

Генетика, 2020, T. 56, № 1, стр. 55-66
Сравнительный анализ генетической и морфолого-анатомической изменчивости можжевельника обыкновенного (Juniperus communis L.)
С. Г. Князева 1, *, Е. В. Хантемирова 2, **
1 Федеральный исследовательский центр Сибирского отделения
Российской академии наук, Институт леса им. В.Н. Сукачева
660049 Красноярск, Россия
2 Институт экологии растений и животных Уральского отделения
Российской академии наук
620144 Екатеринбург, Россия
* E-mail: knyazevas@mail.ru
** E-mail: hantemirova@mail.ru
Поступила в редакцию 05.02.2019
После доработки 21.03.2019
Принята к публикации 09.04.2019
Аннотация
Генетическими и анатомо-морфологическими методами проведено комплексное изучение биологического разнообразия 27 природных популяций Juniperus communis L. на основном ареале. Подтвержден таксономический статус кавказской разновидности J. communis var. оblonga. В свою очередь, популяции этой разновидности на Северном Кавказе разделились по хлоропластным маркерам на две части – западную и восточную. Североамериканский J. communis var. depressa и дальневосточный J. сommunis var. saxatilis не отличаются по морфолого-анатомическим и ядерным маркерам, но различаются по хлоропластным, и, вероятно, являются родственными генетическими линиями, которые в сходных экологических условиях приобрели внешнее конвергентное сходство. Отличить их можно только генетическими методами. J. сommunis var. сommunis и J. сommunis var. saxatilis, растущие в Европе и Азии, не дифференцируются генетически, но имеют статистически значимые отличия по ряду морфологических и анатомических параметров хвои и преобладающей жизненной форме. Поэтому кажется вероятным, что данные разновидности можжевельника являются экотипами, которые приобрели отличия в морфологии и анатомии, произрастая в разных экологических условиях.
Главными факторами, влияющими на распределение биологической изменчивости внутри и между популяциями видов растений, являются их географическое распространение, система размножения и опыления, способ рассеивания семян. Не последнюю роль играют эволюционная история видов, периодические изменения численности их популяций, направления миграций, уменьшение и расширение ареалов, которые происходили в результате климатических флуктуаций [1–3]. С другой стороны, некоторые эколого-морфологические свойства видов (устойчивость к низким температурам, например) могут отражаться на выживаемости видов во время ледниковых периодов и скорости их послеледниковых экспансий [4].
Объект нашего исследования – можжевельник обыкновенный J. communis L. – вечнозеленый хвойный вид из семейства кипарисовых, имеющий обширный циркумбореальный ареал. Благодаря морфологической пластичности и широкой экологической амплитуде, этот вид распространен не только в умеренной зоне – горах Европы и Азии и в подлесках хвойных лесов, но и смог расселиться далеко на север, в арктические горные тундры. Обладая свойствами холодостойкости и ксероморфности, а также способностью образовывать стланиковую форму, можжевельник обыкновенный, по всей видимости, способен был переживать ледниковые периоды не только в южных, но и в северных рефугиумах [5]. Важное эволюционное приобретение J. communis – своеобразные “плоды” в виде сочных шишкоягод, которые распространяются птицами и мелкими млекопитающими, позволяло ему быстро расселяться в межледниковые периоды. А свойство ветроопыляемости способствовало перемешиванию генных потоков. Сочетание этих свойств свидетельствует о большом эволюционном потенциале можжевельника и способности его успешно справляться с климатическими изменениями.
Широкое географическое распространение Juniperus communis L. при большом экологическом диапазоне вызвало образование разнообразных жизненных форм (от стлаников до деревьев). С высокой изменчивостью морфологических признаков этого вида связаны и трудности в подвидовой таксономической классификации. Это весьма полиморфный вид с запутанной и часто спорной таксономией [6, 7]. Форма роста наряду с признаками хвои являются основными критериями для выделения синтаксонов. Так, Альйос Фарджун (A. Farjon) [6] выделяет в Евразии две разновидности можжевельника обыкновенного: J. сommunis L. var. communis (форма прямостоячего или раскидистого кустарника, или дерева) и J. communis var. saxatilis Pall. (форма стелющегося или приподнимающегося кустарника). Последняя разновидность имеет обширную синонимику в ранге вида, подвида, разновидности и формы (J. nana Willd., J. alpinа S.F. Gray, J. pygmaea C. Koch, J. communis var. saxatilis Pall., J. communis f. sibirica Rydb., J. communis f. nana Loud.) [8–11]. В России для этой разновидности принято название J. sibirica – сибирский можжевельник. Обе разновидности имеют сходные морфологические признаки и перекрывающийся ареал на территории России, охватывающий Европейскую часть России, Урал, Сибирь. Фактически не существует ни четких границ ареалов этих разновидностей, ни четких морфологических различий между ними [12–15].
На Кавказе произрастает другая разновидность можжевельника обыкновенного – J. communis var. оblonga hort. ex Loudon. (=J. oblonga Bieb., J. albanica Penzes, J. communis var. caucasica Endl., J. communis var. saxatilis Pall.) [16–18]. Н.Н. Имханицкая [19] поддерживает мнение А.Н. Галушко о подвидовом статусе этого таксона: J. communis subsp. оblonga (Bieb) Galushko. А. Фарджун [6] не признает выделения этого можжевельника даже в качестве разновидности, хотя он имеет достаточно четкие морфологические отличия; в то же время выделяет в качестве отдельной североамериканской разновидности J. communis var. depressa, не имеющего четких морфологических отличий от евроазиатского J. communis var. saxatilis.
Возможно такая неопределенность в трактовке внутривидовых категорий J. communis связана и со сложной филогеографической историей, обнаруженной у этого вида с помощью генетических маркеров [5]. Проблему таких таксономически сложных групп видов могут решить комплексные исследования с применением методов классической морфометрии и молекулярной биологии.
Цель нашей работы – провести комплексное исследование изменчивости природных популяций можжевельника обыкновенного в широком географическом масштабе с помощью анатомо-морфологических и генетических методов – алозимным методом [20] и путем анализа изменчивости хлоропластной (хлДНК) [5], чтобы выявить закономерности, характеризующие вид и его разновидности на протяжении всего ареала.
МАТЕРИАЛЫ И МЕТОДЫ
Нами изучены 27 природных популяций Juniperus communis L., произрастающих на территории Европы, Азии и Северной Америки (рис. 1, табл. 1). Популяции из Томска, Подмосковья и Таллина имеют форму прямостоячего кустарника и скорее всего представляют собой J. сommunis L. var. communis. Можжевельник из Аляски является разновидностью J. communis var. depressa. Кавказские популяции – это, вероятно, J. communis var. оblonga. Остальные популяции вероятно относятся к самой распространенной на территории Евразии вариации: J. communis var. saxatilis.
Рис. 1.
Места сбора популяций и схема ареалов вариаций можжевельника обыкновенного ( – J. сommunis var. saxatilis;
– J. сommunis var. saxatilis;  – J. communis var. communis;
– J. communis var. communis;  – J. communis var. oblonga,
– J. communis var. oblonga,  – J. communis var. depressa).
– J. communis var. depressa).
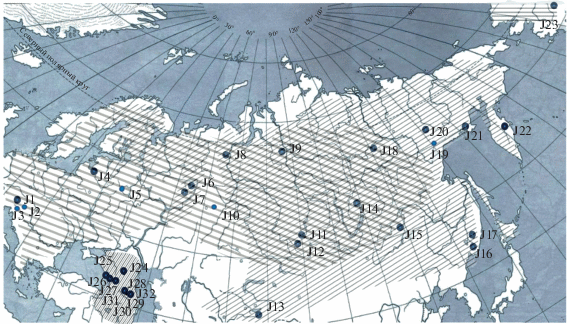
Таблица 1.
Популяции Juniperus communis
| № | Популяция | Широта | Долгота | Высота над ур. м., м |
|---|---|---|---|---|
| J1 | Альпы (Сев. Тироль) | N 47°11′ | E 12°0′ | 1700 |
| J2 | Восточные Альпы | N 47°04′ | E 14°34′ | 1800 |
| J3 | Южные Альпы | N 46°30′ | E 14°45′ | 1759 |
| J4 | Эстония | N 57°27′ | E 24°52′ | 22 |
| J5 | Подмосковье | N 56°50′ | E 36°55′ | 141 |
| J6 | Таганай | N 55°10′ | E 59°40′ | 1000 |
| J7 | Машак | N 54°19′ | E 58°06′ | 669 |
| J8 | Полярный Урал | N 66°50′ | E 65°40′ | 250 |
| J9 | Туруханск | N 65°48′ | E 87°59′ | 40 |
| J10 | Томск | N 56°70′ | E 84°76′ | 133 |
| J11 | Ергаки | N 53°08′ | E 92°56′ | 1750 |
| J12 | Горная Шория | N 52°55′ | E 88°00′ | 1570 |
| J13 | Тянь-Шань | N 43°06′ | E 77°04′ | 3000 |
| J14 | Северобайкальск | N 55°42′ | E 109°04′ | 536 |
| J15 | Газимур | N 52°09′ | E 119°2′ | 936 |
| J16 | Лазо | N 43°31′ | E 134°06′ | 1287 |
| J17 | Сихотэ-Алинь | N 45°00′ | E 136°30′ | 842 |
| J18 | Якутия | N 63°28′ | E 120°3′ | 120 |
| J19 | Колыма | N 63°25′ | E 140°36′ | 120 |
| J20 | Оймякон | N 63°03′ | N 138°09′ | 193 |
| J21 | Магадан | N 59°35′ | E 150°4′ | 118 |
| J22 | Камчатка | N 56°01′ | E 161°11′ | 50 |
| J23 | Аляска | N 64°50′ | E 147°4′ | 135 |
| J24 | Ногайская степь | N 44°04′ | E 45°35′ | 60 |
| J25 | Сочи | N 43°42′ | E 40°10′ | 1000 |
| J26 | Архыз | N 43°34′ | E 41°16′ | 2034 |
| J27 | Нальчик | N 43°08′ | E 43°29′ | 1470 |
| J28 | Гуниб | N 42°24′ | E 46°54′ | 1850 |
| J29 | Талги | N 42°62′ | E 47°26′ | 458 |
| J30 | Теберда | N 43°22′ | E 41°43′ | 1700 |
| J31 | Цудахар | N 42°19′ | E 47°09′ | 1200 |
| J32 | Губден | N 42°32′ | E 47°26′ | 870 |
Проводилось исследование анатомо-морфологической изменчивости параметров хвои (длина, ширина, длина кончика, высота и толщина среза, периметр и площадь среза, периметр и площадь смоляного хода и проводящего пучка, ширина устьичной полоски, число устьичных полосок, толщина покровных тканей, число устьиц и их размеры, число обкладочных клеток, коэффициент Адамса, высота, ширина и форма киля).
Образцы хвои выдерживали в спирте, затем делали поперечные срезы лезвием. С каждой особи брали по пять хвоинок четырех возрастов. Срезы помещали в каплю глицерина и фотографировали с помощью микроскопа Микромед-Р1 с камерой DCM500. Измерения проводились с помощью программы Scopephoto 3.0. Полученные данные обрабатывались статистическими методами.
Также изучена генетическая изменчивость с помощью ядерных (аллозимный анализ, 23 популяции) и хлоропластных (PCR-RFLP) маркеров (30 популяций). Для аллозимного анализа подготовку проб, электрофорез белков и гистохимическое окрашивание выявленных зон ферментативной активности проводили в соответствии с методиками [21, 22], описанными подробно в наших предыдущих работах [20, 23]. Для оценки генетического разнообразия растений использованы восемь локусов (Fdh, Idh, Pgi-A, Pgi-B, 6-Pgd-A, 6‑Pgd-B, Sod-A, Sod-B). Генетические расстояния (D) между популяциями определялись с помощью генетических дистанций Nei [24]. Матрицы значений D были использованы для ординации популяций с применением анализа главных координат (с использованием NTSYS-pc). Данные анализировались с использованием программ BIOSYS-2 [25] и NTSYS-pc [26].
Хлоропластную ДНК экстрагировали из свежей или высушенной в силикагеле хвои можжевельника с помощью СТАБ-метода [27] и использовали ее, как темплату для ПЦР-амплификации двух фрагментов хпДНК: trnT-trnF регион с праймерами a, b, c, d [28] и 16S-trnA регион с праймерами 5'-gaaggctgggctagtgactg-3' and 5'-gagataagcggactcgaacc-3' (Семериков, неопубликованное) и дополнительным внутренним праймером 5'-gaaaccaatgccataagcaa-3'. ПЦР-продукты были разрезаны рестриктазой TaqI для 16S-trnA региона и HinfI для trnT-trnF региона. Продукты рестрикции были разделены индивидуально в 6%-ном денатурирующем полиакриламидном геле с последующим окрашиванием серебром. Для каждого фрагмента все гаплотипы, идентифицированные с помощью RFLP, были секвенированы методом Сэнгера. Подробнее методика PCR-RFLP и секвенирование описаны в нашей предыдущей работе [5].
РЕЗУЛЬТАТЫ
Морфолого-анатомический анализ
Исследование морфологии и анатомии хвои показывает высокую пластичность ее анатомо-морфологических признаков. Кавказские популяции резко отличаются от остальных по большинству изученных признаков. Самая длинная хвоя наблюдается у можжевельника из Ногайской степи – 22 мм, самая широкая – у растений из Гунибского плато – 2.3 мм. Самые высокие средние значения параметров хвои характерны для можжевельника из Гунибской популяции (табл. 2). Растения с плато Гуниб являются уникальными и отличаются значимо по многим признакам (высота и толщина хвои, площадь поперечного разреза, периметр проводящего пучка, высота киля) от всех других популяций, в том числе кавказских. В то же время группа очень неоднородная: с высокой межпопуляционной изменчивостью.
Таблица 2.
Количественные показатели исследованных признаков (Xср ± SX)
| Популяция | Длина хвои, см | Длина кончика хвои, мм | Ширина хвои, мм | Высота хвоинки, мм | Площадь разреза, мм2 |
| Альпы | 8.52 ± 1.9 | 0.37 ± 0.08 | 1.5 ± 0.34 | 0.5 ± 0.11 | 0.47 ± 0.1 |
| Таллин | 13.7 ± 2.7 | 0.5 ± 0.1 | 1.22 ± 0.24 | 0.4 ± 0.07 | 0.29 ± 0.05 |
| Подмосковье | 11.7 ± 2.6 | 0.4 ± 0.09 | 1.31 ± 0.29 | 0.43 ± 0.1 | 0.31 ± 0.07 |
| Таганай | 8.8 ± 1.9 | 0.5 ± 0.11 | 1.3 ± 0.29 | 0.44 ± 0.1 | 0.32 ± 0.07 |
| Машак | 8.01 ± 1.7 | 0.39 ± 0.09 | 1.45 ± 0.32 | 0.5 ± 0.11 | 0.38 ± 0.09 |
| Пол. Урал | 5.9 ± 1.3 | 0.29 ± 0.07 | 1.36 ± 0.3 | 0.44 ± 0.1 | 0.34 ± 0.08 |
| Туруханск | 7.66 ± 1.7 | 0.49 ± 0.11 | 1.27 ± 0.28 | 0.38 ± 0.08 | 0.3 ± 0.07 |
| Томск | 13.6 ± 3.05 | 0.62 ± 0.14 | 1.3 ± 0.29 | 0.38 ± 0.08 | 0.27 ± 0.06 |
| Ергаки | 8.11 ± 1.8 | 0.402 ± 0.09 | 1.34 ± 0.3 | 0.46 ± 0.1 | 0.36 ± 0.08 |
| Горная Шория | 5.36 ± 1.2 | 0.25 ± 0.06 | 1.27 ± 0.28 | 0.46 ± 0.1 | 0.34 ± 0.08 |
| Тянь-Шань | 8.29 ± 1.8 | 0.48 ± 0.11 | 1.45 ± 0.33 | 0.5 ± 0.11 | 0.38 ± 0.09 |
| Северобайкальск | 9.9 ± 2.2 | 0.42 ± 0.09 | 1.32 ± 0.29 | 0.43 ± 0.09 | 0.33 ± 0.07 |
| Газимур | 12.7 ± 2.8 | 0.54 ± 0.12 | 1.33 ± 0.3 | 0.51 ± 0.12 | 0.46 ± 0.1 |
| Лазо | 10.8 ± 2.4 | 0.47 ± 0.11 | 1.36 ± 0.3 | 0.43 ± 0.1 | 0.35 ± 0.08 |
| Сихотэ-Алинь | 11.1 ± 2.5 | 0.42 ± 0.09 | 1.28 ± 0.29 | 0.41 ± 0.09 | 0.33 ± 0.07 |
| Якутия | 7.2 ± 1.5 | 0.4 ± 0.08 | 1.12 ± 0.22 | 0.38 ± 0.08 | 0.25 ± 0.05 |
| Колыма | 8.3 ± 1.8 | 0.36 ± 0.08 | 1.23 ± 0.28 | 0.42 ± 0.09 | 0.36 ± 0.08 |
| Оймякон | 9.1 ± 2.03 | 0.52 ± 0.12 | 1.22 ± 0.27 | 0.46 ± 0.1 | 0.37 ± 0.08 |
| Магадан | 9.9 ± 2.2 | 0.41 ± 0.09 | 1.2 ± 0.27 | 0.39 ± 0.09 | 0.28 ± 0.06 |
| Камчатка | 7.8 ± 1.1 | 0.37 ± 0.05 | 1.36 ± 0.19 | 0.43 ± 0.14 | 0.41 ± 0.06 |
| Аляска | 10.8 ± 1.2 | 0.31 ± 0.06 | 1.35 ± 0.25 | 0.5 ± 0.09 | 0.38 ± 0.07 |
| Ногайская степь | 15.2 ± 3 | 0.74 ± 0.15 | 1.67 ± 0.33 | 0.59 ± 0.11 | 0.56 ± 0.11 |
| Сочи | 9.6 ± 1.9 | 0.5 ± 0.1 | 1.65 ± 0.33 | 0.58 ± 0.11 | 0.52 ± 0.1 |
| Архыз | 12.8 ± 2.5 | 0.58 ± 0.11 | 1.61 ± 0.32 | 0.56 ± 0.11 | 0.56 ± 0.11 |
| Нальчик | 12.3 ± 2.7 | 0.62 ± 0.14 | 1.7 ± 0.38 | 0.61 ± 0.14 | 0.56 ± 0.13 |
| Гуниб | 14.2 ± 2.8 | 0.96 ± 0.2 | 1.85 ± 0.37 | 0.76 ± 0.15 | 0.76 ± 0.15 |
| Талги | 13.2 ± 2.6 | 0.85 ± 0.17 | 1.58 ± 0.32 | 0.58 ± 0.11 | 0.57 ± 0.11 |
| Популяция | S смоляного хода, мм2 | S проводящего пучка, мкм2 | Число обкладочных клеток | Число полосок | Ка |
| Альпы | 0.016 ± 0.003 | 8006.4 ± 1790 | 2.9 ± 0.7 | 1.37 ± 0.31 | 2.54 ± 0.57 |
| Таллин | 0.009 ± 0.0001 | 5348.9 ± 1069.7 | 17.8 ± 3.6 | 2 ± 0.4 | 2.51 ± 0.5 |
| Подмоск. | 0.0105 ± 0.002 | 6215.7 ± 1389.9 | 1.85 ± 0.41 | 1.46 ± 0.33 | 1.66 ± 0.37 |
| Таганай | 0.027 ± 0.006 | 7021.3 ± 1570 | 1.8 ± 0.4 | 1.4 ± 0.3 | 2.35 ± 0.52 |
| Машак | 0.032 ± 0.007 | 6894.1 ± 1541.6 | 1.2 ± 0.3 | 1.1 ± 0.25 | 1.78 ± 0.4 |
| Полярный Урал | 0.017 ± 0.004 | 6260.2 ± 1399.8 | 1.7 ± 0.4 | 1.02 ± 0.23 | 2.47 ± 0.55 |
| Туруханск | 0.03 ± 0.06 | 5912.6 ± 1322.1 | 3.25 ± 0.72 | 1.39 ± 0.31 | 2.48 ± 0.55 |
| Томск | 0.012 ± 0.002 | 5215 ± 1166.1 | 3.45 ± 0.77 | 1.7 ± 0.38 | 1.69 ± 0.38 |
| Ергаки | 0.03 ± 0.007 | 6605.7 ± 1477 | 2.7 ± 0.6 | 1.24 ± 0.27 | 2.1 ± 0.47 |
| Г. Шор. | 0.033 ± 0.008 | 6974.2 ± 1559.5 | 0.2 ± 0.04 | 1.2 ± 0.26 | 2.21 ± 0.5 |
| Тянь-Шань | 0.024 ± 0.005 | 7361.5 ± 1646.1 | 2.5 ± 0.6 | 1.1 ± 0.25 | 1.69 ± 0.38 |
| Северобайк. | 0.021 ± 0.005 | 6643.6 ± 1503.5 | 2.5 ± 0.6 | 1.2 ± 0.3 | 2.01 ± 0.45 |
| Газимур | 0.05 ± 0.01 | 8890.5 ± 1988 | 0.4 ± 0.09 | 1.2 ± 0.27 | 1.4 ± 0.31 |
| Лазо | 0.026 ± 0.006 | 7063.1 ± 1579 | 0.9 ± 0.2 | 1.01 ± 0.22 | 1.3 ± 0.29 |
| С. Алинь | 0.026 ± 0.006 | 6969.2 ± 1558 | 0.9 ± 0.2 | 1.005 ± 0.22 | 1.37 ± 0.3 |
| Якутия | 0.022 ± 0.004 | 4329.7 ± 893.2 | 1.37 ± 0.27 | 1.04 ± 0.21 | 1.65 ± 0.33 |
| Колыма | 0.041 ± 0.01 | 7269.2 ± 1625.5 | 1.8 ± 0.4 | 1 ± 0.22 | 1.46 ± 0.33 |
| Оймякон | 0.047 ± 0.01 | 6785.1 ± 1517.2 | 2.1 ± 0.5 | 1 ± 0.2 | 1.75 ± 0.39 |
| Магадан | 0.029 ± 0.01 | 5440.1 ± 1216.5 | 0.87 ± 0.19 | 1 ± 0.22 | 1.42 ± 0.32 |
| Камчатка | 0.043 ± 0.06 | 8983.6 ± 1270 | 0.95 ± 0.13 | 1 ± 0.14 | 1.94 ± 0.27 |
| Аляска | 0.023 ± 0.004 | 6814.6 ± 1265.4 | 1.3 ± 0.2 | 1 ± 0.18 | 1.41 ± 0.26 |
| Ногайск | 0.007 ± 0.001 | 11055.4 ± 2211 | 7.13 ± 1.42 | 1.82 ± 0.36 | 2.28 ± 0.46 |
| Сочи | 0.02 ± 0.004 | 9758.5 ± 1951.7 | 5.19 ± 1.03 | 1.14 ± 0.22 | 2.48 ± 0.5 |
| Архыз | 0.02 ± 0.003 | 12033.8 ± 2406.8 | 7.45 ± 1.49 | 1.96 ± 0.39 | 2.84 ± 0.56 |
| Нальчик | 0.022 ± 0.004 | 13560 ± 3019.4 | 15 ± 3.3 | 2 ± 0.45 | 2.41 ± 0.54 |
| Гуниб | 0.015 ± 0.003 | 17727.7 ± 3545.5 | 17.7 ± 3.5 | 2 ± 0.4 | 2.51 ± 0.5 |
| Талги | 0.015 ± 0.003 | 13202.1 ± 2640.4 | 13.8 ± 2.7 | 1.76 ± 0.35 | 2.51 ± 0.5 |
Остальные популяции разделены нами по географическому признаку на три группы: европейско-сибирскую, дальневосточную и североамериканскую. В первой группе предположительно могут быть популяции J. сommunis L. var. communis и J. communis var. saxatilis, во второй – J. communis var. saxatilis, в третьей – J. communis var. depressa.
Анализ методом главных компонент (рис. 2, 3) четко отделил кавказские популяции можжевельника как в плоскости первой и второй компонент, так и в плоскости первой и третьей компонент. Кавказские растения отличаются широкой и длинной хвоей, развитым проводящим пучком с большим количеством обкладочных клеток, крупным поперечным срезом, длинным кончиком хвои, широкой устьичной полоской с двумя полосками и Ка > 2 (коэффициент Адамса [7]). Переходной оказалась популяция из Сочи, которая по ряду параметров близка к альпийским растениям.
Рис. 2.
Ординация популяций J. communis в плоскости первых двух главных компонент ( – J. сommunis var. saxatilis (Дальний Восток),
– J. сommunis var. saxatilis (Дальний Восток),  – J. сommunis var. saxatilis (Европа, Сибирь);
– J. сommunis var. saxatilis (Европа, Сибирь);  – J. communis var. communis;
– J. communis var. communis;  – J. communis var. oblonga).
– J. communis var. oblonga).
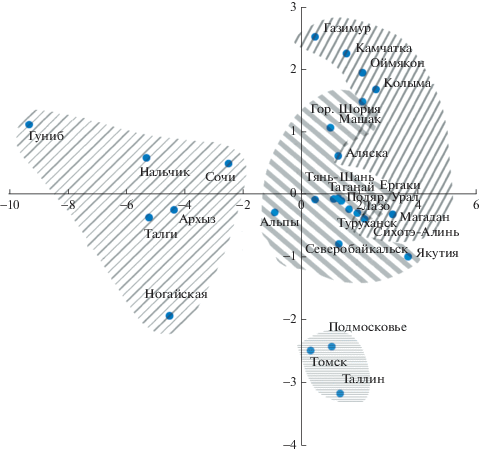
Рис. 3.
Ординация популяций можжевельника обыкновенного в плоскости первой и третьей главных
компонент ( – J. сommunis var. saxatilis (Дальний Восток),
– J. сommunis var. saxatilis (Дальний Восток),  – J. сommunis var. saxatilis (Европа, Сибирь);
– J. сommunis var. saxatilis (Европа, Сибирь);  – J. communis var. communis;
– J. communis var. communis;  – J. communis var. oblonga).
– J. communis var. oblonga).
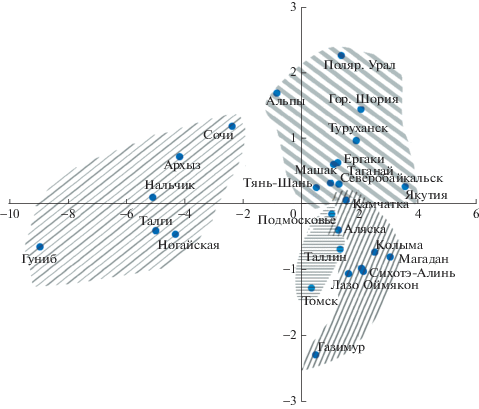
Вторую отдельную группу составили популяции J. сommunis var. communis Томска, Подмосковья и Таллина. В то же время в плоскости первой и третьей компонент эти популяции объединились с другими европейско-сибирскими можжевельниками. Эти популяции, помимо преобладающей прямостоячей формы растений, отличаются длинной, но узкой хвоей, мелкими смоляными ходами и проводящими пучками.
Остальные европейско-сибирские, дальневосточные и североамериканская популяции образовали единое облако. При этом дальневосточные популяции характеризуются более длинной, остроконечной, но узкой хвоей с Ка < 1.5, в то время как европейско-сибирские – более короткой, но широкой с Ка > 1.5. Можжевельник из Аляски по параметрам хвои не отличается от растений дальневосточных популяций.
Аллозимный анализ и анализ изменчивости хлоропластной ДНК
Закономерности, полученные при помощи анатомо-морфологического метода, нашли свое объяснение и стали более понятными, когда мы сгруппировали популяции по их генетическим дистанциям с помощью аллозимного метода и на основании парных Fst по данным изменчивости хлДНК.
Оба метода подтвердили результаты морфолого-анатомического анализа об отсутствии значительных различий между J. c. var. saxatilis и J. с. var. communis и показали слабую подразделенность изученных популяций на основной части ареала (Европа, Сибирь). Это косвенно указывает на значительный поток генов между этими популяциями и их недавнее общее происхождение.
В то же время по обоим молекулярным маркерам выявлена генетическая оригинальность североамериканской и всех дальневосточных популяций. Причем по ядерным маркерам эти две группы оказались близки, что совпадает с результатами морфолого-анатомического анализа (рис. 4). Однако по хлоропластным маркерам гаплотипы этих разновидностей образуют разные ветви на филогенетическом дереве [5], а популяции не объединяются в одну группу (рис. 5). Это несоответствие очевидно объясняется разным типом наследования данных маркеров. Ядерные маркеры наследуются у можжевельника и по материнской, и по отцовской линии, а хлоропластные – только по отцовской. Поток генов с пыльцой вероятно был ограничен гораздо раньше (со времени исчезновения Берингового моста суши), чем с семенами, которые могли распространять перелетные птицы. В целом, это может свидетельствовать об относительно недавнем расхождении этих популяций.
Рис. 4.
Ординация популяций J. сommunis, построенная на основании парных Fst по данным изменчивости хлДНК.
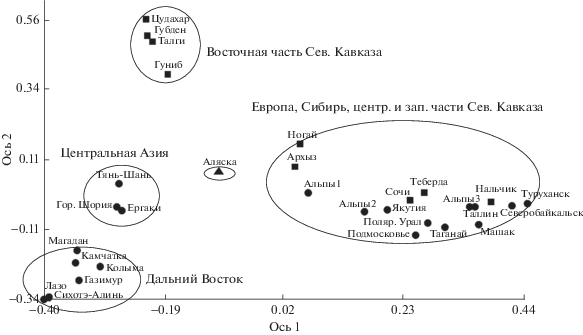
Рис. 5.
Ординация популяций J. communis, построенная на основании генетических дистанций по аллозимным локусам.
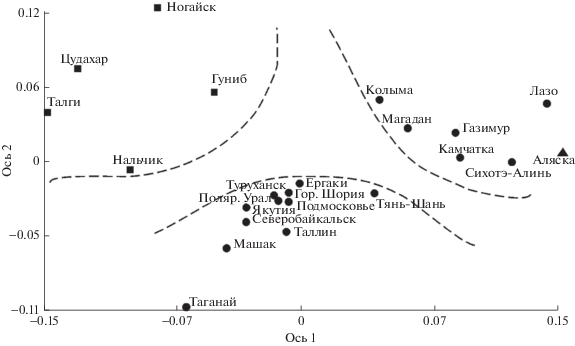
Аллозимный анализ однозначно подтверждает морфологические данные об обособленности J. communis var. оblonga (J. oblonga Bieb.) на Северном Кавказе. В данном регионе хлДНК и данные морфологии указывают на значительную неоднородность этого таксона.
Так на графике ординации по хлоропластным маркерам (рис. 5) видно, что популяции центральной части Северного Кавказа (Нальчик) и западной (Сочи, Архыз), а также полупустынная популяция из Ногайской степи находятся в общей группе со всеми евросибирскими популяциями, в то время как высокогорные популяции восточной части Северного Кавказа (Гуниб, Талги) образуют отдельную группу.
ОБСУЖДЕНИЕ
Исследование анатомо-морфологической и генетической изменчивости позволяет выделить несколько в разной степени обособленных групп. У можжевельника, произрастающего в восточной части Северного Кавказа все три метода подтверждают особый статус J. communis var. оblonga, а в западной и центральной частях Северного Кавказа – только морфологический и изоферментный методы. Тем не менее можно утверждать, что все кавказские популяции, имея крупную, остроконечную хвою с двумя полосками и Ка > 2, скорее всего являются отдельным таксоном (возможно следует поддержать его таксономический статус как самостоятельного вида J. oblonga Bieb. или подвида).
Близость гаплотипов можжевельника из западной и центральной частей Северного Кавказа к евросибирским гаплотипам может свидетельствовать о существовании в прошлом потока генов в этой более гумидной части Северного Кавказа в сторону других европейских горных систем, например Альп. В нашем исследовании в популяции из окрестностей Сочи, которая является крайней западной точкой на Кавказе, обнаружен редкий хлоропластный гаплотип, такой же, как в популяции из австрийских Альп. На многих графиках ординации по морфологическим параметрам популяция из Сочи занимает промежуточное положение между кавказской и евросибирской группами. А по такому показателю, как число полосок, эта популяция вообще не вошла в кавказскую группу.
Популяции восточной части Северного Кавказа, особенно выборка можжевельника, собранная на Гунибском плато, – это древний рефугиум, который оказался изолирован после каких-то климатических или топографических изменений, не вносил вклад в реколонизацию Европы и не имел вторичных контактов с ее популяциями. В Гунибской популяции при аллозимном анализе обнаружены редкие аллели, один из которых (в локусе SodA) был найден также только в популяциях Сихотэ-Алиня и южного Сахалина. По-видимому когда-то эти популяции были связаны непрерывным ареалом вида, сформировавшимся в плиоцене, который был разорван в результате климатических изменений в плейстоцене, в результате чего можжевельник оказался изолирован в рефугиумах Дальнего Востока и Восточного Кавказа. Это предположение подтверждается тем, что в результате анализа изменчивости хлоропластной ДНК группы гаплотипов можжевельника из популяций Дальнего Востока и Восточного Кавказа оказались принадлежащими к одной генетической линии, более древней по отношению к другим линиям [5]. Биогеографическая значимость Кавказского региона и деление его на юге на две части – западную и восточную, отмечены и в исследовании разных видов дубов [29]. Считают, что этот регион, благодаря своей геологической истории, климатическим особенностям и топографии, вероятно с конца миоцена играл важную роль в сохранении и возникновении биологического разнообразия видов.
Генетическая неоднородность кавказских можжевельников, сложность и комплексность их филогеографической истории и, возможно, явления гибридизации отразились и на их морфологических признаках. С этим же по-видимому связаны и разночтения в оценке таксономического статуса кавказских можжевельников у разных исследователей. Однако по главному морфологическому признаку – числу полосок хвоинки, все кавказские популяции (кроме Сочи) хорошо отличаются от других популяций J. communis. Кроме того они отличаются по ядерным маркерам. Принято считать, что изменчивость ядерных маркеров больше отражает таксономическую подразделенность видов, а изменчивость хлоропластная – географическую.
В этом отношении интересна картина, полученная для популяций можжевельника из Центральной Азии (Тянь-Шань, Горная Шория, Ергаки). Эта группа значимо отделилась при анализе хлоропластной ДНК, однако ни морфологический, ни аллозимный методы не показали дифференциации. Возможно такая картина отражает географическую изоляцию этой группы, но не таксономическую.
Методы генетической изменчивости дифференцируют североамериканскую разновидность J. communis var. depressa, произрастающую на Аляске. В то же время по параметрам хвои она не отделилась от дальневосточных популяций. Дальневосточный можжевельник, растущий на востоке за Верхоянским хребтом, имеет ряд особенностей, делающих его ближе к североамериканским растениям, чем сибирским. Этот можжевельник с длинной, но узкой хвоей с одной полосой и Ка (как правило) < 1.5 вероятно можно считать типичным J. communis var. saxatilis, к которому морфологически близка североамериканская популяция. Аллозимный метод также показывает бóльшую близость дальневосточных популяций к североамериканской J. communis var. depressa, чем к европейско-сибирским популяциям.
Генетические методы не выявили различий между J. communis var. communis и J. communis var. saxatilis, произрастающими в Европе и Азии (до Верхоянского хребта). Данные методы также показали слабую подразделенность изученных популяций на этой части ареала. Исследование же морфологии и анатомии хвои показывает высокий уровень изменчивости и неоднородности можжевельника на этой территории. Популяции из Томска, Подмосковья и Таллина характеризуются типичными для J. communis var. communis признаками (жизненная форма в виде деревца или конусовидного кустарника, длинная остроконечная хвоя), в то время как остальные популяции в разной степени проявляют особенности одной или другой вариации и, возможно, являются переходными формами между двумя вариациями. Тем не менее все можжевельники в этой части ареала являются стелющимися или раскидистыми кустарниками, они вероятнее всего относятся к J. communis var. saxatilis.
Отсутствие четких генетических границ, возможно, указывает на их общее происхождение и значительный поток генов между этими популяциями в последний ледниковый период и в межледниковье в результате существования северных микрорефугиумов. Различия же в морфологии и анатомии скорее всего объясняются экологическими и климатическими особенностями мест произрастания. Растения из Томска, Подмосковья и Таллина растут под пологом леса в условиях затенения, повышенной влажности, в то время как остальные популяции произрастают либо в горах, либо в суровых северных условиях.
В целом разные методы исследования биологической изменчивости в применении к можжевельнику обыкновенному подтверждают и уточняют друг друга. Некоторая противоречивость полученных результатов может быть отражением общих закономерностей, касающихся несовпадения скоростей морфологической, молекулярной и хромосомной эволюций.
Таким образом, проведенное комплексное исследование биологического разнообразия и генетической структуры можжевельника обыкновенного на основном ареале показывает высокий уровень изменчивости как морфолого-анатомических, так и генетических признаков, что приводит к большим трудностям при изучении внутривидовой таксономии вида. Не было найдено явных корреляций между фенотипом, генетической изменчивостью и таксономической принадлежностью изученных популяций (за исключением кавказского можжевельника).
Проведенное исследование подтверждает особый таксономический статус J. communis s. l., произрастающего на Кавказе, вероятно подвидового уровня или даже, самостоятельного вида (J. oblonga Bieb.). Реликтовые популяции кавказского можжевельника, особенно в восточной части Северного Кавказа, заслуживают дальнейшего изучения.
Североамериканский J. communis var. depressa и дальневосточный J. communis var. saxatilis не отличаются морфологически и анатомически, но различаются генетически и вероятно являются родственными генетическими линиями, которые произрастают в сходных климатических условиях и приобрели конвергентное внешнее сходство. Отличить их можно только генетически.
J. communis var. communis и J. communis var. saxatilis, произрастающие в Европе и Азии, не дифференцируются генетически, но имеют статистически значимые отличия по ряду морфологических и анатомических параметров хвои и преобладающей жизненной форме. Это позволяет заключить, что J. communis var. communis и J. communis var. saxatilis являются экотипами, которые произрастают на обширном ареале, в разных экологических условиях (под пологом или в горах) и в результате высокой экологической пластичности вида приобретают отличия в морфологии и анатомии. Другими словами, обладая высокой полиморфностью, можжевельник обыкновенный существует одновременно в виде нескольких жизненных форм. Под действием отбора в разных экологических условиях начинает преобладать одна из них, в то же время постоянно существующий обмен генами между популяциями поддерживает генетическое единообразие на обширном ареале.
За помощь в сборе материала на Кавказе мы выражаем благодарность д.б.н. З.М. Асадулаеву и А.М. Мусаеву (Горный ботанический сад Дагестанского научного центра РАН).
Исследования выполнены по базовому проекту № АААА-А17-117101820003-0 и при поддержке гранта РФФИ № 18-04-00616.
Настоящая статья не содержит каких-либо исследований с использованием в качестве объекта животных.
Настоящая статья не содержит каких-либо исследований с участием в качестве объекта людей.
Авторы заявляют, что у них нет конфликта интересов.
Список литературы
Taberlet P., Fumagalli L., Wust-Saucy A.-G., Cosson J.-F. Comparative phylogeography and postglacial colonization routes in Europe // Mol. Ecol. 1998. V. 7. P. 453–464.
Hewitt G.M. The genetic legacy of the Quaternary ice ages // Nature. 2000. V. 405. P. 907–913.
Petit R., Aguinagalde I., de Beaulieu J.-L. et al. Glacial refugia: hotspots but not melting pots of genetic diversity // Science. 2003. V. 300. P. 1563–1565.
Myking T. Evaluating genetic resources of forest trees by means of life history traits – a Norwegian example // Biodiversity and Conservation. 2002. V. 11. P. 1681–1696.
Hantemirova E.V., Heinze B., Knyazeva S.G. et al. A new Eurasian phylogeographical Paradigm? Limited contribution of southern populations to the recolonization of high latitude populations in Juniperus communis L. (Cupressaceae) // J. Biogeogr. 2017. V. 44. P. 271–282.
Farjon A. World Checklist and Bibliography of Conifers. England: The Royal Botanic Gardens, 2001. 309 p.
Adams R.P. Junipers of the World: The Genus Juniperus. 4th edn. Trafford Publishing, 2014. 422 p.
Ареалы деревьев и кустарников СССР. Ленинград: Наука, 1977. Т. 1. 164 с.
Деревья и кустарники СССР. М.: Изд-во Акад. наук СССР, 1949. Т. 1. 461 с.
Комаров В.Л. Сем. XIII. Cupressaceae F.W. Neger. Флора СССР. Т. 1. М.–Л.: Изд-во Академии наук СССР, 1934. С. 179–191.
Коропачинский И.Ю., Встовская Т.Н. Древесные растения азиатской России. Новосибирск, 2002. 402 с.
Князева С.Г. Опыт применения многомерных методов для определения таксономического положения Juniperus sibirica Burgsd. (Cupressaceae) // Ботан. журн. 2004. № 2. С. 236–244.
Князева С.Г. Изменчивость морфологических признаков сибирских видов можжевельника // Лесоведение. 2007. № 1. С. 65–69.
Князева С.Г. Внутривидовая изменчивость можжевельника обыкновенного на территории Сибири и Дальнего Востока // Лесоведение. 2010. № 4. С. 36–44.
Фарукшина Г.Г., Путенихин В.П. Можжевельники обыкновенный и казацкий на Южном Урале: распространение, популяционная структура, сохранение генофонда. Уфа: Гилем, Башк. энцикл., 2016. 168 с.
Галушко А.И. Флора Северного Кавказа. Ростов-на-Дону: : Изд-во Рост. ун-та, 1978. 318 с.
Джанаева В.М. Определитель семейства можжевеловых. Фрунзе: Изд-во Илим, 1969. 93 с.
Черепанов С.К. Сосудистые растения России и сопредельных государств. С-Пб: Мир и семья, 1995. 990 с.
Имханицкая Н.Н. Критическая заметка о кавказских видах секции Juniperus рода Juniperus L. // Новости сист. высш. раст. 1990. № 27. С. 5–16.
Хантемирова Е.В., Беркутенко А.Н., Семериков В.Л. Систематика и геногеография Juniperus communis L. по данным изоферментного анализа // Генетика. 2012. Т. 48. № 9. С. 1–8.
Harris H., Hopkinson D.A. Handbook of Enzyme Electrophoresis. Amsterdam: North_Holland Publ. Comp., 1976.
Подогас А.В., Шурхал А.В., Семериков В.Л., Ракицская Т.А. Генетическая изменчивость ферментов хвои сосны кедровой сибирской (Pinus sibirica Du Tour) // Генетика. 1991. Т. 27. № 4. С. 695–703.
Хантемирова Е.В., Семериков В.Л. Аллозимный полиморфизм разновидностей можжевельника обыкновенного // Лесоведение. 2009. № 1. С. 74–77.
Nei M. Estimation of average heterozygosity and genetic distance from a small number of individuals // Genetics. 1978. V. 83. P. 583–590.
Swofford D.L., Selander R.B. BIOSYS_1: a FORTRAN program for the comprehensive analysis of electrophoretic data in population genetics and systematics // J. Heredity. 1981. V. 72. P. 281–283.
Rohlf F.J. NTSYS_pc: Numerical Taxonomy and Multivariate Analysis System. Ver. 2.02. [Computer progr.]. N.Y.: Exeter Publ. Ltd., 1988.
Devey M.E., Bell J.S., Smith D.N. et al. A genetic linkage map for Pinus radiata based on RFLP, RAPD and microsatellite markers // Theor. Applied Genet. 1996. V. 92. P. 673–679.
Taberlet P.T., Geilly L., Patou G., Bouvet J. Universal primers for amplification of three non-coding regions of chloroplast DNA // Plant Mol. Biol. 1991. V. 17. P. 1105–1109.
Ekhvaia J., Simeone M.K., Silakadze N., Abdaladze O. Morphological diversity and phylogeography of the Georgian durmast oak (Q. petraea subsp. iberica) and related Caucasian oak species in Georgia (South Caucasus) // Tree Genetics & Genomes. 2018. V. 14. P. 17.
Дополнительные материалы отсутствуют.