растительных клеток гликолизу подвергаются сахароза и глюкоза, а в пластидах гетеротрофных тканей и хлоропластах – преимущественно крахмал. Дисахарид сахароза предварительно гидролизуется, образуя мономеры глюкозу и фруктозу. Чтобы стать субстратом, доступным для дыхательных превращений, крахмал деградирует под действием ферментов. Образовавшиеся глюкозные мономеры выделяются из пластид в цитозоль, где и подвергаются анаэробному распаду. В цепи реакций гликолиза выделяют несколько ключевых этапов: фосфорилирование молекулы глюкозы (Глю) и ее расщепление на две фосфотриозы, окисление фосфоглицеринового альдегида до фосфоенолпирувата (ФЕП) с восстановлением НАД+ и превращение ФЕП в пировиноградную кислоту (ПВК). Под влиянием внешних факторов метаболизм сахарозы и ФЕП может осуществляться по альтернативным путям. Например, участие пирофосфата в фосфорилировании фруктозо-6-фосфата (Фру-6-Ф) или в активном транспорте Н+ в вакуоль из цитозоля при стрессе способствует сохранению пула АТФ [8].
В отличие от животных клеток, где весь ФЕП превращается в ПВК, в растительных клетках часть ФЕП преобразуется в малат [9]. При взаимодействии ФЕП с ФЕП-карбоксилазой и ${\text{HCO}}_{3}^{ - }$ образуется оксалоацетат (ОА), который превращается в малат при участии НАД(Ф)·Н-зависимой малатдегидрогеназы. В цитоплазме возможно также прямое превращение ПВК в малат (восстановительное карбоксилирование), но вклад этого процесса малосущественен [7]. Ключевые ферменты гликолиза контролируются аденилатами, его скорость возрастает при повышении потребности в энергии (АТФ). Однако наличие обходных путей снижает зависимость дыхания от аденилатов.
Наряду с гликолизом в клетках функционирует пентозофосфатный путь (ПФП) распада глюкозы, который включает окислительную и восстановительную ветви [10]. Окислительная ветвь превращает Глю-6-Ф в СО2 с образованием рибулозо-5-фосфата (Рибулозо-5-Ф) и НAД(Ф)·Н. Ток углеводов через окислительный ПФП усиливается при стрессе и нарушении митохондриальных функций, обеспечивая тем самым образование предшественников для синтеза нуклеотидов, сохранение метаболического и редокс-гомеостаза. При окислительном стрессе митохондрий окислительный ПФП и анаплеротические пути метаболизации Глю способствуют поддержанию пула НАД·Н [11]. Неокислительная ветвь ПФП метаболизирует гликолитические интермедиаты, снабжая метаболитами синтез нуклеиновых и аминокислот, что особенно важно для пролиферирующих клеток. Неокислительный ПФП может также поставлять в гликолиз промежуточные продукты метаболизации рибозо-5-фосфата.
Продукты гликолиза поступают из цитозоля в митохондрии с помощью специфических транслокаторов [8]. ПВК окисляется в ЦТК. Малат декарбоксилируется с участием НАД-зависимого малик-энзима до ПВК и/или окисляется малатдегидрогеназой с образованием ОА. Если бы ПВК была единственным субстратом, поставляемым в ЦТК, то экспорт метаболитов из цикла для различных биосинтезов (включая ассимиляцию азота) привел бы к его остановке. ЦТК обеспечивает клетки и ткани различными метаболитами, тесно связан с обменом углеводов, белков и липидов. Большинство ферментов ЦТК локализованы в матриксе митохондрий или образуют комплексы, ассоциированные с внутренней мембраной. Реакции дегидрирования приводят к образованию НАД·Н и ФАД·Н2, которые окисляются в мЭТЦ. Регуляция цикла осуществляется соотношением концентраций НАД+/НАД·Н и АДФ/АТФ.
В условиях стресса, помимо ПВК и малата, окислению в митохондриях могут подвергаться продукты катаболизма белков и липидов. Например, при углеводном голодании в качестве дыхательного субстрата используются аминокислоты [12]. При осмотическом и солевом стрессах накапливается значительное количество пролина, который затем может быть окислен в процессе дыхания [13]. У С3-растений в условиях, активирующих фотодыхание, в митохондриях окисляется НАД·Н, образованный при превращении двух молекул глицина в серин [14].
В глиоксисомах локализован глиоксилатный цикл, представляющий собой модификацию ЦТК [15]. Он участвует в утилизации запасных липидов, ß-окисление которых приводит к образованию ацетил-КоА. Сукцинат выходит из глиоксисом и может участвовать в ЦТК или глюконеогенезе. В реакции дегидрирования малата до ОА восстанавливается одна молекула НАД+. Таким способом из запасных веществ образуются необходимые для роста проростков метаболиты и НАД·Н.
Тонкий контроль метаболизма углеводов в дыхательных путях растительной клетки осуществляется, в первую очередь, ферментами, катализирующими реакции гликолитического превращения гексоз в ПВК или ОА, превращения ПВК в ацетил-КоА с последующей его метаболизацией в ЦТК [7]. Активность ферментов модулируется концентрацией субстрата, рН, аллостерическими эффекторами и обратимыми ковалентными модификациями в результате фосфорилирования/дефосфорилирования, взаимопревращения тиол-дисульфид, взаимодействия серосодержащих аминокислот белков-ферментов с NO [8].
Исследования дыхательного процесса выходят за рамки изучения только субстратов, катализирующих их ферментов, последовательности и направленности реакций. С помощью omics-технологий получены сведения о функционировании целых интерактомов, представляющих полный набор белок-белковых взаимодействий и “ченнелинга” метаболитов в отдельной клетке. Так, ферменты гликолиза фосфоглицератмутаза и енолаза образуют метаболон, связанный вне хлоропластов с триозофосфатным транслокатором, и вне митохондрий – с пируваткиназой и потенциалзависимым анионным каналом (VDAC) [16]. Образование такого интерактома не связано с проявлением специфической активности ферментов, а направлено на поддержание высокоэффективной координации хлоропластов и митохондрий и энергетического баланса клетки. В ЦТК выявлено 125 нековалентно связанных метаболонов, представляющих собой взаимодействия между ферментами цикла и 37 различными митохондриальными белками [17]. Эти исследования дают новые представления о пространственных аспектах функционирования ЦТК и его связи с метаболизмом аминокислот, синтезом АТФ, поддержанием редокс-статуса мЭТЦ, липидным обменом, сигналингом при стрессе.
Более 50% объема поверхности внутренней мембраны митохондрий занято белками, из них 35% – это белки-переносчики е– и протонов от НАД(Ф)·Н на О2 и АТФ-синтазы [18]. мЭТЦ включает четыре основных трансмембранных мультиферментных комплекса (I – IV) и два небольших по молекулярной массе компонента (убихинон и цитохром с), выполняющих функцию переноса е– между мультиферментными комплексами. Комплекс I (НАД·Н-дегидрогеназный) осуществляет перенос е- от НАД·Н к убихинону Q. Его мембранная часть содержит четыре Fe-S-центра. В состав периферической части входит ФМН-зависимая НАД·Н (убихинон-оксидоредуктаза), которая содержит три Fe-S-центра. Комплекс II (сукцинат-дегидрогеназный) катализирует окисление сукцината убихиноном с участием флавиновой (ФАД-зависимой) сукцинат : убихинон-оксидоредуктазы. В состав фермента входят три Fe-S-центра и цитохром b. Комплекс III (цитохром b/с1 − комплекс) переносит е– от восстановленного убихинона на цитохром с. Он содержит цитохромы b556 и b560, Fe-S белок Риске, цитохром с и центры связывания для убихинона. Комплекс IV (цитохром а/а3 − комплекс) является терминальным, переносит е– от цитохрома с к О2, в его состав входят цитохромы а и а3 (цитохромоксидаза или кислород-оксидоредуктаза) и два атома Cu. Комплексы I, III и IV образуют связанные с мембраной высокоупорядоченные суперкомплексы (респирасомы), что способствует ускорению тока е– вдоль мЭТЦ [19, 20].
Ток е- через комплексы I, III и IV обеспечивает энергией работу Н+-помп и образование протонного потенциала (ΔµН+), являющегося мерой свободной энергии, которая реализуется в синтезе АТФ с участием АТФ-синтазы (комплекс V).
Современные расчеты показывают, что при полном функционировании гликолиза, ЦТК и основного цитохромного пути в мЭТЦ количество образуемого АТФ на 1 моль окисленной глюкозы составляет 29 моль, а не 36, как считалось ранее, поскольку при сопряжении в комплексе IV прокачивается только два протона [21, 22]. В расчетах энергетической эффективности дыхания (ЭЭД) также учтены траты ΔµН+ для транспорта метаболитов через внутреннюю мембрану, утечка е– и другие потери.
мЭТЦ включает пути, не связанные с генерацией ΔµН+. К ним относят конкурирующие с комплексом I ротенон-нечувствительные дегидрогеназы, способные окислять НАД(Ф)·Н и восстанавливать пул убихинона [23, 24]. Активность альтернативных дегидрогеназ влияет на редокс-баланс клетки (соотношение НАД(Ф)+/НАД(Ф)·Н) и регулируется Са2+. Митохондриальное окисление 1 моля цитозольного НАД(Ф)·Н позволяет получить до 1.5 молей АТФ [22]. К системам, рассеивающим протонный градиент, относятся разобщающие белки (UCP), аналогичные UCP-белкам животных [24–26]. Полагают, что UCP поддерживают редокс-баланс в мЭТЦ, способствуя снижению скорости генерации АФК в митохондриях, регулируют скорость ЦТК, участвуют в транспорте метаболитов из митохондрий и, в целом, модулируют энергетический обмен.
Наиболее важным ферментом, обеспечивающим перенос е– по альтернативному нефосфорилирующему пути, является альтернативная цианидрезистентная оксидаза (АО). АО – терминальная убихинол:кислород-оксидоредуктаза, которая переносит е– с убихинона непосредственно на О2, минуя генерирующие электрохимический градиент протонов комплексы III и IV [27]. Вовлечение АО приводит к снижению ЭЭД. При транспорте е– по альтернативному пути (АП) величина YАТФ/глюкоза равна 11 моль АТФ/моль глюкозы [22], что на 62% меньше, чем по цитохромному пути. АО представляет собой гомодимер с мол. м. 32–37 кДа [28, 29]. Домены N-терминала мономеров АО ковалентно связаны дисульфидным мостиком остатков цистеина и ответственны за формирование димерной структуры. Восстановление дисульфидной связи в сульфгидрильную приводит к активации АО. Активность АО зависит от редокс-состояния фермента, восстановленности пула убихинона, содержания ключевых метаболитов (α-кетокислот), количества белка фермента и его изоформы [30, 31].
Показано участие АО в термогенезе ароидных [32], поддержании баланса между углеводным метаболизмом и скоростью электронного транспорта [30], регуляции скорости генерации АФК [33, 34], окислении образующегося в хлоропластах восстановителя [35], ретроградной сигнализации [4, 24, 36]. Экспрессия генов АО отмечена у прорастающих семян, формирующихся цветков и созревающих плодов, что свидетельствует об участии АП в процессах роста и развития [24, 37].
Активацию энергетически малоэффективного АП наблюдали при действии стресс-факторов (дефицит влаги, минеральных элементов, засоление и др.), вызывающих подавление основного (цитохромоксидазного) пути транспорта е– [38–41]. Вовлечение АП нередко сопровождалось активацией других энергодиссипирующих механизмов и антиоксидантной системы [42–46]. Это может указывать на участие АО в координации защитных механизмов клетки и регуляции генов стресс-ответа [34, 36, 47]. Современные данные подтверждают справедливость мнения О.А. Семихатовой [6] о том, что растения всегда имеют максимальную эффективность дыхания, а изменение показателей, характеризующих ЭЭД, лишь свидетельствует о перестройке их метаболического и энергетического баланса.
Особый интерес представляет вопрос о дыхании фотосинтезирующих клеток на свету. О.А. Семихатова неоднократно подчеркивала [6, 48], что для решения вопроса об ЭЭД растения необходимо учитывать снабжение энергетическими эквивалентами и метаболитами за счет фотосинтеза и наличие энергодиссипирующих дыхательных путей. Анализ накопленных к настоящему времени данных показывает, что дыхание на свету участвует в образовании и экспорте конечных продуктов фотосинтеза, контролирует уровень накопления восстановителей в клетке, защищает фотосинтетический аппарат от фотоингибирования, способствует сохранению гомеостаза клеток и нормальной жизнедеятельности растений [49–52]. Наблюдаемое снижение выделения СО2 при дыхании на свету связано с реорганизацией метаболизма фотосинтезирующей клетки [51, 53]. На свету цитозольный гликолиз ограничен, а ЦТК модифицирован в результате использования части метаболитов в азотном обмене и повышения содержания малата и фумарата. мЭТЦ остается функциональной, и в зависимости от условий (например, на интенсивном свету) транспорт е– может активироваться за счет вовлечения АО. Свет может влиять на экспрессию генов АО через фоторецепторную систему клетки [54]. Высокая освещенность и другие стрессы усиливают оксигеназную функцию РБФ-карбоксилазы и, следовательно, образование НАД·Н фотодыхательного происхождения, который окисляется в митохондриях с поглощением О2 [14]. АО участвует в окислении экспортируемых из хлоропластов восстановителей, тем самым “разгружая” фотосинтетическую ЭТЦ [34, 52].
Тот факт, что в процессе дыхания окисляется значительная часть фотосинтетически ассимилированного углерода, и за период вегетации растения переводят в СО2 количество дыхательного субстрата сопоставимое с накопленной биомассой, ставит вопрос о дыхательной цене процессов, обеспечивающих жизнедеятельность растений. Это важно в контексте разработки стратегий повышения урожайности основных продовольственных культур и сохранения природного биоразнообразия. Остановимся на основных компонентах и процессах формирования дыхательной цены.
У молодых интенсивно растущих растений (органов) подавляющая часть процесса дыхания (60–70%) связана с ростом. Согласно теоретическим расчетам [55] и имеющимся экспериментальным данным [2] из единицы дыхательного субстрата – глюкозы, которая используется в качестве источника С-скелетов и для генерации АТФ и НАД(Ф)·Н, синтезируется 0.75–0.85 единиц вегетативной биомассы. Трансформация субстрата в биомассу сопровождается выделением СО2 (дыхание на рост). Дыхательная цена синтеза биомассы зависит от ее состава, так как затраты на синтез белков и липидов примерно вдвое выше, чем на синтез углеводов или органических кислот. Поэтому эффективность роста клубней картофеля на 30% выше, чем листьев, содержащих на порядок меньше углеводов, но в 4 раза больше белков. Скорость дыхания растений и их органов коррелирует с относительной скоростью роста и снижается по мере затухания ростовых процессов. В зрелых растениях превалирует дыхание, связанное с поддержанием структурной целостности и функциональной активности клеток и тканей. Дыхательная цена поддержания варьирует в пределах от 0.02 до 0.04 г глюкозы/г сухой биомассы в сутки. Примерно 60% обусловлено затратами на обновление деградирующих компонентов биомассы (белков-ферментов, липидов, нуклеиновых кислот и др.), остальная часть связана с поддержанием ионных градиентов на мембранах и транспортом через мембраны метаболитов и ионов. По данным [56], от 7 до 55% дыхания зрелых листьев приходится на поддержание их донорной функции. Апопластная загрузка 1 г углерода, входящего в состав сахарозы, требует окисления в процессе дыхания количества глюкозы, содержащей 0.06 г углерода [57]. В ночное время зрелые листья Phaseolus vulgaris потребляли на апопластную загрузку флоэмы в среднем 30%, а листья Alocasia odora с симпластным типом загрузки только 10% от общего количества производимого дыханием АТФ [58]. При этом скорость образования АТФ в листьях фасоли была в 2 раза выше, чем у теневыносливой алоказии, и составляла в среднем 0.48 мкмоль АТФ/(м2 с).
Существенная часть поставляемой дыханием энергии связана с поглощением и ассимиляцией азота, так как потребность растений в азоте в 4–7 раз выше, чем в других элементах. Теоретически дыхательная цена поглощения 1 моля ${\text{NO}}_{3}^{ - }$ составляет порядка 0.43 моля О2, а ${\text{NH}}_{4}^{ + }$ вдвое меньше [57]. По разным оценкам, поглощение 1 г нитрата азота эквивалентно выделению 2–3 г СО2 и составляет около 40% всех дыхательных затрат корней [59]. Цена нетто-поглощения 1 моля ${\text{NO}}_{3}^{ - }$ у медленно растущих видов растений в 1.5–4 раза выше, чем у быстрорастущих [60]. По мнению авторов, это может быть обусловлено разницей в затратах на поддержание градиента ионов на мембранах и активацией нефосфорилирующих дыхательных путей.
Вовлечение АП и других митохондриальных энергодиссипирующих систем увеличивает затраты на рост и поддержание структурной целостности и функциональной активности уже существующей биомассы [22, 61]. Высказано мнение, что АП относится скорее к дыханию поддержания, нежели роста [62–64]. Вместе с тем, существуют данные об участии АП в поддержании активного роста, особенно при стрессе. Активация АП при засолении коррелировала с повышенным уровнем амино- и органических кислот у устойчивого к засолению генотипа Medicago truncatula TN6.18 [41] и в кластерных корнях Hakea prostrate при дефиците фосфора – с усилением синтеза и процессов экссудации карбоксилатов [65–67]. Активация АП в условиях гипоксии способствовала значительному усилению генерации АТФ в корнях Arabidopsis thaliana [62]. По мнению G.C. Vanlerberghe c соавт. [34], вовлечение АП в нестабильных и стрессовых условиях обеспечивает повышение устойчивости растений и более быстрое восстановление их роста после прекращения стресса. Однако и в оптимальных для роста условиях присутствие АП способствовало поддержанию редокс-баланса в мЭТЦ и сдерживанию образования АФК [62, 69].
Таким образом, основные дыхательные траты связаны с самим ростом и процессами, поддерживающими рост. Энергодиссипирующие системы, к которым относится АП, снижают эффективность преобразования углерода в биомассу, но являются неотъемлемой частью дыхательного процесса в норме и при стрессе [34, 69]. На современном этапе развития науки генно-инженерные стратегии повышения продуктивности направлены на сокращение потерь углерода в дыхании на поддержание, включающие “замедление ненужного оборота белка”, изменения направленности метаболических процессов, подавление активности нефосфорилирующих путей, увеличение эффективности ионного транспорта [70].
Итак, достижения последних десятилетий существенно расширили представления о вовлеченности процесса дыхания в метаболизм и биоэнергетику растительной клетки, роли дыхания в росте, продуктивности и устойчивости растений. Развитие молекулярно-генетических методов, omics-технологий и подходов системной биологии открывает новые возможности для более глубокого изучения организации и регуляции дыхания, нефосфорилирующих (энергетически малоэффективных) дыхательных путей, их значения для онтогенетического развития и формирования адаптивных реакций. Сбор и анализ информации о дыхании и эффективности дыхательных затрат дополнит характеристику свойств различных фенотипов, позволит раскрыть причины физиологической пластичности и проявления широкого спектра фенотипических модификаций у отдельных видов, генотипов и функциональных групп растений, оценить перспективы их использования в селекции и биотехнологии. Важное место должны занять исследования дыхания на экосистемном уровне в связи с биогенным круговоротом углерода и азота, возможным увеличением тепловой диссипации энергии в условиях повышения температуры среды и аридизации климата. Это будет способствовать пониманию глобального значения дыхания (от участия в сигналинге растительной клетки до функционирования экосистем), создаст базу для оптимизации дыхательных затрат, их соответствия потребностям растений в энергии и метаболитах, повышения продуктивности и устойчивости агро- и экосистем в условиях нестабильности климата.
Работа выполнена в рамках темы госбюджетных научных исследований “Фотосинтез, дыхание и биоэнергетика растений и фототрофных организмов (физиолого-биохимические, молекулярно-генетические и экологические аспекты)” (рег. № 122040600021-4).
Авторы заявляют об отсутствии конфликта интересов. Настоящая статья не содержит каких-либо исследований с участием людей и животных в качестве объектов исследований.
Список литературы
Amthor J.S. Respiration and Crop Productivity. New York: Springer-Verlag, 1989. 215 p.
Головко Т.К. Дыхание растений (физиологические аспекты). Санкт-Петербург: Наука, 1999. 204 с.
Gonzalez-Meler M.A., Taneva T. Integrated effects of atmospheric CO2 concentration on plant and ecosystem respiration // Plant respiration. Advances in photosynthesis and respiration / Eds. H. Lambers, M. Ribas-Carbo. Springer: Dordrecht. 2005. V. 18. P. 225. https://doi.org/10.1007/1-4020-3589-6_13
Van Aken O. Mitochondrial redox systems as central hubs in plant metabolism and signaling // Plant Physiol. 2021. V. 186. P. 36. https://doi.org/10.1093/plphys/kiab101
Crawford T., Lehotai N., Strand E. The role of retrograde signals during plant stress responses // J. Exp. Bot. 2018. V. 69. P. 2783. https://doi.org/10.1093/jxb/erx481
Семихатова О.А. Энергетика дыхания растений в норме и при экологическом стрессе. Ленинград: Наука, 1990. 72 с.
Семихатова О.А., Чиркова Т.В. Физиология дыхания раcтений. Санкт-Петербург: Издательство Санкт-Петербургского университета, 2001. 224 с.
Plaxton W., Podesta F.E. The functional organization and control of plant respiration // Crit. Rev. Plant Sci. 2006. V. 25. P. 159. https://doi.org/10.1080/07352680600563876
Plaxton W.C. The organization and regulation of plant glycolysis // Annu. Rev. Plant Physiol. Plant Mol. Biol. 1996. V. 47. P. 185. https://doi.org/10.1146/annurev.arplant.47.1.185
Stincone A., Prigione A., Cramer T., Wamelink M.M.C., Campbell K., Cheung E., Olin-Sandoval1 V., Grüning A.-M., Krüger A., Alam M.A., Keller M.A., Breitenbach M., Brindle K.M., Rabinowitz J.D., Ralser M. The return of metabolism: biochemistry and physiology of the pentose phosphate pathway // Biol. Rev. 2015. V. 90. P. 927. https://doi.org/10.1111/brv.12140
Moon S.J., Dong W., Stephanopoulos G.N., Sikes H.D. Oxidative pentose phosphate pathway and glucose anaplerosis support maintenance of mitochondrial NADPH pool under mitochondrial oxidative stress // Bioeng. Transl. Med. 2020. V. 5. e10184. https://doi.org/10.1002/btm2.10184
Fujiki Y., Ito M., Itoh T., Nishida I., Watanabe A. Activation of the promoters of Arabidopsis genes for the branched-chain α-keto acid dehydrogenase complex in transgenic tobacco BY-2 cells under sugar starvation // Plant Cell Physiol. 2002. V. 43. P. 275. https://doi.org/10.1093/pcp/pcf032
Schertl P., Cabassa C., Saadallah K., Bordenave M., Savour A., Braun H.-P. Biochemical characterization of proline dehydrogenase in Arabidopsis mitochondria // FEBS J. 2014. V. 281. P. 2794. https://doi.org/10.1111/febs.12821
Wingler A., Lea P.J., Quick W.P., Leegood R.C. Photorespiration: metabolic pathways and their role in stress protection // Philos. Trans. R. Soc. 2000. V. 355. P. 1517. https://doi.org/10.1098/rstb.2000.0712
Biochemistry and molecular biology of plants / Eds. B.B. Buchanan, W. Gruissem, R. Jones. Rockville, Md.: Amer. Soc. Plant Physiol., 2000. 1357 p.
Zhang Y., Sampathkumar A., Kerber S.M.-L., Swart C., Hille C., Seerangan K., Graf A., Sweetlove L., Fernie A.R. A moonlighting role for enzymes of glycolysis in the co-localization of mitochondria and chloroplasts // Nat. Commun. 2020. V. 11. P. 4509. https://doi.org/10.1038/s41467-020-18234-w
Zhang Y., Swart C., Alseekh S., Scossa F., Jiang L., Obata T., Graf A., Fernie A.R. The extra-pathway interactome of the TCA cycle: expected and unexpected metabolic interactions // Plant Physiol. 2018. V. 177. P. 966. https://doi.org/10.1104/pp.17.01687
Fuchs P., Rugen N., Carrie C., Elsӓsser M., Finkemeier I., Giese J., Hildebrandt T.M., Kühn K., Maurino V.G., Ruberti C., Schallenberg-Rüdinger M., Steinbeck J., Braun H.P., Eubel H., Meyer E.H., Müller-Schüssele S.J., Schwarzlӓnder M. Single organelle function and organization as estimated from Arabidopsis mitochondrial proteomics // Plant J. 2020. V. 101. P. 420. https://doi.org/10.1111/tpj.14534
Ramírez-Aguilar S.J., Keuthe M., Rocha M., Fedyaev V.V., Kramp K., Gupta K.J., Rasmusson A.G., Schulze W.X., Van Dongen J.T. The composition of plant mitochondrial supercomplexes changes with oxygen availability // J. Biol. Chem. 2011. V. 286. P. 43045. https://doi.org/10.1074/jbc.M111.252544
Rakhmankulova Z.F. Respiratory supercomplexes of plant mitochondria: structure and possible functions // Russ. J. Plant Physiol. 2014. V. 61. P. 721. https://doi.org/10.1134/S1021443714060168
Amthor J.S. Respiration and carbon assimilate use // Physiology and determination of crop yield / Eds. K.J. Boote, J.M. Bennett, T.R. Sinclair, G.M. Paulsen. American Society of Agronomy, Crop Science Society of America, Soil Science Society of America. 1994. P. 221. https://doi.org/10.2134/1994.physiologyanddetermination.c15
Amthor J.S. The McCree-de Wit-Penning de Vries-Thornley respiration paradigms: 30 years later // Ann. Bot. 2000. V. 86. P. 1. https://doi.org/10.1006/anbo.2000.117
Rasmusson A.J., Geisler D.A., Møller I.M. The multiplicity of dehydrogenases in the electron transport chain of plant mitochondria // Mitochondrion. 2008. V. 8. P. 47. https://doi.org/10.1016/j.mito.2007.10.004
Pu X., Lvy X., Tan T., Fu F., Qin G., Lin H. Roles of mitochondrial energy dissipation systems in plant development and acclimation to stress // Ann. Bot. 2015. V. 116. P. 583. https://doi.org/10.1093/aob/mcv063
Sluse F.E, Jarmuszkiewicz W. Uncoupling proteins outside the animal and plant kingdoms: functional and evolutionary aspects // FEBS Lett. 2002. V. 510. P. 117. https://doi.org/10.1016/S0014-5793(01)03229-X
Vercesi A.E., Borecky J.M.I.G., Arruda P., Cuccovia I.M., Chaimovich H. Plant uncoupling mitochondrial proteins // Annu. Rev. Plant Biol. 2006. V. 57. P. 383. https://doi.org/10.1146/annurev.arplant.57.032905.10533
Vanlerberghe G.C., McIntosh L. Alternative oxidase: from gene to function // Annu. Rev. Plant Physiol. Plant Mol. Biol. 1997. V. 48. P. 703. https://doi.org/10.1146/annurev.arplant.48.1.703
Andersson M.E., Nordlund P. A revised model of the active site of the alternative oxidase // FEBS Lett. 1999. V. 449. P. 17. https://doi.org/10.1016/s0014-5793(99)00376-2
Albury M.S., Elliott C., Moore A.L. Towards a structural elucidation of the alternative oxidase in plants // Physiol. Plant. 2009. V. 137. P. 316. https://doi.org/10.1111/j.1399-3054.2009.01270.x
Millenaar F.F., Lambers H. The alternative oxidase: in vivo regulation and function // Plant Biol. 2003. V. 5. P. 2. https://doi.org/10.1055/s-2003-37974
Selinski J., Hartmann A., Deckers-Hebestreit G., Day D.A., Whelan J., Scheibe R. Alternative oxidase isoforms are differentially activated by tricarboxylic acid cycle intermediates // Plant Physiol. 2018. V. 76. P. 1423. https://doi.org/10.1104/pp.17.01331
Meeuse B.J.D. Thermogenic respiration in aroids // Annu. Rev. Plant Physiol. 1975. V. 26. P. 117. https://doi.org/10.1146/annurev.pp.26.060175.001001
Maxwell D.P., Wang Y., McIntosh L. The alternative oxidase lowers mitochondrial reactive oxygen production in plant cells // Proc. Natl. Acad. Sci. U.S.A. 1999. V. 96. P. 8271. https://doi.org/10.1073/pnas.96.14.8271
Vanlerberghe G.C., Dahal K., Alber N.A., Chadee A. Photosynthesis, respiration and growth: a carbon and energy balancing act for alternative oxidase // Mitochondrion. 2020. V. 52. P. 197. https://doi.org/10.1016/j.mito.2020.04.001
Dinakar C., Raghavendra A.S., Padmasree K. Importance of AOX pathway in optimizing photosynthesis under high light stress: role or pyruvate and malate in activating AOX // Physiol. Plant. 2010. V. 139. P. 13. https://doi.org/10.1111/j.1399-3054.2010.01346.x
Garmash E.V. Signal pathways for regulation of plant alternative oxidase genes’ expression // Russ. J. Plant Physiol. 2022. V. 69. P. 3. https://doi.org/10.1134/S1021443722010058
Clifton R., Lister R., Parker K.L., Sappl P.G., Elhafez D., Millar A.H., Day D.A., Whelan J. Stress-induced co-expression of alternative respiratory chain components in Arabidopsis thaliana // Plant Mol. Biol. 2005. V. 58. P. 193. https://doi.org/10.1007/s11103-005-5514-7
Dahal K., Vanlerberghe G.C. Alternative oxidase respiration maintains both mitochondrial and chloroplast function during drought // New Phytol. 2017. V. 213. P. 560. https://doi.org/10.1111/nph.14169
Ribas-Carbo M., Aroca R., Gonzàlez-Meler M.A., Irigoyen J.J., Sánchez-Dìaz M. The electron partitioning between the cytochrome and alternative respiratory pathways during chilling recovery in two cultivars of maize differing in chilling sensitivity // Plant Physiol. 2000. V. 122. P. 199. https://doi.org/10.1104/pp.122.1.199
Gonzàlez-Meler M.A., Giles L., Thomas R.B., Siedow J.N. Metabolic regulation of leaf respiration and alternative pathway activity in response to phosphate supply // Plant Cell Environ. 2001. V. 24. P. 205. https://doi.org/10.1111/j.1365-3040.2001.00674.x
Del-Saz N.F., Clemente-Moreno M.J., Mhadhbi H., Flexas J., Fernie A.R., Ribas-Carbó M. Salinity tolerance is related to cyanide-resistant alternative respiration in Medicago truncatula under sudden severe stress // Plant Cell Environ. 2016. V. 39 P. 2361. https://doi.org/10.1111/pce.12776
Szal B., Lukawska K., Zdolinska I., Rychter A.M. Chilling stress and mitochondrial genome rearrangement in the MSC16 cucumber mutant affect the alternative oxidase and antioxidant defense system to a similar extent // Physiol. Plant. 2009. V. 137. P. 435. https://doi.org/10.1111/j.1399-3054.2009.01255.x
Grabelnych O.I., Borovik O.A., Tauson E.L., Pobezhimova T.P., Katyshev A.I., Pavlovskaya N.S., Koroleva N.A., Lyubushkina I.V., Bashmakov V.Yu., Popov V.N., Borovskii G.B., Voinikov V.K. Mitochondrial energy-dissipating systems (alternative oxidase, uncoupling proteins, and external NADH dehydrogenase) are involved in development of frost-resistance of winter wheat seedlings // Biochemistry (Moscow). 2014. V. 79. P. 506. https://doi.org/10.1134/S0006297914060030
Vishwakarma A., Tetali S.D., Selinski J., Scheibe R., Padmasree K. Importance of the alternative oxidase (AOX) pathway in regulating cellular redox and ROS homeostasis to optimize photosynthesis during restriction of the cytochrome oxidase pathway in Arabidopsis thaliana // Ann. Bot. 2015. V. 116. P. 555. https://doi.org/10.1093/aob/mcv122
Garmash E.V., Velegzhaninov I.O., Grabelhych O.I., Borovik O.A., Silina E.V., Voinikov V.K., Golovko T.K. Expression profiles of genes for mitochondrial respiratory energy-dissipating systems and antioxidant enzymes in wheat leaves during de-etiolation // J. Plant Physiol. 2017. V. 215. P. 110. https://doi.org/10.1016/j.jplph.2017.05.023
Garmash E.V., Velegzhaninov I.O., Ermolina K.V., Rybak A.V., Malyshev R.V. Altered levels of AOX1a expression result in changes in metabolic pathways in Arabidopsis thaliana plants acclimated to low dose rates of ultraviolet B radiation // Plant Sci. 2020. V. 291. P. 110332. https://doi.org/10.1016/j.plantsci.2019.110332
Van Aken O., Giraud E., Clifton R., Whelan J. Alternative oxidase: a target and regulator of stress responses // Physiol. Plant. 2009. V. 137. P. 354. https://doi.org/10.1111/j.1399-3054.2009.01240.x
Семихатова О.А., Заленский О.В. Сопряженность процессов фотосинтеза и дыхания // Физиология фотосинтеза / Под ред. Ничипоровича А.А. Москва: Наука, 1982. С. 130.
Krömer S. Respiration during photosynthesis // Annu. Rev. Plant Physiol. Plant Mol. Biol. 1995. V. 46. P. 45. https://doi.org/10.1146/annurev.pp.46.060195.000401
Nunes-Nesi A., Sweetlove L.J., Fernie A.R. Operation and function of the tricarboxylic acid cycle in the illuminated leaf // Physiol. Plant. 2007. V. 129. P. 45. https://doi.org/10.1111/j.1365-3040.2011.02332.x
Tcherkez G., Gauthier P., Buckley T.N., Busch F.A., Barbour M.M., Bruhn D., Heske M.A., Gon H.Y., Crous K.J., Griffin K., Way D., Turnbull M., Adams M.A., Atkin O.K., Farquhar G.D., Cornic G. Leaf day respiration: low CO2 flux but high significance for metabolism and carbon balance // New Phytol. 2017. V. 216. P. 986. https://doi.org/10.1111/nph.14816
Garmash E.V. Mitochondrial respiration of the photosynthesizing cell // Russ. J. Plant Physiol. 2016. V. 63. P. 13. https://doi.org/10.1134/S1021443715060072
Harry V., Igamberdiev A.U., Keerberg O., Pӓrnik T., Atkin O., Zaragoza-Castells J., Gardeström P. Respiration in photosynthetic cells: gas exchange components, interaction with photorespiration and the operation of mithochondria in the light // Plant respiration: from cell to ecosystem / Eds. H. Lambers, M. Ribas-Carbo. Springer: Dordrecht. 2005. P. 43. http://hdl.handle.net/ 1885/26829
Zhang D.-W., Xu F., Zhang Z.-W., Chen Y.-E., Du J.-B., Jia S.-D., Yuan S., Lin H.-H. Effects of light on cyanide-resistant respiration and alternative oxidase function in Arabidopsis seedlings // Plant Cell Environ. 2010. V. 33. P. 2121. https://doi.org/10.1111/j.1365-3040.2010.02211.x
Penning de Vries F.W.T., Brunsting A.H.M., Van Laar H.H. Products, requirements and efficiency of biosynthesis: a quantitative approach // J. Theor. Biol. 1974. V. 45. P. 339. https://doi.org/10.1016/0022-5193(74)90119-2
Bouma T.J., De Visser R., Van Leeuwen P.H., De Kock M.J., Lambers H. The respiratory energy requirements involved in nocturnal carbohydrate export from starch-storing mature source leaves and their contribution to leaf dark respiration // J. Exp. Bot. 1995. V. 46. P. 1185.
Cannell M.G.R., Thornley J.H.M. Modelling the components of plant respiration: some guiding principles // Ann. Bot. 2000. V. 85. P. 45. https://doi.org/10.1006/anbo.1999.0997
Noguchi K., Go C.-S., Terashima I., Ueda S., Noguchi T.Y. Activities of the cyanide-resistant respiratory pathway in leaves of sun and shade species // Aust. J. Plant Physiol. 2001. V. 28. P. 27. https://doi.org/10.1071/PP00056
Szaniawski R.K., Kielkiewicz M. Maintenance and growth respiration in shoots and roots of sunflower plants grown at different root temperatures // Physiol. Plant. 1982. V. 54. P. 500. https://doi.org/10.1111/j.1399-3054.1982.tb00716.x
Scheurwater I., Clarkson D.T., Purves J.V., Van Rijt G., Saker L.R., Welschen R., Lambers H. Relatively large nitrate efflux can account for the high specific respiratory costs for nitrate transport in slow-growing grass species // Plant Soil. 1999. V. 215. P. 123. https://doi.org/10.1023/A:1004559628401
Thornley J.H.M. Energy, respiration, and growth in plants // Ann. Bot. 1971. V. 35. P. 721. https://doi.org/10.1093
Florez-Sarasa I., Bouma T.J., Medrano H., Azcon-Bieto J., Ribas-Carbo M. Contribution of the cytochrome and alternative pathways to growth respiration and maintenance respiration in Arabidopsis thaliana // Physiol. Plant. 2007. V. 129. P. 143. https://doi.org/10.1111/j.1399-3054.2006.00796.x
Priault P., Vidal G., De Paepe R., Ribas-Carbo M. Leaf age-related changes in respiratory pathways are dependent on complex I activity in Nicotiana sylvestris // Physiol. Plant. 2007. V. 129. P. 152. https://doi.org/10.1111/j.1399-3054.2006.00836.x
Rakhmankulova Z.F., Fedyaev V.V., Podashevka O.A., Usmanov I.Yu. Alternative respiration pathways and secondary metabolism in plants with different adaptive strategies under mineral deficiency // Russ. J. Plant Physiol. 2003. V. 50. P. 206. https://doi.org/10.1023/A:1022973130775
Shane M.W., Cramer M.D., Funayama-Noguchi S., Cawthray G.R., Millar A.H., Day D.A., Lambers H. Developmental physiology of cluster-root carboxylate synthesis and exudation in Harsh hakea. Expression of phosphoenolpyruvate carboxylase and the alternative oxidase // Plant Physiol. 2004. V. 135. P. 549. https://doi.org/10.1104/pp.103.035659
Florez-Sarasa I., Lambers H., Wang X., Finnegan P.M., Ribas-Carbo M. The alternative respiratory pathway mediates carboxylate synthesis in white lupin cluster roots under phosphorus deprivation // Plant Cell Environ. 2014. V. 37. P. 922. https://doi.org/10.1111/pce.12208
Del-Saz N.F., Romero-Munar A., Cawthray G.R., Aroca R., Baraza E., Flexas J., Ribas-Carbo M. Arbuscular mycorrhizal fungus colonization in Nicotiana tabacum decreases the rate of both carboxylate exudation and root respiration and increases plant growth under phosphorus limitation // Plant Soil. 2017. V. 416. P. 97. https://doi.org/10.1007/s11104-017-3188-y
Vishwakarma A., Kumari A., Mur L.A.J., Gupta K.J. A discrete role for alternative oxidase under hypoxia to increase nitric oxide and drive energy production // Free Radic. Biol. Med. 2018. V. 122. P. 40. https://doi.org/10.1016/j.freeradbiomed.2018.03.045
Del-Saz N.F., Ribas-Carbo M., McDonald A.E., Lambers H., Fernie A.R., Florez-Sarasa I. An in vivo perspective of the role(s) of the alternative oxidase pathway // Trends Plant Sci. 2018. V. 23. P. 206. https://doi.org/10.1016/j.tplants.2017.11.006
Amthor J.S., Bar-Even A., Hanson A.D., Millar A.H., Stitt M., Sweetlove L.J., Tyerman S.D. Engineering strategies to boost crop productivity by cutting respiratory carbon loss // Plant Cell. 2019. V. 31. P. 297. https://doi.org/10.1105/tpc.18.00743
Дополнительные материалы отсутствуют.
Инструменты
Физиология растений