

Генетика, 2021, T. 57, № 1, стр. 116-122
Результаты длительного мониторинга и эксперимента по обогащению генофонда популяций кустарниковой улитки Bradybaena fruticum (Müll.) на урбанизированных территориях
В. М. Макеева 1, *, И. Д. Алазнели 1, А. В. Смуров 1, Д. В. Политов 2, Ю. С. Белоконь 2, М. М. Белоконь 2
1 Московский государственный университет им. М.В. Ломоносова
119991 Москва, Россия
2 Институт общей генетики им. Н.И. Вавилова Российской академии наук
119991 Москва, Россия
* E-mail: vmmakeeva@yandex.ru
Поступила в редакцию 19.02.2020
После доработки 18.06.2020
Принята к публикации 08.07.2020
Аннотация
Результаты длительного мониторинга динамики фенетических признаков в десяти популяциях кустарниковой улитки из Подмосковья и парков г. Москвы зафиксировали постоянство частоты аллеля полосатости раковин в крупных природных популяциях и изменение частот в мелких городских изолятах. Одновременно в череде поколений выявлена стойкая тенденция увеличения частоты желтых раковин, что объясняется происходящим осветлением местообитаний и служит подтверждением адаптивности признака цвета раковин. Представлены результаты длительного эксперимента по поддержанию жизнеспособности четырех городских популяций кустарниковой улитки Bradybaena fruticum (Müll.), начатого с обогащения их генофонда в 2003 г. и проведения контроля в 2005 г., выявившего изменение частоты аллеля полосатости раковины. Повторный контроль состояния генофонда оздоровленных популяций (2017–2019 гг.) выявил сохранение частоты этого признака с 2005 г. на протяжении двух поколений моллюсков, в том числе в двух популяциях при p < < 0.01, одной p < 0.05. Таким образом, контроль результатов эксперимента по обогащению популяций подтверждает, что способ поддержания жизнеспособности популяций на урбанизированных территориях (защищенный патентом на изобретение) эффективен для восстановления генетического разнообразия и жизнеспособности популяций животных.
Сохранение биоразнообразия является одним из необходимых условий устойчивого развития человечества [1]. Сохранение биоразнообразия связано не только с сохранением местообитаний и поддержанием численности вида, но в значительной степени – с поддержанием генетического разнообразия популяций.
Устойчивость популяций животных и растений обеспечивается благодаря гомеостазу, т.е. постоянству соотношения частот аллелей генов. Использование частот аллелей генов позволяет оценить генотипическую структуру популяций. В условиях урбанизированных ландшафтов фрагментация как правило ведет к изоляции мелких популяций, сокращению их генетического разнообразия (вследствие активизации дрейфа генов и инбридинга) и вымиранию [2, 3]. Поэтому для обеспечения жизнеспособности популяций на урбанизированных территориях необходим контроль состояния генофонда и поддержание его разнообразия [4].
Это является главной методологической основой геноурбанологии – нового научного направления, разработанного авторами [4–6]. В рамках геноурбанологии получен патент на изобретение № 2620079 “Способ поддержания жизнеспособности популяций животных или растений на урбанизированных территориях” [7], который позволяет остановить процессы устойчивого необратимого изменения генофондов, вызванные антропогенной изоляцией. Апробация способа была проведена на примере четырех популяций кустарниковой улитки, обитающих на особо охраняемых территориях города Москвы, в 2003–2005 гг. [6].
Необходимо отметить, что ранее сокращение генетического разнообразия было выявлено в популяциях четырех модельных видов животных и растений в условиях урбанизированного ландшафта города Москвы и Подмосковья, что несомненно свидетельствует об уменьшении адаптивного потенциала и жизнеспособности популяций [8–10].
Задача настоящего исследования состоит в анализе динамики генофонда популяций кустарниковой улитки по результатам длительного эколого-генетического мониторинга фенетических признаков, а также – в проведении контрольного обследования состояния генофонда экспериментально обогащенных в 2003 г. четырех городских популяций этого модельного объекта.
Кустарниковая улитка широко распространена в европейской части России, Западной Сибири – в пойменных лесах и на склонах сырых оврагов, в том числе и в антропогенных местообитаниях. Этот вид включен в список видов, предложенных для внесения в Красную Книгу города Москвы [11].
Кустарниковая улитка является удобным объектом для мониторинга динамики структуры популяций, т.к. обладает ярко выраженным генетически детерминированным полиморфным признаком – наличием или отсутствием полосы на раковине, с доминированием аллеля бесполосости [12]. Раковины также полиморфны по цвету. Выделяются красные, коричневые и желтые морфы. Цвет раковины не является четким дискретным признаком и детерминирован полигенно. Продолжительность жизни особей составляет 5–6 лет.
Эколого-генетический мониторинг популяций кустарниковой улитки начат в 1975 г. [13] и продолжается по настоящее время. Описания местообитаний исследованных за этот период 20-ти популяций улиток в Москве и Подмосковье, а также методика сбора материала, результаты оценки состояния генофонда с использованием биохимических маркеров и фенетических признаков подробно изложены ранее [8, 14–17].
В настоящем сообщении приведены результаты мониторинга десяти популяций, из которых шесть обитают на особо охраняемых территориях (ООПТ) г. Москвы, четыре – в дальнем и ближнем Подмосковье. Мониторинг двух популяций проводится с 1975 г., одной – с 1989 г., одной – с 1998 г., остальных – с 2003 г. Последнее контрольное обследование состояния генофонда популяций проведено в 2017–2019 гг., объем материала составляет 1310 экземпляров. Для изучения динамики частот фенетических признаков во времени использованы также данные исследований популяций улиток, проведенных ранее [13–17] в количестве более 12 000 раковин.
Пункты сбора улиток в Москве: Национальный парк Лосиный остров; особо охраняемые территории (ООПТ) г. Москвы: природно-исторические парки: Измайлово, Кузьминки–Люблино; парк Битцевский лес; природный заказник Воробьевы горы, усадьба Узкое, Главный Ботанический сад. Улиток собирали в трех пунктах на западе Московской обл., в Звенигородском р-не: природный заказник “Звенигородская биостанция МГУ и карьер Сима”, дер. Городок на берегу Можайского водохранилища, дер. Мешково (Киевское шоссе).
Анализ состояния генофонда десяти популяций кустарниковой улитки в пространстве и во времени проводился с использованием двух фенетических признаков: наличия или отсутствия полосы на раковине и наличия ярко желтой окраски раковины.
Для сравнения выборок из исследованных популяций использовалась частота аллеля полосатости, которая рассчитывалась по доле полосатых особей, проявляющих рецессивный ген [18]. При этом доминантная гомозигота и гетерозигота имеют одинаковое фенотипическое выражение – отсутствие полосы на раковине. В расчетах частот аллелей исходили из предположения, что соотношение генотипов в популяциях соответствует равновесию Харди–Вайнберга, поскольку данные, полученные нами ранее, указывают на равновесное распределение генотипов аллозимных локусов у данного вида [8].
Для проведения сравнительного графического анализа популяций в пространстве и во времени рассчитывались доверительные границы для доли, среднее квадратическое отклонение (σp), средняя ошибка для доли (Sp) [18].
Выборки сравнивались с помощью критерия Фишера с предварительным преобразованием долей в радианы (метод φ) [19].
Анализ частот раковинных фенотипов выявил стойкое сохранение частоты аллеля полосатости – генетически детерминированного признака наличия или отсутствия полосы на раковине в популяциях, разделенных антропогенными барьерами (рис. 1, табл. 1). Постоянство частот сохраняется не только в ограниченный период времени (3–5 лет), но и на протяжении 3–7 поколений (одно поколение шесть лет). В целом длительный мониторинг выявил колебания частот признаков вокруг средней величины, что подтверждает адаптивный характер полиморфизма (выявленный ранее), дающий возможность популяции выжить в колеблющихся условиях среды [16, 17] (табл. 1). Необходимо отметить, что постоянство генетической структуры выявлено главным образом для крупных природных популяций (Звенигород, Городок, Мешково). В то же время в мелких изолированных городских популяциях, испытывающих максимальное антропогенное воздействие, отмечены достоверные изменения частот аллелей (Воробьевы горы, Кузьминки, Лосиный остров) (рис. 1). Полученные данные о постоянстве частот полосатых морф в природных популяциях кустарниковой улитки согласуются с ранними работами авторов [17] и с современными работами других исследователей, выявившими стойкое сохранение частот полосатых морф на протяжении шестнадцати лет в популяции наземного моллюска Helix albescens, Rossmassler в Крыму [20] и достоверные изменения частот фенетических признаков в популяциях наземных моллюсков Bradybaena fruticum [21] и Сepaea vindobonensis, Ferussuc [22], обитающих на урбанизированных территориях.
Рис. 1.
Частоты аллеля полосатости раковин в десяти популяциях кустарниковой улитки в парках г. Москвы и в Подмосковье по результатам длительного мониторинга.
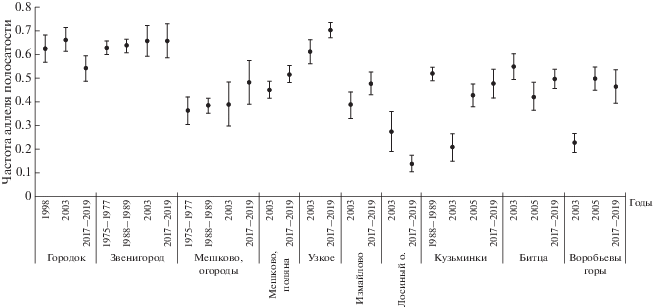
Таблица 1.
Частоты аллеля полосатости, полосатых и желтых раковин в десяти популяциях кустарниковой улитки парков Москвы и в Подмосковье по результатам длительного мониторинга
| Пункт сбора | Год | Численность | Частота полосатых раковин | Частота аллеля полосатости | Частота желтых раковин | σp | Sp | ||
|---|---|---|---|---|---|---|---|---|---|
| всего | полосатых | желтых | |||||||
| Городок | 1998 | 134 | 52 | 32 | 0.3881 | 0.6229 | 0.2388 | 0.485 | 0.030 |
| 2003 | 171 | 75 | 47 | 0.4386 | 0.6623 | 0.2749 | 0.473 | 0.026 | |
| 2017–2019 | 168 | 49 | 55 | 0.2917 | 0.5401 | 0.3274 | 0.498 | 0.027 | |
| Звенигород | 1975–1977 | 4250 | 1672 | 349 | 0.3934 | 0.6272 | 0.0821 | 0.484 | 0.005 |
| 1989 | 567 | 229 | 42 | 0.4789 | 0.6920 | 0.0741 | 0.462 | 0.015 | |
| 2003 | 102 | 44 | 16 | 0.4314 | 0.6568 | 0.1569 | 0.475 | 0.033 | |
| 2017–2019 | 86 | 37 | 28 | 0.4302 | 0.6559 | 0.3256 | 0.475 | 0.036 | |
| Мешково, огороды | 1975–1977 | 3185 | 417 | 343 | 0.1309 | 0.3618 | 0.1077 | 0.481 | 0.029 |
| 1988 | 451 | 66 | 100 | 0.1118 | 0.3344 | 0.2217 | 0.486 | 0.016 | |
| 2003 | 53 | 8 | – | 0.1509 | 0.3885 | 0.0000 | 0.487 | 0.047 | |
| 2017–2019 | 56 | 13 | 6 | 0.2321 | 0.4818 | 0.1071 | 0.500 | 0.047 | |
| Мешково, поляна у Киевского шоссе | 2003 | 367 | 74 | 22 | 0.2016 | 0.4490 | 0.0599 | 0.497 | 0.018 |
| 2017–2019 | 188 | 50 | 27 | 0.2660 | 0.5157 | 0.1436 | 0.500 | 0.026 | |
| Узкое | 2003 | 436 | 163 | 96 | 0.3739 | 0.6114 | 0.2202 | 0.487 | 0.017 |
| 2017–2019 | 128 | 63 | 3 | 0.4922 | 0.7016 | 0.0234 | 0.458 | 0.029 | |
| Измайловский парк | 2003 | 195 | 29 | 12 | 0.1487 | 0.3856 | 0.0615 | 0.487 | 0.025 |
| 2017–2019 | 66 | 15 | 21 | 0.2273 | 0.4767 | 0.3182 | 0.499 | 0.043 | |
| Лосиный остров | 2003 | 320 | 24 | 18 | 0.0750 | 0.2739 | 0.0563 | 0.446 | 0.018 |
| 2017–2019 | 104 | 2 | 39 | 0.0192 | 0.1387 | 0.3750 | 0.346 | 0.024 | |
| Кузьминки | 1988–1989 | 561 | 150 | 27 | 0.3825 | 0.5171 | 0.0481 | 0.500 | 0.015 |
| 2003 | 94 | 4 | 5 | 0.0426 | 0.2063 | 0.0532 | 0.405 | 0.030 | |
| 2005 | 204 | 37 | 34 | 0.1814 | 0.4259 | 0.1667 | 0.494 | 0.024 | |
| 2017–2019 | 128 | 29 | 24 | 0.2266 | 0.4760 | 0.1875 | 0.499 | 0.031 | |
| Битцевский лес | 2003 | 160 | 48 | 9 | 0.3000 | 0.5477 | 0.0563 | 0.498 | 0.028 |
| 2005 | 129 | 23 | 16 | 0.1783 | 0.4222 | 0.1240 | 0.494 | 0.031 | |
| 2017–2019 | 289 | 71 | 39 | 0.2457 | 0.4957 | 0.1349 | 0.500 | 0.021 | |
| Воробьевы горы | 2003 | 216 | 11 | 30 | 0.0509 | 0.2257 | 0.1389 | 0.418 | 0.020 |
| 2005 | 202 | 50 | 32 | 0.2475 | 0.4975 | 0.1584 | 0.500 | 0.025 | |
| 2017–2019 | 97 | 21 | 16 | 0.2165 | 0.4653 | 0.1649 | 0.499 | 0.036 | |
В целом результаты анализа популяций моллюсков (с использованием аллеля полосатости), разграниченных естественными и созданными человеком барьерами, позволяют сделать заключение о решающей роли антропогенной инсуляризации в формировании пространственно-популяционной структуры вида. Сильное влияние антропогенной инсуляризации за период наблюдения (43 года) сказалось также в резком сокращении площади обитания и численности особей в изолятах (в 25–50 раз), в сокращении генетического разнообразия популяций. За период длительного мониторинга исчезло семь популяций моллюсков (33.3% от общего числа популяций) и 19 участков, различающихся фитоценотически (54.3% от общего числа участков) вследствие разрушения их местообитаний.
Сравнительный анализ популяций в пространстве и во времени с использованием признака частоты ярко-желтых раковин выявил стойкую тенденцию увеличения частоты желтых раковин (табл. 1), (различия достоверны в восьми из десяти популяций). Это может быть объяснено происходящим осветлением местообитаний, отмеченным нами ранее [15, 17], и служит подтверждением адаптивности признака цвета раковин. Необходимо отметить, что случаи отклонения от выявленной тенденции, например в популяции “Узкое” (табл. 1), связаны с сильным антропогенным воздействием, выражающемся в постоянном сенокошении в местах обитания улиток, приводящему к сокращению площади обитания и численности популяции.
Также было подтверждено выявленное нами ранее [15, 17] преобладание желтых бесполосых раковин в открытых фитоценозах по сравнению с затененными, что обеспечивает бóльшую защиту моллюсков при апостатическом (хищниками) отборе. Кроме того, тот или иной цвет раковины обеспечивает разную степень регуляции нагрева тела, следовательно, разную скорость потери воды. Эксперименты, проведенные нами ранее, подтвердили бóльшую выживаемость бесполосых моллюсков с желтым цветом фона раковин при нагревании, по сравнению с другими морфами [23].
В 2003 г. в четырех популяциях из парков Москвы (Кузьминки, Битцевский лес, Воробьевы горы, Ботанический сад), испытывающих максимальный антропогенный пресс (генофонд и эффективная численность были оценены как критические) [8], было проведено экспериментальное оздоровление генофонда [5, 6]. Решение об оздоровлении популяций было принято на основании результатов оценки состояния генофонда 20 популяций кустарниковой улитки из парков Москвы и Подмосковья (13 полиморфных изоферментных локусов, 2730 экземпляров) [8]. Для оздоровления генофонда использована разработанная технология сохранения жизнеспособности популяций животных на урбанизированных территориях, защищенная впоследствии патентом [7]. В городские популяции было внесено определенное расчетное число улиток из эталонных популяций Московской обл. (Звенигород, Городок, Мешково). В 2005 г. проведено первое контрольное определение состояния генофонда, которое выявило изменение частот аллеля полосатости у потомства во всех популяциях. Повторный контроль состояния генофонда оздоровленных популяций проведен по прошествии 12 лет – двух поколений (2017–2019 гг.). При этом одна из четырех обогащенных популяций (Ботанический сад) исчезла вследствие полного разрушения местообитания. Контроль выявил сохранение частот полосатых раковин и аллеля полосатости в обогащенных популяциях с 2005 г. (табл. 2). При этом в двух популяциях наблюдались достоверные отличия (по сравнению с 2003 г.) при p < 0.001, в одной – при p < 0.05. Необходимо отметить, что тенденция увеличения частоты ярко желтых раковин сохранилась и в обогащенных популяциях.
Таблица 2.
Степень различия популяций по частотам аллеля полосатости, по результатам длительного эксперимента по оздоровлению генофонда
| Пункт сбора | Год | Численность | Частота | d.f. | F-критерий | Вероятность различия по сравнению с 2005 г. | ||
|---|---|---|---|---|---|---|---|---|
| всего | полоса-тых | полоса-тых | аллеля | |||||
| Кузьминки | 2003 | 94 | 4 | 0.0426 | 0.2063 | 296 | 14.7698 | 0.999 |
| 2005 | 170 | 37 | 0.2176 | 0.4665 | – | – | – | |
| 2017–2019 | 128 | 29 | 0.2266 | 0.4760 | 330 | 0.7983 | Различия нет | |
| Битцевский лес | 2003 | 160 | 48 | 0.3000 | 0.5477 | 287 | 4.5255 | 0.95 |
| 2005 | 129 | 23 | 0.1783 | 0.4222 | – | – | – | |
| 2017–2019 | 289 | 71 | 0.2457 | 0.4957 | 416 | 1.9390 | Различия нет | |
| Воробьевы горы | 2003 | 216 | 11 | 0.0509 | 0.2257 | 416 | 34.6080 | 0.999 |
| 2005 | 202 | 50 | 0.2475 | 0.4975 | – | – | – | |
| 2017–2019 | 97 | 21 | 0.2165 | 0.4653 | 297 | 0.2727 | Различия нет | |
Таким образом, результаты эксперимента по обогащению генофонда популяций кустарниковой улитки подтвердили эффективность использования разработанного способа для восстановления их генетического разнообразия и жизнеспособности.
В заключение необходимо отметить, что фенетический признак наличия или отсутствия полосы на раковине с успехом был использован (для практических целей) для быстрой оценки состояния генофонда после проведенного оздоровления популяций на особо охраняемых территориях (ООПТ) г. Москвы [3, 5, 6]. Подобные фенетические признаки могут быть использованы и для других видов флоры или фауны при организации мероприятий по их сохранению на городских ООПТ. Однако несомненно, что более точную оценку дает использование кодоминантных генетических маркеров [8–10].
Длительный эксперимент по поддержанию жизнеспособности популяций был начат в рамках НИОКР Департамента природопользования и охраны окружающей среды г. Москвы в соответствии с целевой программой по восстановлению биоразнообразия Москвы по проекту “Восстановление генетического разнообразия исчезающих видов беспозвоночных животных на ООПТ города Москвы (на примере кустарниковой улитки), 2002–2003 гг.”.
Настоящее исследование (продолжение длительного эксперимента) проведено в рамках темы государственного задания, регистрационный № АААА-А16-116042010089-2 “Экология”. “Биосферные функции природных экосистем и рациональное природопользование”. Частично – в рамках темы государственного задания № 0112-2019-0001, а также по Программе обучения в аспирантуре, совместно реализуемой Московским государственным университетом им. М.В. Ломоносова и университетом МГУ–ППИ в Шеньчжэне, Китай.
Все применимые международные, национальные и/или институциональные принципы ухода и использования животных были соблюдены.
Авторы заявляют, что у них нет конфликта интересов.
Список литературы
Конвенция устойчивого развития мира // Программа действий: повестка дня на 21 век и другие документы конференции в Рио-де-Жанейро. Женева: Центр “За наше общее будущее”, 1993. 70 с.
Wright S. Coefficient of inbreeding and relationship // Amer. Natur. 1922. V. 56. P. 330–339.
Макеева В.М. Эколого-генетические основы охраны животных антропогенных экосистем (на примере Москвы и Подмосковья) // Автореф. дис. докт. биол. наук. М.: МГУ им. Ломоносова, 2008. 47 с.
Макеева В.М., Белоконь М.М., Смуров А.В. Геноурбанология как основа устойчивого сохранения биоразнообразия и экосистем в условиях глобальной урбанизации // Усп. соврем. биол. 2013. Т. 133. № 1. С. 19–34. https://doi.org/10.1134/S207908641304004
Makeeva V.M., Smurov A.V., Politov D.V. et al. Technology for restoring and maintaining sustainability of populations: Practical and theoretical results of genourbanology // The Open Conf. Proc. J. 2015. V. 6. P. 1–9. DOI 102174/221028920150601001
Макеева В.М., Смуров А.В. Эколого-генетическая диагностика состояния и методы восстановления популяций животных городских особо охраняемых природных территорий (на примере модельных видов в городе Москве) // Науч. ведом. Белгородского ун-та. Серия “Естеств. науки”. 2011. Вып. 14. № 3(98). С. 104–110.
Макеева В.М., Смуров А.В. Патент № 2620079. Способ поддержания жизнеспособности популяций животных или растений на урбанизированных территориях. 2017.
Макеева В.М., Белоконь М.М., Малюченко О.П. Оценка состояния генофонда природных популяций беспозвоночных животных в условиях фрагментированного ландшафта Москвы и Подмосковья (на примере кустарниковой улитки, Bradybaena fruticum (Müll.)) // Генетика. 2005. Т. 41. № 11. С. 1495–1510.
Макеева В.М., Белоконь М.М., Малюченко О.П., Леонтьева О.А. Оценка состояния генофонда природных популяций позвоночных животных в условиях фрагментированного ландшафта Москвы и Подмосковья (на примере бурых лягушек) // Генетика. 2006. Т. 42. № 5. С. 628–642.
Макеева В.М., Смуров А.В., Политов Д.В. и др. Сравнительная оценка состояния генофонда и жизнеспособности лесопосадок из парков города Москвы и естественных популяций из Подмосковья на примере ели европейской (Picea abies (L.) Karst.) // Генетика. 2018. Т. 54. № 9. С. 1015–1025. https://doi.org/10.1134/S0016675818090096
Красная книга города Москвы. М.: АБФ, 2001. 610 с.
Хохуткин И.М. О наследовании признака “опоясанности” в естественных популяциях наземного брюхоногого моллюска Bradybaena fruticum (Müll.) // Генетика. 1979. Т. 15. № 5. С. 868–871.
Макеева В.М. Популяционная структура кустарниковой улитки Bradybaena fruticum (Müll.) в условиях антропогенного ландшафта Подмосковья // Автореф. дис. … канд. биол. наук. М.: МГУ им. Ломоносова, 1980. 24 с.
Макеева В.М. Эколого-генетический анализ структуры колоний кустарниковой улитки Bradybaena fruticum (Müll.) в условиях антропогенного ландшафта Подмосковья // Журн. общей биол. 1988. Т. 49. № 3. С. 333–342.
Макеева В.М. Роль естественного отбора в формировании генетического своеобразия популяций моллюсков (на примере кустарниковой улитки Bradybaena fruticum (Müll.)) // Журн. общей биологии. 1989. Т. 30. № 1. С. 101–107.
Макеева В.М., Пахорукова Л.В., Уголкова Н.Г. Анализ динамики полиморфных признаков в популяциях кустарниковой улитки, Bradybaena fruticum (Müll.), в целях экологического мониторинга // Журн. общей биол. 1995. Т. 56. № 5. С. 570–587.
Макеева В.М., Пахорукова Л.В. Роль естественного отбора в поддержании устойчивости генотипического состава природных популяций кустарниковой улитки Bradybaena fruticum (Müll), Mollusca, Gastropoda по результатам экологического мониторинга // Журн. общей биологии. 2000. Т. 61. № 6. С. 638–661.
Рокицкий П.Я. Введение в статистическую генетику. Минск: Высш. школа, 1974. 447 с.
Зайцев Г.Н. Методика биометрических расчетов. М.: Наука, 1979. 255 с.
Крамаренко С.С., Крамаренко А.С. Пространственно-временная изменчивость фенетической структуры метапопуляции наземного моллюска Helix albescens Rossmassler, 1839 (Gastropoda: Pulmonata: Helicidae) // Науч. ведомости Белгородского ун-та. Серия “Естеств. науки”. 2009. № 11(66). С. 55–61.
Снегин Э.А. Временная динамика генетической структуры и эффективная численность популяций Bradybaena fruticum Müll. (Mollusca, Gastropoda, Pulmonata) в условиях Юга Среднерусской возвышенности // Экология. 2015. № 3. С. 198–205. https://doi.org/10/7868/S0367059715020110
Крамаренко С.С., Хохуткин И.М., Гребенников М.Е. Особенности фенетической структуры наземного моллюска Cepae vindobonensis (Pulmonata: Helicidae) в урбанизированных и природных популяциях // Экология. 2007. № 1. С. 42–48.
Макеева В.М., Матекин П.В. Физиологический эффект полиморфных признаков кустарниковой улитки Bradybaena fruticum (Müll.) // Журн. общей биол. 1994. Т. 55. № 3. С. 347–355.
Дополнительные материалы отсутствуют.