

Генетика, 2021, T. 57, № 12, стр. 1386-1395
Филогенетический анализ южносибирской гаплогруппы Q-YP1102 по данным о Y-SNP- и Y-STR-маркерах у тувинцев и окружающих популяций
А. Т. Агджоян 1, 2, *, Л. Д. Дамба 4, В. М. Гурьянов 1, В. В. Запорожченко 1, 2, О. П. Балановский 1, 2, 3
1 Институт общей генетики им. Н.И. Вавилова Российской академии наук
119991 Москва, Россия
2 Медико-генетический научный центр им. академика Н.П. Бочкова
115522 Москва, Россия
3 АНО “Биобанк Северной Евразии”
115201 Москва, Россия
4 Научно-исследовательский институт медико-социальных проблем
и управления Республики Тыва
667003 Кызыл, Россия
* E-mail: aagdzhoyan@gmail.com
Поступила в редакцию 31.05.2021
После доработки 18.06.2021
Принята к публикации 26.07.2021
Аннотация
Гаплогруппа Q Y-хромосомы составляет основу генофонда американских индейцев, а в Евразии наибольших частот достигает в популяциях Центральной и Южной Сибири. Проведенное генотипирование обширной панели SNP-маркеров, выделяющих около 60 ветвей в составе гаплогруппы Q, позволило провести углубленный филогеографический анализ в популяциях Южной Сибири и смежных регионов. Изучение шести популяций тувинцев и тоджинцев выявило три ветви гаплогруппы Q, восходящие к одному корню – YP1102. Наиболее частая из них линия YP1691 достигает максимума в генофонде северо-восточных тувинцев-тоджинцев (51%), понижаясь в частоте с востока на запад до 4%. Подобному тренду следует распространение и базальной линии YP1102*. Третья линия BZ99* редка у тувинцев (2%), но заметна в генофонде хакасов (9%). Филогенетический анализ STR-гаплотипов показал распространение одних и тех же кластеров не только у разных групп тувинцев, но и среди окружающих популяций. Структура основного кластера свидетельствует об увеличении численности носителей линии YP1691 в период позднего железного века. Карта частоты линии YP1691 отражает последствия активных миграционных процессов в течение нашей эры на территории Южной и Западной Сибири. Среди древних популяций Сибири вариант YP1102* найден у пяти индивидов: четырех представителей бронзового века Прибайкалья и одного представителя культуры скифо-сибирского типа на границе Монголии и Тувы. Данные древней ДНК и более высокое генетическое разнообразие в населении Южной Сибири позволяют предположить происхождение гаплогруппы Q-YP1102 в данном регионе и ее дальнейшее проникновение на запад и север Сибири, в том числе, возможно, с предками популяций енисейской лингвистической семьи.
Филогеографические исследования Y-хромосомы широко используются в популяционной генетике и в смежных дисциплинах (археологии, генетической генеалогии, криминалистике) для изучения миграции древнего населения, в том числе известных из небиологических источников (лингвистики, археологии, ранней истории), а также для анализа происхождения отдельных генеалогических линий. При изучении генофонда Сибири основное внимание уделяется двум гаплогруппам Y-хромосомы, которые отражают наследие древнего населения и связи между крупными частями Сибири: паттерн распространения гаплогруппы N связывает Южную и Западную Сибирь, а гаплогруппы Q – Южную и Центральную Сибирь. Особая роль населения Южной Сибири, и в первую очередь Алтае-Саянского нагорья, в формировании общей структуры населения, лингвистического и генетического разнообразия Сибири хорошо известна. В предыдущих работах мы изучили генофонд популяций Саян (тувинцев и тоджинцев) по панели основных гаплогрупп Y‑хромосомы [1–3], в том числе определены частоты гаплогруппы Q. Предметом данного исследования является углубленный анализ филогенетики и филогеографии гаплогруппы Q у тувинцев и других популяций Южной Сибири и смежных регионов на основе глубокого субтипирования нескольких десятков SNP-маркеров, определяющих отдельные линии внутри гаплогруппы Q.
Ввиду значительного возраста гаплогруппы Q‑M242 (около 30 тыс. лет) и разного паттерна распространения ее ветвей в Евразии и Америке (она охватывает практически весь генофонд американских индейцев и является маркером первичного заселения человеком Нового Света) сохраняется широкий интерес исследователей и к истории этой гаплогруппы на ее “прародине” – в Сибири. В связи с этим продолжаются работы и попытки изучения филогеографии гаплогруппы Q-M242 на современном уровне – с использованием данных NGS-анализа в современных и древних популяциях. Например, работа [4] является практически исчерпывающей по охвату ветвей гаплогруппы Q в популяциях Евразии и Америки, не фокусируясь на какой-либо из них. Основная задача исследования состояла в изучении доисторических миграций между континентами на основе подробного филогенетического дерева гаплогруппы Q. При этом вопросы распространения конкретных ветвей внутри отдельных регионов, включая Сибирь, практически не рассматривались.
В других недавних работах анализ гаплогруппы Q не был основным предметом исследования, а являлся частью более широкой задачи – изучения генофонда Северо-Восточной Азии [5] или в целом Восточной Азии [6] на основе полногеномных данных о современных и древних популяциях. В работе [5] основные выводы связаны с динамикой населения в Сибири в период от позднего палеолита до неолита на основе полногеномных панелей SNP-маркеров, но также включают и результаты анализа Y-хромосомы у некоторых древних индивидов (обнаружены только гаплогруппы Q или N). Исследование [6] охватывает территории Монголии (в том числе приграничье с Тувой), Северного Китая, Приамурья и Японии, включает результаты анализа образцов древней ДНК за последние 8 тыс. лет, среди которых встречена и гаплогруппа Q‑M242.
В более локальном географическом (территория Тувы) и хронологическом (последние 2.5 тыс. лет) масштабе изучены древние популяции скифского времени в работе [7]. Показано, что сообщества сибирских скифов были организованы как патрилокальные и патрилинейные (в мужских захоронениях обнаружены близкие родственники), по структуре генофонда эти группы оказались более близки к населению Азии, чем к западным группам скифов. В данной работе у пяти индивидов захоронения Аржан (раннескифского времени) обнаружена гаплогруппа Y-хромосомы Q-L330 (на филогенетическом дереве – общий корень линий YP1102 и YP771), однако сравнение полученных данных проведено преимущественно в масштабе древнего населения других регионов и эпох.
Пожалуй, единственным примером изучения линий высокого разрешения внутри гаплогруппы Q-M242 в Южной Сибири является работа исследователей из Томского НИМЦ, представленная на одной из конференций [8]. В.Н. Харьковым и коллегами было изучено распространение двух относительно недавних линий (Q-B31 и Q-B33) в Западной и Южной Сибири. Исследователи пришли к заключению, что территория Тувы являлась источником происхождения данных линий Y-хромосомы, причем этот общий генетический компонент находит параллели и в распространении аутосомных маркеров.
Мы поддерживаем гипотезу южносибирского происхождения тех ветвей гаплогруппы Q, которые распространены одновременно и в Южной, и в Центральной, и в Западной Сибири. Для более глубокого изучения этого вопроса необходимо подробное изучение распространения различных ветвей гаплогруппы Q в популяциях Южной Сибири. До настоящего времени данных генотипирования популяций Южной Сибири по панели маркеров, основанной на подробной филогении гаплогруппы Q-M242, в научной литературе не представлено. Поэтому целью данного исследования было изучение филогенетической структуры и филогеографического паттерна распространения различных линий гаплогруппы Q-M242 в популяциях Южной Сибири и смежных регионов, а средством достижения этой цели стало генотипирование более полусотни SNP-маркеров, отражающих глубокую филогенетическую структуру гаплогруппы, в сопоставлении с данными по STR-маркерам.
МАТЕРИАЛЫ И МЕТОДЫ
Изучены 545 образцов ДНК из пяти популяций Саянского нагорья, представляющих разные географические группы: западных, центральных, южных и юго-восточных тувинцев, а также тоджинцев, выборки охарактеризованы в табл. 1. Образцы были собраны Л.Д. Дамба под руководством Е.В. Балановской согласно правилам Биобанка Северной Евразии [9] в экспедиционных обследованиях 2015–2017 гг. как описано ранее [1–3, 10, 11]. В выборки включены только неродственные между собой индивиды, все предки которых на протяжении не менее трех поколений относили себя к данному этносу и родились на данной территории. Сбор образцов сопровождался письменным информированным согласием обследуемых под контролем Этической комиссии ФГБНУ “МГНЦ”.
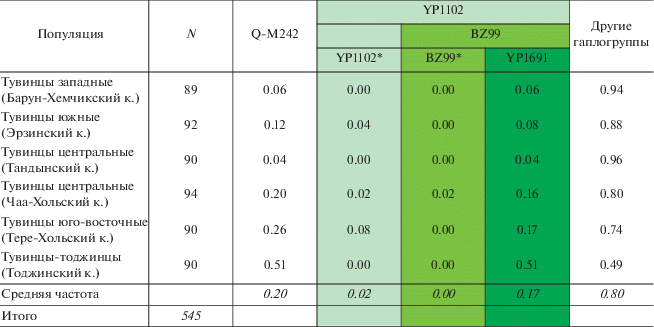
Частоты основных гаплогрупп Y-хромосомы в популяциях тувинцев были представлены ранее [2], в том числе было показано, что гаплогруппа Q-M242 составляет заметную часть генофонда саянских популяций. В данной работе проведено углубленное изучение гаплогруппы Q у тувинцев и окружающих популяций Сибири на основе генотипирования 58 SNP-маркеров, маркирующих разные филогенетические линии внутри гаплогруппы Q-M242. Для субгаплогруппы YP1102, выявленной у тувинцев, проведено генотипирование той же панели 58 маркеров у окружающих популяций, суммарная проанализированная выборка носителей Q-YP1102 составила 394 образца. Из них 136 образцов были дополнительно генотипированы по панели STR-маркеров.
Материалом для исследования послужила геномная ДНК, выделенная из образцов венозной крови методом фенол-хлороформной экстракции и с помощью автоматизированной системы QiaSymphony.
Принадлежность образцов ДНК к линиям внутри гаплогруппы Q-M242 Y-хромосомы определялась по данным анализа 58 SNP-маркеров: B143, B280, B31, BZ1000, BZ1030, BZ1499, BZ1520, BZ3140, BZ321, BZ3908, BZ427, BZ431, BZ664, BZ99, F1893, L275, L330, L332, L53, L54, L712, L713, L804, L940, M25, M3, M378, SK1995, Y1230, Y12452, Y18091, Y2137, Y2200, Y2211, Y2659, Y2677, Y2990, Y5186, Y521, Y558, Y661, YP1095, YP1102, YP1226, YP1669, YP1690, YP1691, YP3952, YP4000, YP4004, YP4055, YP4385, YP4387, YP745, YP771, YP812, Z35990, Z36048. Генотипирование проведено с помощью технологии OpenArray на приборе QuantStudio 12KFlex, позволяющей проводить генотипирование методом ПЦР в реальном времени для большого числа образцов по всем SNP-маркерам. Все перечисленные SNP были включены в кастомную OpenArray панель.
Фрагментный анализ 17 STR-локусов проведен на приборе ABI 3130xl (Applied Biosystems) с использованием набора Y-filer PCR Amplification Kit (Applied Biosystems). Для сравнения привлечены гаплотипы южных алтайцев, хакасов, изученные в [12], а также наши данные для сибирских татар и народов Монголии.
Филогенетическая сеть построена в программе Network 10.2.0.0 [13] по данным о 15 STR-маркерах (DYS389I, DYS389II, DYS390, DYS456, DYS19, DYS458, DYS437, DYS438, DYS448, GATAH4, DYS391, DYS392, DYS393, DYS439, DYS635) и по трем SNP-маркерам (YP1102, BZ99, YP1691; использовались с более высоким значением веса), оформлена в программе Network Publisher v 2.1.2.5 [13].
Возраст кластеров на основе STR-маркеров оценен по числу мутационных шагов (ρ-статистика) [14] с использованием длины поколения 31 год [15] и двух скоростей мутирования: “генеалогической” и “комбинированной”. “Генеалогическая” скорость (0.027 мутаций на локус за поколение) учитывает накопление мутаций в среднем для разных STR-локусов Y-хромосомы, ее значение было получено в исследованиях при прямом подсчете мутаций в известных родословных [16]. “Комбинированная” скорость (0.042 мутаций на локус за поколение) представляет собой сумму скоростей мутирования для каждого из проанализированных 15 STR-локусов, данные получены с ресурса YHRD.org (Available at: https://yhrd.org/ pages/resources/mutation_rates. Accessed: 25.08.2020). Датировки для линий YP1102, BZ99, YP1691 приводятся по данным компании Yfull (рассчитаны при анализе полных последовательностей Y-хромосом).
Также был проведен анализ опубликованных данных о древней ДНК с гаплогруппой Q-M242. Были проанализированы BAM файлы из четырех работ [5, 6, 17, 18] и был установлен итоговый генотип до уровня маркера YP1691. Для двух индивидов из работ [5, 6] принадлежность к линии YP1102(xB31, Y12452, BZ99) показана в [19].
Карты распространения линий гаплогруппы Q-M242 построены с помощью оригинального картографического пакета “GeneGeo” [20] методом средневзвешенной интерполяции с радиусом влияния 1000 км и значением весовой функции, равным 3 (подробное описание технологии и процесса создания карт приведено в [21]).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Разнообразие ветвей гаплогруппы Q-M242 в популяциях Саян
Результаты анализа 58 SNP-маркеров внутри гаплогруппы Q-M242 выявили в генофонде тувинцев и тоджинцев только три линии – YP1102(xB31, Y12452, BZ99), BZ99(xZ35990, YP1691) и YP1691, восходящие к одному корню – варианту YP1102 (табл. 1). Время появления SNP-мутации YP1102, оцененное по полногеномным данным, составляет около 4.6 тыс. лет назад [19].
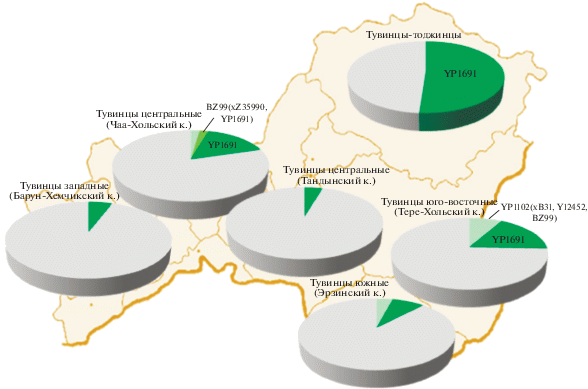
Наиболее частой из трех является линия YP1691: она охватывает 17% генофонда тувинцев в целом, при этом достигая максимума на северо-востоке у тоджинцев (51%), сохраняя частоту, близкую к средней, у юго-восточной и центральной групп (Тере-Хольского и Чаа-Хольского кожуунов Тувы) и снижаясь в юго-западном направлении (до 8% – у южных тувинцев, 6% – у западных и до 4% – у популяции центральных тувинцев Тандынского кожууна; рис. 1).
Наиболее редкой из трех является ветвь BZ99(xZ35990, YP1691), которая вместе с YP1691 восходит к общему SNP-маркеру BZ99. Линия BZ99(xZ35990, YP1691) встречена только у двух представителей центральной группы тувинцев Чаа-Хольского кожууна (2%), она может включать в себя один или два пока неизвестных варианта (запись “BZ99(xZ35990, YP1691)” обозначает генотип, относящийся к “материнской” линии BZ99, но исключающий принадлежность к “дочерним” линиям Z35990 и YP1691; далее по тексту упоминается в виде BZ99*).
Наконец, третья из обнаруженных ветвей – линия YP1102(xB31, Y12452, BZ99), далее по тексту упоминается как YP1102* – встречается у трех из шести популяций тувинцев: центральных Чаа-Хольского кожууна (2%), южных (4%) и юго-восточных (8%, рис. 1). Частота данной линии сравнительно невелика, но при этом, как и для гаплогруппы Q‑M242 в целом, прослеживается понижение доли YP1102* у тувинцев в направлении с востока на запад.
Наличие двух линий гаплогруппы Q-M242 на разных концах ареала тувинцев может указывать на их наследование от одной прапопуляции, в которой могли быть представлены и предковая ветвь YP1102, и разные варианты внутри нее. В таком случае присутствие только одной линии YP1691 у тувинцев-тоджинцев (где гаплогруппа Q-M242 охватывает половину генофонда) можно полагать следствием дрейфа генов в изолированной популяции, издревле проживающей в горно-таежной местности. В пользу этого свидетельствует наличие двух ветвей гаплогруппы Q (YP1102* и YP1691) у южных соседей тоджинцев – юго-восточных и южных тувинцев. Иными словами, у древнего населения восточных областей Тувы, на основе которого сформировались тоджинцы, юго-восточные и южные тувинцы, в генофонде могли быть представлены обе линии гаплогруппы Q, но впоследствии у тоджинцев в результате дрейфа генов сохранилась (и значительно увеличилась в частоте) только одна – линия YP1691. Однако подобное отразилось бы на внутренней структуре линии YP1691 в популяции тоджинцев, например привело бы к снижению разнообразия STR-гаплотипов и увеличению числа носителей одного гаплотипа. На представленной далее филогенетической сети рассмотрено разнообразие STR-гаплотипов Y-хромосомы в пределах линии YP1102 не только у разных групп тувинцев, но среди и носителей этого маркера из других популяций Сибири.
Филогенетический анализ Q-YP1102 по STR-маркерам
Популяционный скрининг маркеров внутри гаплогруппы Q-M242 в популяциях Северной Евразии позволил обнаружить линию YP1102 (и три ветви внутри нее, встреченные у тувинцев) среди других народов Сибири. Результаты филогенетического анализа для носителей линий YP1102*, BZ99* и YP1691 (рис. 2) представили возможность изучить их разнообразие на основе другой системы генетических маркеров – “быстромутирующих” STR Y-хромосомы.
Рис. 2.
Филогенетическая сеть гаплогруппы Q-YP1102 в населении Сибири. а – принадлежность гаплотипов к субтипам гаплогруппы Q (согласно легенде слева); б – принадлежность носителей гаплотипов к популяциям Сибири (согласно легенде слева).
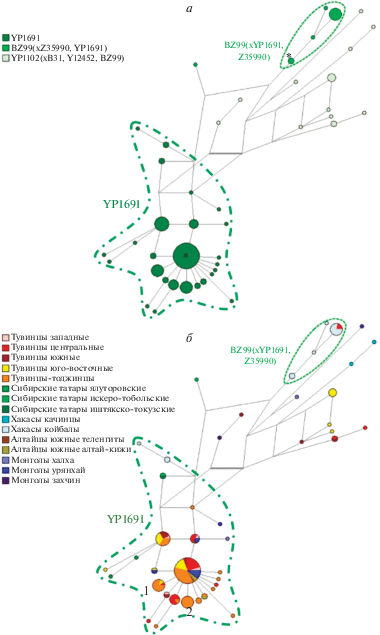
Филогенетическая сеть (рис. 2) представлена в двух вариантах: в верхнем блоке (рис. 2,а) разными цветами показаны гаплотипы носителей каждой из трех линий гаплогруппы Q-M242, в нижнем блоке (рис. 2,б) разные цвета обозначают популяции, из которых происходят носители гаплотипов. На данной сети (рис. 2,а) выделяются два основных кластера, соответствующих двум ветвям гаплогруппы Q-M242: более обширный YP1691 и небольшой по числу носителей BZ99*. Гаплотипы носителей третьей линии YP1102* рассеяны на филогенетической сети и отчетливой кластеризации для них не наблюдается.
Кластер BZ99* включает преимущественно гаплотипы хакасов-койбалов, но также – и двух тувинцев центральной группы из Чаа-Хольского кожууна (рис. 2,б). Возвращаясь к ранее выдвинутому вопросу, можно сделать вывод: линия BZ99* у тувинцев – это одна ранее неизвестная ветвь (не две), и встречается она также у хакасов. Время формирования данного кластера составляет около 1.5 тыс. лет (средняя двух оценок, табл. 2), а время возникновения самой SNP-мутации BZ99 – около 3.9 тыс. лет назад (на основе полногеномных данных компании Yfull [19]). Возможно, что эта ветвь проникла в генофонд тувинцев при контактах с хакасами, либо же является наследием древней популяции, проживавшей в ареалах обоих народов.
Таблица 2.
Датировки кластеров STR-гаплотипов на филогенетической сети
| Кластер | Состав кластера | Количество образцов в кластере | Количество гаплотипов в кластере | Специфичность кластера (Sp) | Возраст, число мутаций (ρ ± σ) | Возраст кластера, лет (по STR-гаплотипам, “генеалогическая” скорость) | Возраст кластера, лет (по STR-гаплотипам, “комбинированная” скорость) | Возраст SNP-линии, лет (по Yfull) |
|---|---|---|---|---|---|---|---|---|
| YP1691 | Тувинцы | 106 | 26 | 81% | 1.04 ± 0.35 | 1191 ± 400 | 771 ± 260 | 2300 ± 800 |
| BZ99* | Хакасы | 13 | 4 | 85% | 1.62 ± 1.10 | 1855 ± 1260 | 1200 ± 815 | 3900 ± 900 |
Более обширный кластер YP1691 (рис. 2,б) кроме тувинцев включает гаплотипы сибирских татар, южных алтайцев, хакасов, народов северо-запада Монголии – урянхайцев, халха, захчин. Данный кластер звездчатой формы, с выраженным гаплотипом основателя (довольно частым: 39 носителей, представлен почти у всех популяций из кластера). Датировка кластера YP1691, рассчитанная на основе данных об STR-маркерах, составляет в среднем около 1 тыс. лет (средняя двух оценок, табл. 2), а датировка появления SNP-мутации YP1691 – около 2.3 тыс. лет назад (на основе полногеномных данных компании Yfull [19https://www.yfull.com/tree/Q-YP1102/]).
Более половины гаплотипов кластера YP1691 (14 из 26, в том числе – и гаплотип основателя) встречено у тувинцев-тоджинцев (рис. 2,б). При этом заметное накопление образцов в пределах одного гаплотипа прослеживается только для двух групп (обозначены цифрами 1 и 2 на рис. 2,б). Гаплотип 1 (N = 9) встречен у двух тувинцев из центральной и юго-восточной групп, а среди тувинцев-тоджинцев – у семи индивидов, четыре из которых представляют род бараан, другие носители – рода маады, чооду, ак. Иными словами, накопление этого гаплотипа у тувинцев-тоджинцев связано именно с принадлежностью большинства носителей к одной родовой группе. Гаплотип 2 (N = = 9) встречен только у тувинцев-тоджинцев, в пяти родовых группах: кол (N = 3), хойук (N = 2), ак (N = 2), единичные – бараан и чооду. В данном случае некоторое увеличение числа носителей также наблюдается в пределах первых трех родовых групп.
Отметим, что и в других популяциях тувинцев из кластера YP1691 встречаются носители нескольких разных гаплотипов. Например, у западных тувинцев, где общая частота линии YP1691 составляет лишь 6% (пять носителей в популяции), встречено четыре разных гаплотипа (рис. 2,б); аналогичная закономерность прослеживается для юго-восточных, южных и центральных групп.
Иными словами, во всех изученных популяциях тувинцев сохраняется заметное разнообразие гаплотипов и отчетливого указания на последствия дрейфа генов (как причины повышения частоты линии) не наблюдается. Напротив, структура кластера YP1691 (много производных гаплотипов, отходящих от основателя) отражает скорее последствия демографического роста в прапопуляции, из которой данная линия гаплогруппы Q проникла в генофонды не только разных территориальных групп тувинцев, но и других народов Сибири. Например, самый частый гаплотип (он же – “основатель” кластера YP1691) встречается, кроме тувинцев, у южных алтайцев (двух алтай-кижи, одного теленгита) и урянхайцев Монголии (N = 4, таблица Приложения), а производные от основателя гаплотипы распространены еще шире (среди хакасов и сибирских татар).
Носители линии YP1691 из популяций сибирских татар и хакасов, находясь в составе одноименного кластера, тем не менее отдалены от гаплотипа основателя и ближайших к нему (рис. 2,б). Возможно, что при увеличении числа образцов из популяций Западной Сибири на филогенетической сети может проявиться отдельный – специфичный для них – кластер.
Филогеографический анализ распространения Q-YP1102 в Сибири
Карты распространения рассматриваемых ветвей гаплогруппы Q-M242 в Сибири представлены на рис. 3: линии YP1102 в целом (рис. 3,а) и ее ветвей (рис. 3,б–г).
Рис. 3.
Филогенетическая схема гаплогруппы Q-M242 с указанием датировок (по данным Yfull) и карт распространения в Сибири трех изученных линий. а – карта суммарной частоты линии YP1102; б – карта распространения линии YP1102*; в – карта распространения линии BZ99*; г – карта распространения линии YP1691. На всех картах изученные популяции тувинцев показаны красными точками, все группы сравнения (современное население), для которых проведен скрининг той же панели маркеров, – черными точками. Значение частоты генотипа на карте соответствует цветовой шкале в верхнем поле каждой карты. На карте линии YP1102* звездами разного цвета обозначены древние популяции, у представителей которых найдена данная ветвь гаплогруппы Q, расшифровка данных обозначений приведена на рисунке слева (датировки и периоды, названия археологических культур и стоянок, ссылки на источник данных).
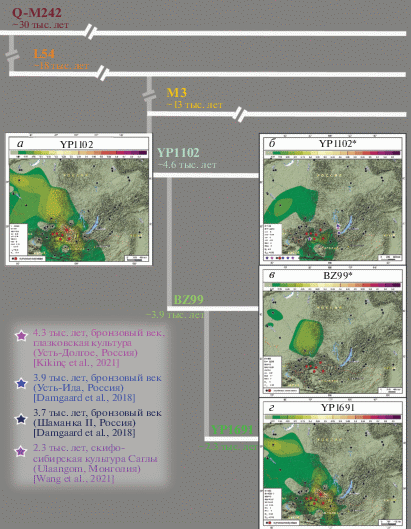
Самая недавняя по датировке (2.3 тыс. лет; табл. 2) и частая у тувинцев ветвь YP1691 выявлена нами и у многих других народов (рис. 3): на северо-западе Монголии (урянхайцы, частота 18%), в Южной Сибири (алтай-кижи – 8%, теленгиты – 1.5%, хакасы – 2.5%) и Западной Сибири (тоболо-иртышские и барабинские сибирские татары – до 3.5%, ханты – 13%).
Ветвь BZ99* в Сибири обнаруживается только на отдельных территориях, не образующих непрерывного ареала (рис. 3): с наибольшей частотой встречена у хакасов (9%), у тоболо-иртышских татар составляет 3%, у хантов – 1%. Напомним, что BZ99* – самая редкая линия гаплогруппы Q у тувинцев: она встречена только в центральной популяции Чаа-Хальского кожууна у двух индивидов (рис. 1,а).
Базальная ветвь YP1102* за пределами ареала тувинцев встречается с низкой частотой: у народов Монголии (захчин, урянхайцы, халха) – с частотой 1–3%, у хакасов – 2%, у барабинских сибирских татар – 3% (рис. 3).
Итак, паттерн трех карт достаточно близок: хотя бы с низкими частотами, но каждая ветвь представлена и в Южной, и в Западной Сибири. Ареал ветвей YP1102* и BZ99* входит в область распространения самой молодой ветви YP1691, поэтому обобщенная карта распространения всей линии YP1102 незначительно отличается от таковой для YP1691 (рис. 3).
В научной литературе о географии линии YP1102 и ее ветвей представлены только фрагментарные данные. О присутствии линии YP1102 в генофонде кетов и селькупов – коренного населения междуречья Оби и Енисея – известно из работы [4] по данным полногеномного анализа. О присутствии линии B33 (ветвь внутри линии YP1691 [ISOGG]) в генофонде кетов, селькупов и ряда популяций Южной Сибири и ее корреляции с “енисейским” генетическим компонентом изложено в сообщении В.Н. Харькова и соавт. [8]. Таким образом, представленные в данном исследовании карты и сведения из научной литературы указывают на распространение и ветви YP1691, и всей линии YP1102 на территории также Центральной Сибири. Принимая во внимание датировку ветви YP1691 (около 2.3 тыс. лет), следует предполагать, что ее носители расселились столь широко (территории Южной, Западной и Центральной Сибири) в течение последних двух тысячелетий. Менее очевидно, какая территория явилась источником распространения этой ветви. Присутствие в генофонде и тувинцев, и хакасов всех трех ветвей (YP1102*, BZ99*, YP1691) может указывать на Южную Сибирь как прародину всей линии YP1102.
Данные NGS-анализа Y-хромосом из древних популяций Сибири позволили обнаружить базальную ветвь YP1102* у пяти мужчин: четырех представителей бронзового века Прибайкалья (образцы DA339, DA343, DA356 из работ [17, 18], образец irk022 из [5]) и одного индивида из могильника культуры Саглы скифо-сибирского типа на границе Монголии и Тувы [6]. На карте ветви YP1102* обозначены места археологических стоянок, где были обнаружены носители данной линии гаплогруппы Q-M242 (рис. 2,б), и датировки (округленные значения радиоуглеродных дат, приведенных в указанных публикациях). Иными словами, линия YP1102* присутствовала среди древнего населения Сибири разного географического положения и разных периодов времени.
Можно сделать вывод, что большинство известных древних представителей ветви YP1102* проживало хотя и за пределами современного распространения данной линии Y-хромосомы, но в регионе, непосредственно примыкающем к ее ареалу – на территории Прибайкалья, около 4 тыс. лет назад (рис. 3,б). Три из четырех индивидов (DA343, DA356, irk022) происходят из населения глазковской археологической культуры [5, 18].
Пятый носитель линии YP1102* – представитель скифского мира и населения, проживавшего почти в пределах современного ареала тувинцев более 2 тыс. лет назад (рис. 2,б). О наличии линии YP1102* на данной территории еще раньше – в бронзовом веке – пока можно только предполагать.
Присутствие в населении Южной Сибири базальной линии YP1102* в течение последних 4 тыс. лет (документированной на этой территории и около 4 тыс. лет назад, и около 2 тыс. лет назад, и в настоящее время), широкая представленность ветвей BZ99* и YP1691 в современных популяциях региона (преимущественно у тувинцев) могут указывать на вероятное происхождение всей линии YP1102 из данного ареала. В таком случае наличие линии YP1102 (и ее ветви YP1691) в генофонде современных популяций кетов и селькупов может указывать на происхождение части их предков из древнего населения Южной Сибири. Это согласуется с лингвистическими данными о территориально более южном происхождении народов енисейской языковой семьи [22], единственным современным представителем которой являются кеты.
Структура кластера YP1691 на филогенетической сети свидетельствует о росте численности населения Южной Сибири позднего железного века. Широкий ареал линии YP1691, вероятно, является следствием активных миграционных процессов в течение последних двух тысяч лет, которые привели к ее распространению из Южной Сибири на север и запад.
Авторы выражают благодарность всем донорам образцов, которые принимали участие в данном исследовании, а также АНО “Биобанк Северной Евразии” за предоставление коллекций ДНК.
Исследование выполнено при финансовой поддержке Министерства науки и образования РФ (Госконтракт № 011-17 от 26.09.2017) в рамках научно-технической программы Союзного государства “ДНК-идентификация” (работы по генотипированию панелей SNP-маркеров, картографический анализ), Государственного задания Министерства науки и высшего образования РФ для Медико-генетического научного центра им. акад. Н.П. Бочкова (работы по генотипированию STR-маркеров, филогенетический анализ), Государственного задания Министерства науки и высшего образования РФ для Института общей генетики им. Н.И. Вавилова РАН (интерпретация результатов, написание текста статьи).
Все процедуры, выполненные в исследовании с участием людей, соответствуют этическим стандартам институционального и/или национального комитета по исследовательской этике и Хельсинкской декларации 1964 г. и ее последующим изменениям или сопоставимым нормам этики.
От каждого из включенных в исследование участников было получено информированное добровольное согласие.
Авторы заявляют, что у них нет конфликта интересов.
Список литературы
Дамба Л.Д., Балановская Е.В., Агджоян А.Т. и др. Генофонд трех восточных родов тувинцев по данным полиморфизма Y-хромосомы // Вестник Моск. ун-та. Серия XXIII. Антропология. 2019. № 1. С. 74–85. https://doi.org/10.32521/2074-8132.2019.1.074-085
Балановская Е.В., Дамба Л.Д., Агджоян А.Т. и др. Генофонд охотников-оленеводов Южной Сибири: тофалары и тоджинцы // Вестник Моск. ун-та. Серия XXIII. Антропология. 2019. № 4. С. 67–80. https://doi.org/10.32521/2074-8132.2019.4.067-080
Жабагин М.К., Дамба Л.Д., Короткова Н.А. и др. Анализ родоплеменной структуры тувинцев по маркерам Y-хромосомы // Генетика. 2020. Т. 56. № 6. С. 732–738. https://doi.org/10.31857/S0016675820060132
Grugni V., Raveane A., Ongaro L. et al. Analysis of the human Y-chromosome haplogroup Q characterizes ancient population movements in Eurasia and the Americas // BMC Biol. 2019. V. 17. № 3. https://doi.org/10.1186/s12915-018-0622-4
Kılınç G.M., Kashuba N., Koptekin D. et al. Human population dynamics and Yersinia pestis in ancient northeast Asia // Sci. Adv. 2021. V. 7(2):eabc4587. https://doi.org/10.1126/sciadv.abc4587
Wang C.C., Yeh H.Y., Popov A.N. et al. Genomic insights into the formation of human populations in East Asia // Nature. 2021. V. 591. P. 413–419. https://doi.org/10.1038/s41586-021-03336-2
Mary L., Zvénigorosky V., Kovalev A. et al. Genetic kinship and admixture in Iron Age Scytho-Siberians // Hum. Genet. 2019. V. 138. P. 411–423. https://doi.org/10.1007/s00439-019-02002-y
Харьков В.Н., Новикова Л.М., Хитринская И.Ю., Степанов В.А. Енисейский генетический субстрат в генофонде коренного населения Южной и Западной Сибири // VII Съезд Вавиловского общества генетиков и селекционеров (ВОГиС), посвященный 100-летию кафедры генетики СПбГУ, и ассоциированные симпозиумы: медж. конгресс (Санкт-Петербург, 18–22 июня 2019). Санкт-Петербург: Изд-во ВВМ, 2019. С. 337.
Балановская Е.В., Жабагин М.К., Агджоян А.Т. и др. Популяционные биобанки: принципы организации и перспективы применения в геногеографии и персонализированной медицине // Генетика. 2016. Т. 52. № 12. С. 1371–1387. https://doi.org/10.7868/S001667581612002X
Дамба Л.Д., Айыжы Е.В., Монгуш Б.Б. и др. Комплексный подход в изучении родовой структуры тувинцев Республики Тыва на примере родов монгуш и ооржак // Вестник Тув. гос. ун-та. Вып. 2. Естеств. и сельскохоз. науки. 2018. № 2(37). С. 37–44.
Дамба Л.Д., Айыжы Е.В., Монгуш Б.Б. и др. Краткий обзор некоторых родоплеменных групп тувинцев по данным междисциплинарных исследований // Вестник Тув. гос. ун-та. Вып. 2. Естеств. и сельскохоз. науки. 2019. № 2(45). С. 19–30. https://doi.org/10.24411/2077-5326-2019-10003
Балаганская О.А. Полиморфизм Y-хромосомы у тюркоязычного населения Алтая, Саян, Тянь-Шаня и Памира в контексте взаимодействия генофондов Западной и Восточной Евразии: Автореф. дис. … канд. биол. наук. М.: МГНЦ РАМН, 2011. 27 с.
Электронный ресурс: http://www.fluxus-engineering.com (дата обращения – 28.12.2020).
Forster P., Harding R., Torroni A., Bandelt H.J. Origin and evolution of Native American mtDNA variation: a reappraisal // Am. J. Hum. Genet. 1996. V. 59. № 4. P. 935–945.
Fenner J.N. Cross-cultural estimation of the human generation interval for use in genetics-based population divergence studies // Am. J. Phys. Anthropol. 2005. V. 128. P. 415–423. https://doi.org/10.1002/ajpa.20188
Балановский О.П., Запорожченко В.В. Хромосома-летописец: датировки генетики, события истории, соблазн ДНК-генеалогии // Генетика. 2016. Т. 52. № 7. С. 810–830. https://doi.org/10.7868/S0016675816070043
Damgaard P.D.B., Marchi N., Rasmussen S. et al. 137 ancient human genomes from across the Eurasian steppes // Nature. 2018. V. 557. P. 369–374. https://doi.org/10.1038/s41586-018-0094-2
Damgaard P.D.B., Martiniano R., Kamm J. et al. The first horse herders and the impact of early Bronze Age steppe expansions into Asia // Science. 2018. V. 360(6396). https://doi.org/10.1126/science.aar7711
Электронный ресурс: Филогенетическое дерево линии Q-YP1102 на сайте компании Yfull https://www.yfull.com/tree/Q-YP1102/ (дата обращения – 28.05.2021).
Кошель С.М. Геоинформационные технологии в геногеографии // Современная географическая картография. М.: Дата+, 2012. С. 158–166.
Балановский О.П. Генофонд Европы. М.: Тов-во науч. изд. КМК, 2015. 354 с.
Vajda E.J. Yeniseian Peoples and Languages: A History of Their Study with an Annotated Bibliography and a Source Guide. England: Curzon Press, 2001. 389 p.
Дополнительные материалы отсутствуют.