

Генетика, 2022, T. 58, № 11, стр. 1225-1239
Дендрогеномика – новая междисциплинарная область исследований адаптивного генетического потенциала лесных древесных популяций, интегрирующая дендрохронологию, дендроэкологию, дендроклиматологию и геномику
К. В. Крутовский 1, 2, 3, 4, *
1 Гёттингенский университет им. Георга-Августа
37077 Гёттинген, Германия
2 Институт общей генетики им. Н.И. Вавилова Российской академии наук
119991 Москва, Россия
3 Сибирский федеральный университет, Институт фундаментальной биологии и биотехнологии
660041 Красноярск, Россия
4 Воронежский государственный лесотехнический университет им. Г.Ф. Морозова
394087 Воронеж, Россия
* E-mail: kkrutovsky@gmail.com
Поступила в редакцию 30.05.2022
После доработки 21.06.2022
Принята к публикации 23.06.2022
- EDN: DHNAPI
- DOI: 10.31857/S0016675822110054
Аннотация
Данный обзор посвящен дендрогеномике, новой междисциплинарной области исследований, интегрирующей дендрохронологию, дендроэкологию, дендроклиматологию, генетику и геномику и позволяющей на основе совместного анализа данных, полученных дендрологическими и геномными методами, исследовать на новом уровне важные биологические проблемы, в частности такие, как временная динамика формирования границы леса, пространственно-временная структура популяций, а также изучение адаптивного генетического потенциала лесных древесных популяций. Данные проблемы особенно актуальны в контексте глобального потепления, которое способствует активному продвижению леса на север и горные высоты, ранее малоприспособленные для древесных растений, а также для понимания достаточно ли наблюдаемого уровня адаптивного генетического потенциала лесных древесных популяций для их выживания в новых климатических условиях.
Этот специальный выпуск журнала посвящен юбилею выдающегося ученого-генетика Юрия Петровича Алтухова, примечательными чертами которого, присущими ему, как и всякому настоящему исследователю, были широкий взгляд, стремление к новому, выход за рамки своей дисциплины. Благодаря ему и под его руководством многие новые междисциплинарные области исследования были заложены и успешно развивались в созданной им лаборатории популяционной генетики, в том числе такая интересная и важная область исследований, как лесная генетика. Будучи его учеником я также всегда стремился следовать этому. В частности, на базе Института фундаментальной биологии и биотехнологии Сибирского федерального университета (СФУ) в г. Красноярске нами был создан один из первых в стране научно-образовательный центр геномных исследований в 2012 г., а через два года, в 2014 г. была создана первая и до сих пор единственная в стране лаборатория лесной геномики. Одно из современных направлений, которое развивается в рамках этой лаборатории совместно с сотрудниками научно-образовательной лаборатории дендроэкологии и экологического мониторинга Хакасского технического института (филиала СФУ), является дендрогеномика – новая междисциплинарная область, интегрирующая дендрохронологию, дендроэкологию, дендроклиматологию, генетику и геномику и позволяющая на основе совместного анализа данных, полученных дендрологическими и геномными методами, исследовать на новом уровне важные биологические проблемы, в частности такие, как временная динамика формирования границы леса, пространственно-временная структура популяций, а также изучение адаптивного генетического потенциала лесных древесных популяций. Данные проблемы особенно актуальны в контексте глобального потепления, которое способствует активному продвижению леса на север и горные высоты, ранее малоприспособленные для древесных растений, а также для понимания достаточно ли наблюдаемого уровня адаптивного генетического потенциала лесных древесных популяций для их выживания в новых климатических условиях.
ГЛОБАЛЬНОЕ ИЗМЕНЕНИЕ КЛИМАТА И АДАПТИВНЫЙ ГЕНЕТИЧЕСКИЙ ПОТЕНЦИАЛ ЛЕСНЫХ ПОПУЛЯЦИЙ
Имеют ли современные лесные популяции адаптивный генетический потенциал, достаточный для адаптации к изменению климата и позволяющий им выжить в условиях более частых и сильных средовых стрессов, таких как катастрофические и повторяющиеся засухи, пожары и инвазии вредителей, является одним из наиболее острых вопросов, связанных с природоохранными мероприятиями и определяющих их стратегию [1–3]. Бореальные (северные) леса, представляющие 30% (1224 млн га) всех лесов мира, особенно важны в этом отношении, и в первую очередь бореальные леса России (главным образом Сибири и Дальнего Востока), составляющие 69% всех бореальных лесов на планете [4]. Благосостояние бореальных экосистем определяется ключевыми системообразующими лесными древесными видами, в первую очередь хвойными, такими как ель, лиственница, пихта и сосна [5]. Поэтому большинство исследований, представленных в этом миниобзоре, сосредоточены в первую очередь на этих видах. Для данных видов, как и для большинства хвойных, характерны высокая фенотипическая пластичность и высокий уровень генетической изменчивости. Поэтому вызывает огромный интерес как эта изменчивость влияет на рост и адаптацию древесных видов [6–8].
ДЕНДРОХРОНОЛОГИЯ, ГОДИЧНЫЕ КОЛЬЦА ПРИРОСТА ДРЕВЕСИНЫ И ДЕНДРОФЕНОТИПЫ
Дендрохронология путем изучения изменчивости и датировки годичного прироста древесины изучает влияние экологических факторов и событий в истории лесных популяций на рост и развитие деревьев [9, 10]. Годичные кольца прироста древесины содержат много информации об индивидуальной реакции деревьев на биотические и абиотические факторы окружающей среды, такие как динамика численности вредителей, температура, осадки [11–14]. Они регистрируют засухи, инвазии вредителей и другие значительные изменения среды, которые сильно влияют на годичный прирост [15–26]. Изменения годичного прироста древесины можно синхронизировать по времени с реальными событиями, используя климатические и метеорологические данные. Основываясь на этой информации и индивидуальной реакции дерева, можно выделить разные индивидуальные дендрофенотипы деревьев и прокоррелировать их изменчивость с генетической изменчивостью [27–42].
Наиболее часто используемые параметры дендрофенотипической изменчивости – это величина годового кольцевого прироста (tree ring width – TRW), которая может быть конвертирована в годовой прирост базальной площади (annual basal area increment – BAI), стандартизированная TRW (standardised ring width index – RWI), средняя величина кольцевого прироста (average tree ring width – AvTRW) и ее варианса (variance of tree ring width – VarTRW). Последние два параметра были использованы, например, для изучения как средняя гетрозиготность дерева по совокупности генетических маркеров может влиять на величину прироста как показателя гетерозиса и на стабильность (вариансу) прироста как показателя гомеостаза [27].
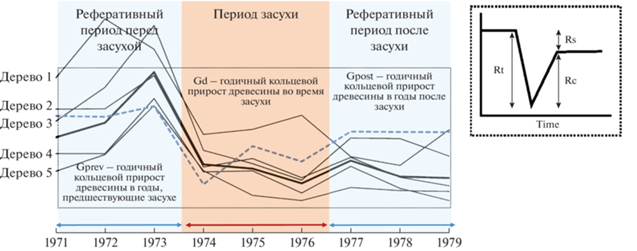
Другие важные индивидуальные дендрофенотипы, предложенные Lloret et al. [43] – это индексы сопротивления Rt (= Gd/Gprev), устойчивости Rs (= Gpost/Gprev) и восстановления Rc (= Gpost/ Gd), где Gprev – средний прирост (измеренный используя либо TRW, либо BAI) в течение двух–трех лет до стресса (например, засухи или инвазии вредителей), Gd – средний прирост в течение стресса и Gpost – средний прирост в течение двух–трех лет после стресса (рис. 1; см. также [44–54] для обзора этих, а также аналогичных индексов). Они отражают индивидуальный ответ дерева на биотический или абиотический стресс и часто используются для анализа стабильности индивидуального прироста деревьев при воздействии засухи (например, [55–59]) и других стрессовых событий, например инвазий вредителей [60–62].
Рис. 1.
Черные линии обозначают годичный кольцевой прирост древесины (TRW – tree-ring width) на примере пяти деревьев лиственницы сибирской из одной из популяций Кузнецкого Алатау в Южной Сибири [57] за период с 1971 по 1979 гг., включая период засухи в 1974–1976 гг. Черная жирная линия обозначает TRW в среднем на популяцию. Голубая прерывистая линия обозначает стандартизированный индекс осадков-эвапотранспирации для района исследования (SPEI – standardized precipitation-evapotranspiration index). Условно обозначены также измерения дендрофенотипов: индексы сопротивления Rt (= Gd/Gprev), устойчивости Rs (= Gpost/Gprev) и восстановления Rc (= Gpost/Gd), где Gprev – средний прирост в течение трех лет до засухи, Gd – средний прирост в течение засухи и Gpost – средний прирост в течение трех лет после засухи.
Например, используя базу данных годичных колец выживших и уже мертвых деревьев из 118 лесонасаждений восьми видов покрытосеменных и 14 голосеменных (>3500 деревьев), в исследовании [63] было показано, что устойчивость роста во время сильных засух и способность к возобновлению роста определяют способность пережить будущие засухи. Было обнаружено, что деревья, которые погибли во время сильной засухи, были менее устойчивы также к предыдущим нелетальным засухам по сравнению с выжившими деревьями того же вида.
ДЕНДРОГЕНОМИКА
Геномика позволяет секвенировать и аннотировать геномы индивидуальных деревьев и генотипировать их по многим генам. В частности, геномы нескольких основных видов хвойных полностью просеквенированы и аннотированы (см. для обзора [64–66], включая сибирскую лиственницу Larix sibirica Ledeb. [67]). Наличие таких геномных данных и референсных геномов позволило разработать большое число высокоизменчивых генетических маркеров, таких как микросателлиты (например, [68–71]). Данные маркеры в основном представляют некодирущие районы и являются селективно-нейтральными. Они очень удобны для изучения пространственной структуры популяций, их дифференциации вследствие изоляции, миграции, потока генов, родства, индивидуального и популяционного уровня изменчивости и там, где важно исключить влияние отбора или можно его игнорировать. Однако данные маркеры не подходят для изучения связи между отдельными генами и контролируемыми ими индивидуальными фенотипами. Полногеномное секвенирование является наиболее информативным методом генотипирования и изучения индивидуальной генетической изменчивости по таким важным функциональным генам. Но из-за огромного размера генома хвойных – от 12 млрд пн (гигабаз – Гб) у лиственницы сибирской [67] до 31 Гб у сахарной сосны, Pinus lambertiana Dougl. [72] – этот метод все еще непозволительно дорог для индивидуального генотипирования большого числа деревьев. В качестве альтернативы используются методы частичного секвенирования генома, такие как таргетное секвенирование (например, [73–76]) и секвенирование районов, связанных с сайтами рестрикции, – restriction-site associated DNA sequencing (RADseq) (например, [28–30, 39, 77–81]).
Возможность индивидуально генотипировать большое число деревьев в популяциях по большому числу генов и генетических маркеров позволяет связать генетическую изменчивость с изменчивостью конкретных дендрофенотипов [27–29, 31–42]. Такой междисциплинарный подход позволяет интегрировать дендрохронологию, дендроэкологию, дендроклиматологию, генетику и геномику и был впервые использован в работах [27–29] и приобрел популярность в последние годы [31–42]. Мы предложили назвать эту новую дисциплину, которая развивает данный подход, дендрогеномикой, а сам термин “дендрогеномика” впервые был предложен нами в статье [30].
Краткий обзор наиболее недавних опубликованных исследований в данной области и некоторых неопубликованных данных автора приведен в табл. 1, и здесь мы выделили только несколько важных проблем, для решения которых дендрогенетика и дендрогеномика оказались очень эффективными. Конечно, ими не исчерпывается весь спектр исследований дендрогеномики. Ограниченный размер журнальной статьи не позволяет описать все исследования в данной области, поэтому автор вынужден ограничиться только несколькими наиболее недавними и характерными примерами и заранее приносит извинения, если какие-то важные исcледования и публикации не вошли в данный миниобзор.
Таблица 1.
Примеры исследований в области дендрогенетики и дендрогеномики
| Популяция (публикация) | Дендрологические и другие данные | Генетические и геномные данные | Основные результаты |
|---|---|---|---|
| Лиственница сибирская (Larix sibirica Ledeb.) | |||
| 100 деревьев из двух популяций в Восточной Сибири [27] | Возраст, ширина кольцевого прироста и его дисперсия | Восемь ядерных микросателлитных маркеров | Не обнаружено достоверной связи между ростом дерева и индивидуальной гетерозиготностью |
| Пять популяций, расположенных через 30–50 км в лесостепной зоне вдоль предгорий Батеневского хребта (Кузнецкий Алатау, Южная Сибирь, Россия), 180 деревьев (35 на популяцию) (неопубл. данные автора*) | Возраст, ширина кольцевого прироста, индивидуальные дендрофенотипы (сопротивления Rt, устойчивости Rs и восстановления Rc), отражающие реакцию роста дерева на засуху для 5 периодов засухи [57] | 44 748 снипов (SNPs), генотипированных в 127 деревьях с помощью ddRAD с использованием двух рестриктаз EcoRI и MseI | 22 снипа коррелировали со средней шириной годичных колец (mean TRW) и 288 – с коэффициентом вариации ширины колец (var TRW); обнаружены также тремя различными методами 356 снипов, частоты которых коррелировали с дендрофенотипами, отражающими ответ на засуху, и проведена детальная аннотация их геномных районов |
| Тсуга горная (Tsuga mertensiana Bong. Carr) | |||
| 163 дерева вдоль градиента высоты, от альпийской границы деревьев до низовья долины в бассейне р. Палмер-Крик в Национальном лесу Чугач на полуострове Кенай, Аляска [28, 29] | Возраст, ширина кольцевого прироста и его дисперсия | 4665 снипов, полученных с помощью ddRADseq с использованием двух рестриктаз SphI и MluCI | Установлена точная возрастная, генетическая, родственная и репродуктивная структура деревьев на границе леса, используя 353 высокоинформативных генетических маркеров-снипов (отобранных из 171 019 снипов); средняя дистанция, на которую распространялись семена, была всего 73 м, однако родство между деревьями было низким; признаков клонального размножения не обнаружено [28]; не обнаружено существенной связи между ростом деревьев и индивидуальной гетерозиготностью, но была значительная положительная связь между средней шириной кольцевого прироста и его дисперсией, подразумевающая, что в целом быстрорастущие деревья в стрессовых условиях, таких как альпийская граница леса, растут нестабильно независимо от уровня индивидуальной гетерозиготности [29] |
| Пихта европейская (Abies alba Mill.) | |||
| 193 дерева из Национального парка “Баварский лес”, переживших сильную засуху в 1976 г. [31] | Возраст, ширина кольцевого прироста, индивидуальные дендрофенотипы (Rt, Rs и Rc), отражающие реакцию роста дерева на засуху | 130 снипов, прогенотипированных в 103 генах, чья изменчивость предположительно связана с ответом на различные стрессы | Изменчивость 15 генов, включая гены фотосинтеза и устойчивости к засухе, была связана с изменчивостью дендрофенотипов |
| Ель европейская (Picea abies L.) | |||
| 328 деревьев на трех постоянных участках с разным лесохозяйственным управлением [32] | Возраст, ширина кольцевого прироста, пространственные данные | Пять ядерных микросателлитных маркеров | Сформировалась сходная пространственная структура с точки зрения возраста деревьев и нейтрального генетического разнообразия, несмотря на различные методы лесохозяйственного управления |
| 147 деревьев из 11 районов происхождения в испытательном участке географических культур Поррау, расположенном за пределами естественного ареала ели обыкновенной на высоте 250 м над уровнем моря на северо-востоке Австрии [35] | Дендроклиматические временные ряды, индивидуальные дендрофенотипы (Rt, Rs, Rc и относительной устойчивости RR), отражающие индивидуальную реакцию роста дерева на засуху; характеристики древесины и взаимосвязи роста с климатом | ~1700 снипов, прогенотипированных с помощью custom Illumina InfiniumHD iSelect BeadChip, включающего 3257 снипов (SNP assays) | Обнаружены значимые ассоциации между 29 снипами и признаками, связанными с засухой, влиянием климата на рост и свойствами древесины, которые объясняли от 11 до 43% фенотипической изменчивости |
| 482 дерева из двух популяций в Южной и Центральной Европе [36] | Возраст, ширина кольцевого прироста, климатические, пространственные данные | 11 несцепленных ядерных микросателлитных маркеров | Генетическое сходство не было связано с изменчивостью прироста древесины, тогда как пространственное расположение деревьев (т.е. особенности микросреды) влияло достаточно сильно |
| Три участка на южной окраине ареала вида в пределах природного заповедника Камполино (северные Апеннины, Италия) и два участка в центральной части ареала вида на территории Баварского национального парка (Богемский массив, Германия); всего 518 взрослых деревьев и 604 саженца [37] | Ширина кольцевого прироста, преобразованная в приращение базальной площади (BAI); 11 дендрофенотипов, сгруппированных в три класса признаков, которые описывают (1) индивидуальный возраст и скорость роста, (2) индивидуальную чувствительность к климату и (3) индивидуальные реакции роста на прошлые экстремальные климатические явления (сопротивление, восстановление и устойчивость); три пространственные переменные; климатические данные | 11 несцепленных ядерных и три хлоропластных микросателлитных локуса, использованных для анализа семейной структуры с целью определения репродуктивного успеха | Изучено влияние дендрофенотипических, пространственных и климатических переменных на индивидуальный рост и репродуктивный успех деревьев, который был положительно связан со скоростью роста и возрастом дерева; на материнский репродуктивный успех также положительно влияла корреляция между ростом и средней температурой предыдущего вегетационного сезона; деревья с наивысшей приспособленностью оказались те, которые способны поддерживать высокие темпы роста, несмотря на потенциальные ограничения роста, вызванные репродуктивными затратами и лимитирующими климатическими условиями |
| Сосна Веймута (Pinus strobus L.) | |||
| 233 дерева из географических культур, представляющих 38 популяций с почти всего ареала вида [33] | Ширина кольцевого прироста, климатические данные | 153 снипа, прогенотипированных с помощью Sequenom iPlex Gold MassARRAY, который включал снипы 52 генов-кандидатов, связанных с ростом и фенологией, ранее идентифицированных у Picea glauca, и 68 генов, случайно выбранных из генома | Обнаруженные генотип–дендрофенотип ассоциации подтвердили 9 генов-кандидатов, идентифицированных ранее в ассоциативном исследовании связи генетической изменчивости с изменчивостью средовых факторов |
| Лиственница Гмелина (Larix gmelinii (Rupr.) Rupr.) | |||
| 194 дерева в северно-центральной части Сибири на юге Таймырского полуострова [84] | Возраст, ширина кольцевого прироста, пространственные данные | 16 ядерных микросателлитных маркеров | Обнаружено, что преобладающим типом размножения был клональный (58.76%) путем распространения рамет на короткие расстояния; наблюдались клональные группы из двух и более особей; возраст некоторых клонов достигает 2200 лет, что совпадает с серьезным изменением окружающей среды, вынудившим отступить линию деревьев в регионе; особи с клональным способом роста, по-видимому, отобраны и поддерживались естественным образом, поскольку это снижает вероятный риск вымирания в суровых условиях, хотя теперь это может быть неадаптивно и препятствовать расширению леса при сильно повышающихся в настоящее время температурах |
| Пихта испанская (Abies pinsapo Boiss.) | |||
| 42 дерева в старом лесонасаждении, созданном в 1910-х гг. в центрально-восточной Испании (~500 км к северу от естественного ареала распространения вида), изученном на предмет его пригодности для ex situ консервации [86] | Ширина кольцевого прироста и дискриминация изотопа углерода 13С (Δ13C) для оценки динамики роста и эффективности использования воды (WUEi) соответственно | Пять ядерных микросателлитных маркеров | Хотя генетическое разнообразие данного лесонасаждения было наполовину ниже такового в естественных популяциях, оно имело важное значение для сохранения этого эндемичного вида, учитывая его хороший многолетний рост и физиологию |
| Ель канадская, или ель белая (Picea glauca (Moench) Voss) | |||
| 2203 дерева, представляющих участки с разными экологическими условиями, включая популяции на границе леса [85] | Пространственные, климатические и дендрохронологические данные | 11 ядерных микросателлитных маркеров | Обнаружено, что особенности микросреды в целом оказали большее влияние на показатели роста, чем генетическое сходство между отдельными деревьями; влияние климата на рост различалось между участками, но было меньше, чем влияние размера дерева; большая внутрииндивидуальная изменчивость ростовых реакций может свидетельствовать о высокой фенотипической пластичности ели белой, которая может амортизировать кратковременные изменения окружающей среды и, таким образом, позволяет выдерживать современные меняющиеся климатические условия |
| 1473 дерева из 197 свободноопыляемых семей, представляющих 43 популяции [38] | Климатические (градиенты температуры, осадков и засушливости), 11 дендрометрических и связанных с засухой признаков (т.е. анатомические признаки, реакция роста и чувствительность к климату) | 6153 снипа в 2606 генах-кандидатах; транскриптомные данные для 37 491 гена | 285 генов были в значительной степени связаны с климатическими факторами или фенотипическими признаками, в том числе 110, которые дифференциально экспрессировались в ответ на засуху в контролируемых условиях теплицы; сеть фенотип-генотип-окружающая среда включала 8 генов, участвующих в адаптации ели к засухе, экспрессия четырех из которых была чувствительна к засухе |
| 1540 деревьев – потомство от открытого опыления трех деревьев, выращенных и тестируемых на нескольких участках генетических испытаний по всей провинции Альберта (Канада) [41] | 15 признаков, связанных с ростом, качеством древесины, засухоустойчивостью и чувствительностью, оцененных на основе данных дендрохронологии и годичных колец, а также уровень монотерпенов (как средство химической защиты от вредителей и фитопатогенов), отражающий стресс, вызванный засухой | 467 224 снипа, генотипированных путем секвенирования (GBS) с использованием рестриктазы EcoT22-I | Отбор по высоте дерева, как правило, увеличивает также диаметр и эффективность использования воды, но снижает плотность древесины, угол микрофибрилл и засухоустойчивость |
| 19-летнее потомство 38 семейств поликросс-тестирования в двух контрастных средах обитания в провинции Квебек, Канада (Québec, Canada) [42] | Возраст, ширина кольцевого прироста, преобразованная в приращение базальной площади (BAI), индивидуальные дендрофенотипы (Rt, Rs, Rc и RR), диаметр на высоте груди (DBH), высота дерева и акустические свойства | 4091 снип, прогенотипированный с помощью Infinium iSelect SNP array | Деревья с более активным ростом в течение своей жизни были более устойчивы к засухе; точность прогнозирования фенотипов устойчивости с помощью геномной селекции (GBLUP) была сопоставима с традиционной селекцией (ABLUP) |
| Нотофагус Домбея (Nothofagus dombeyi (Mirb.) Blume) | |||
| 19 пар деревьев, одна со здоровой (HC), а другая с частично пораженной (PA) кроной на четырех участках в северной Патагонии, Аргентина, пострадавших от засухи [39] | Ширина кольцевого прироста, преобразованная в приращение базальной площади (BAI); дендрофенотипы, отражающие индивидуальную реакцию роста дерева на засуху (Rt, Rs, Rc и RR) | 5224 снипа, генотипированных путем секвенирования (GBS) с использованием рестриктазы ApeKI | 33 адаптивных SNP, дискриминирующих PA от HC деревьев, 8 из которых были связаны с водным стрессом, 6 с дендрофенотипами, и локализованных в генах, кодирующих такие важные белки как большая субъединица хлоропластной рибулозо-1,5-бисфосфаткарбоксилазы/оксигеназы, этилен-чувствительный транскрипционный фактор ERF062 и промотор-связывающий белок SPL |
| Нотофагус макрокарпа (Nothofagus macrocarpa (A.DC.) F.M. Vázquez & R.A. Rodr.) | |||
| 74 дерева в пяти популяциях из центрального региона Чили [40] | Дендрофенотипы, отражающие индивидуальную реакцию роста дерева на засуху (Rt, Rs, Rc и RR); размер дерева | Восемь ядерных микросателлитных маркеров | Различия по дендрофенотипам в основном объясняются влиянием размера деревьев и месторасположения, а не генетическим разнообразием; наблюдаемые различия в устойчивости деревьев к внешним воздействиям между участками можно объяснить значительной фенотипической пластичностью, обусловленной генетически разнообразными генофондами |
Динамика границы леса
Ожидается, что граница леса будет подниматься на более высокие высоты и продвигаться дальше на север в связи с глобальным потеплением [82]. Хотя этот процесс очень гетерогенный [83], важно понимать насколько он устойчив, насколько он связан с миграцией семян и переносом пыльцы, за счет каких деревьев происходит формирование популяций на границе леса, которая фактически является экотоном, какова репродуктивная структура этих популяций. Чтобы ответить на некоторые эти вопросы, мы исследовали дендрохронологически и генетически деревья горной тсуги (Tsuga mertensiana Bong. Carr) вдоль одного из горных склонов на п-ове Кенай, Аляска, определив с помощью кернов их точный возраст и прогенотипировав их по однонуклеотидным полиморфизмам (т.н. “снипам” от английского SNPs, означающего single nucleotide polymorphisms), используя секвенирование ДНК, связанной с сайтами рестрикции двух рестриктаз SphI и MluCI (ddRADseq) [28]. Сочетание двух подходов – дендрохронологического и геномного – позволило установить точную возрастную, генетическую, родственную и репродуктивную структуру деревьев на границе леса, используя 353 высокоинформативных генетических маркера-снипа, отобранных из 171 019 снипов (табл. 1). Средняя дистанция, на которую распространялись семена, была всего 73 м, однако родство между деревьями в популяции было низким, а миграция семян и поток генов высокими.
В исследовании [84] с помощью дендрохронологии и кольцевого прироста был определен возраст 194 деревьев лиственницы Гмелина (Larix gmelinii (Rupr.) Rupr.) на северной границе ареала обитания вида на п-ове Таймыр (север Восточной Сибири), а с помощью 16 полиморфных микросателлитных локусов установлен также их генотип. Обнаружено, что преобладающий тип размножения в этом районе клональный (58.76%) путем распространения рамет на короткие расстояния. При этом возраст клонов часто достигал 2200 лет. Авторы предполагают, что особи с клональным способом размножения поддерживались естественным отбором, поскольку это снижало вероятный риск вымирания в суровых условиях, но теперь эта способность может стать неадаптивной и замедлять расширение границы леса в новых условиях сильно повышающихся в настоящее время температур.
Совместное изучение индивидуальных генетических, дендрохронологических и дендроклиматических данных позволило детально изучить структуру популяции и индивидуальный рост ели канадской, или ели белой (Picea glauca (Moench) Voss) на границе леса в Аляске [85]. Данное исследование показало, что 1) особенности микросреды в целом оказали большее влияние на показатели роста, чем генетическое сходство между деревьями; 2) влияние климата на рост различалось между участками, но было меньше, чем влияние размера дерева; 3) большая внутрииндивидуальная изменчивость ростовых реакций может свидетельствовать о высокой фенотипической пластичности ели белой, которая может амортизировать кратковременные изменения окружающей среды и, таким образом, позволяет выдерживать современные меняющиеся климатические условия.
Пространственно-временная структура популяций
Через изучение динамики кольцевого прироста в сопоставлении с климатическими и эпидемиологическими данными методами дендрохронологии можно понять, как разные абиотические (температура, осадки, аридность и т.д.) и биотические (вспышки вредителей, инвазии фитопатогенов и пр.) средовые факторы могут влиять на рост деревьев. При этом важно вычленить влияние и других факторов, таких, например, как генетическое родство и пространственная структура. В работе [36], используя в комплексе индивидуальные дендрохронологические, генетические и пространственные данные для двух популяций ели обыкновенной (Picea abies L.), было показано, что генетическое сходство не было связано с изменчивостью прироста древесины, тогда как пространственное расположение деревьев (т.е. особенности микросреды) влияло на прирост достаточно сильно.
В комплексном мультидисциплинарном исследовании [32] с использованием дендрохронологических, дендроэкологических и генетических данных было изучено как разные лесохозяйственные мероприятия и методы лесопользования и управление лесными угодьями влияют на пространственно-возрастную структуру лесонасаждений (табл. 1).
Совместный анализ генетических данных и долговременной физиологической информации, полученной на основе изучения годичных колец, был использован для изучения старовозрастного лесонасаждения пихты испанской (Abies pinsapo Boiss.), заложенного еще в 1910-х гг. в центрально-восточной Испании (~500 км к северу от естественного ареала распространения вида), на предмет его пригодности в качестве ex situ популяции в природоохранных целях [86].
Представленные выше примеры мультидисциплинарных дендрогенетических исследований подчеркивают необходимость и эффективность изучения дендрохронологических данных вместе с генетическими на индивидуальном уровне для понимания динамики роста природных популяций и их адаптивного потенциала.
Рост и репродукция деревьев
Корреляция между наблюдаемой индивидуальной гетерозиготностью деревьев, определенной в среднем по большому числу генетических маркеров со средней индивидуальной шириной годичных колец (average tree ring width – AvTRW – прокси гетерозиса), и ее дисперсией (variance of tree ring width – VarTRW – прокси гомеостаза) была изучена у лиственницы сибирской [27] и горной тсуги (Tsuga mertensiana Bong. Carr) [29], чтобы проверить гипотезу о том, что деревья с более высокой индивидуальной гетерозиготностью также будут иметь более высокий и устойчивый прирост, предполагая, что они могут быть более устойчивыми к климату и окружающей среде, в том числе в экстремальных условиях на границе альпийского леса. Результаты показали, что не было существенной связи между ростом дерева и индивидуальной гетерозиготностью. Однако между средней шириной годичных колец и дисперсией прироста годичных колец у горной тсуги наблюдалась значительная положительная взаимосвязь, подразумевающая, что в целом быстрорастущие деревья в стрессовых условиях, таких как альпийская линия деревьев, растут нестабильно независимо от уровня индивидуальной гетерозиготности.
Влияние дендрофенотипических, пространственных и климатических переменных на индивидуальный рост деревьев и их отцовский и материнский репродуктивный успех было изучено в пяти популяциях европейской ели с использованием полиморфных генетических маркеров для анализа семейной структуры, необходимой для оценки индивидуального репродуктивного успеха (табл. 1; [37]). Использованный в этой работе дендрогенетический подход позволил найти функциональные связи между индивидуальными экофизиологическими особенностями деревьев и их эволюционной и адаптивной значимостью.
Генетические механизмы устойчивости к стрессам
Поиск связей между изменчивостью генов и индивидуальной изменчивостью дендрофенотипов помогает понять генетические механизмы устойчивости к стрессам. В одном из первых дендрогенетических исследований использование совместно дендрохронологических и генетических данных для 233 деревьев сосны Веймута (Pinus strobus L.) позволило обнаружить ассоциации между изменчивостью дендрофенотипов, отражающих индивидуальную устойчивость к засухе, и изменчивостью ряда генов [33].
В исследовании [31] на выборке в 193 дерева европейской пихты (Abies alba Mill.) из Национального парка “Баварский лес” (Германия), переживших сильную засуху в 1976 г., была изучена связь между изменчивостью индивидуальных дендрофенотипов (индексов сопротивления Rt, устойчивости Rs и восстановления Rc) и изменчивостью 130 однонуклеотидных полиморфизмов (снипов), прогенотипированных в 103 генах-кандидатах, чья функция предположительно связана также с ответом на различные стрессы. Было обнаружено 15 генов, включая гены фотосинтеза и устойчивости к засухе, чья изменчивость статистически достоверно коррелировала с изменчивостью дендрофенотипов.
Fasanella et al. [39] обнаружили шесть снипов, чья изменчивость ассоциировалась с изменчивостью дендрофенотипов, отражающих индивидуальный ответ деревьев нотофагуса Домбея (Nothofagus dombeyi (Mirb.) Blume) на засуху (табл. 1).
Приведенные примеры использования отражающих индивидуальный ответ деревьев на стресс дендрохронологических и дендроэкологических данных в ассоциативных генетических исследованиях демонстрируют новый эффективный подход для оценки устойчивости деревьев к средовым стрессам и для лучшего понимания генетических механизмов и функциональной адаптации лесных деревьев.
Полногеномный ассоциативный анализ и геномная селекция
Все более интенсивные и частые засухи, связанные с изменением климата, требуют от селекционеров рассматривать засухоустойчивость в качестве важнейшего селекционного признака в селекционных работах, а адаптацию к засухе – приоритетом в усилиях лесоводов по лесовосстановлению [41, 42, 87]. Такие сложные количественные признаки как засухоустойчивость контролируются множеством генов, соответственно селекция должна вестись одновременно по большому числу генов [88, 89]. Полногеномный ассоциативный анализ устойчивости к засухе путем поиска связи между устойчивыми дендрофенотипами и полногеномной генетической изменчивостью позволяет найти эти гены, чья изменчивость связана с засухоустойчивостью [31, 33, 35, 38, 39, 90, 91 ]. Например, значительные связи были обнаружены между 29 снипами и признаками, связанными с засухой, качеством и влиянием климата на рост, в исследованиях 11 географических культур ели европейской в Австрии [35]. Эти связи объясняли от 11 до 43% изменчивости признаков. Большинство этих снипов расположены внутри экзонов генов, наиболее важные из которых экспрессируются преимущественно в тканях камбия и ксилемы. Данное исследование подтверждает высокую адаптивную изменчивость ели европейской в Центральной и Юго-Восточной Европе и демонстрирует как количественные генетические, дендроклиматические и геномные данные могут быть совместно использованы для понимания генетической основы адаптации деревьев к экстремальным климатическим условиям. Обнаруженные связи затем могут быть использованы в геномной селекции для предсказания устойчивых к засухе фенотипов на основе многолокусных генотипов [41, 42, 92] и в геномном редактировании [89, 93, 94].
ЗАКЛЮЧЕНИЕ
Представленный миниобзор демонстрирует, что использование данных дендрохронологии, дендроэкологии и дендроклиматологии совместно с генетическими данными позволяет более полно изучать структуру, динамику и адаптацию лесных древесных популяций. Индивидуальные дендрофенотипы отражают индивидуальный ответ деревьев на средовые факторы и стрессы и могут быть эффективно использованы для изучения генетических механизмов адаптации и лежащих в ее основе индивидуальных генов путем исследования ассоциаций изменчивости большого числа генов с изменчивостью дендрофенотипов. Исследования, проводимые в рамках описанной в данном миниобзоре новой интегративной дисциплины – дендрогеномики, позволяют более эффективно выявлять адаптивные генетические изменения в лесных популяциях. Знание их адаптационного потенциала крайне необходимо для прогнозирования последствий глобального изменения климата и разработки природоохранных и лесовосстановительных программ для смягчения его потенциального негативного воздействия.
Мы призываем к более широкому сотрудничеству дендроэкологов и лесных генетиков для изучения индивидуальных дендрофенотипов совместно с их генотипами, для поиска генетических ассоциаций важных адаптивных признаков с конкретными генами, которые их контролируют.
Автор выражает благодарность Д.Ф. Жирновой и Л.В. Белокопытовой (лаборатория дендроэкологии и экологического мониторинга Хакасского технического института – филиала СФУ, г. Абакан, Российская Федерация) и С.В. Новиковой и Н.В. Орешковой (лаборатория лесной геномики Сибирского федерального университета, г. Красноярск, Российская Федерация) за помощь в подготовке данного обзора.
Работа выполнена при финансовой поддержке Российского фонда фундаментальных исследований в рамках научных проектов № 19-14-00120 “Изучение генетической адаптации деревьев к стрессовым средовым факторам на основе полногеномного и дендрохронологического анализа в контексте глобального изменения климата” и № 22-14-00083 “Дендрогеномное изучение адаптации кедра сибирского в Западном Саяне к экстремальным средовым факторам”.
Настоящая статья не содержит каких-либо исследований с использованием в качестве объекта животных.
Настоящая статья не содержит каких-либо исследований с участием в качестве объекта людей.
Автор заявляет, что у него нет конфликта интересов.
Список литературы
Park A., Puettmann K., Wilson E. et al. Can boreal and temperate forest management be adapted to the uncertainties of 21st century climate change? // Crit. Rev. Plant Sci. 2014. V. 33. № 4. P. 251–285. https://doi.org/10.1080/07352689.2014.858956
Isaac-Renton M., Montwé D., Hamann A. et al. Northern forest tree populations are physiologically maladapted to drought // Nat. Commun. 2018. V. 9. № 5254. P. 1–9. https://doi.org/10.1038/s41467-018-07701-0
Sánchez-Pinillos M., D’Orangeville L., Boulanger Y. et al. Sequential droughts: A silent trigger of boreal forest mortality // Glob. Change Biol. 2022. V. 28. № 2. P. 542–556. https://doi.org/10.1111/gcb.15913
Kellomäki S. Management of boreal forests: Theories and applications for ecosystem services // Background: Management of Forest for Varying Ecosystem Services. Cham, Switzerland: Springer Nature Switzerland AG, 2022. P. 1–9. https://doi.org/10.1007/978-3-030-88024-8_1
Tishkov A. Boreal forests // The Physical Geography of Northern Eurasia / Ed. Shahgedanova M. Oxford; N.Y.: Oxford Univ. Press, 2002. P. 217–233.
Prunier J., Verta J.-P., MacKay J.J. Conifer genomics and adaptation: at the crossroads of genetic diversity and genome function // New Phytol. 2016. V. 209. № 1. P. 44–62. https://doi.org/10.1111/nph.13565
Neale D.B., Wheeler N.C. The Conifers: Genomes, Variation and Evolution. Cham: Springer, 2019. 590 p. https://doi.org/10.1007/978-3-319-46807-5
Amaral J., Ribeyre Z., Vigneaud J. et al. Advances and promises of epigenetics for forest trees // Forests. 2020. V. 11. № 9:976. P. 1–21. https://doi.org/10.3390/f11090976
Vaganov E.A., Hughes M.K., Shashkin A.V. Growth Dynamics of Conifer Tree Rings: Images of Past and Future Environments. Berlin; Heidelberg; N.Y. USA: Springer-Verlag, 2006. 358 p. https://doi.org/10.1007/3-540-31298-6
Arzac A., Fonti A.V., Vaganov E.A. An overview on dendrochronology and quantitative wood anatomy studies of conifers in Southern Siberia (Russia) // Progress in Botany. Berlin; Heidelberg: Springer, 2021. P. 1–22. https://doi.org/10.1007/124_2021_52
Fritts H.C., Vaganov E.A., Sviderskaya I.V., Shashkin A.V. Climatic variation and tree-ring structure in conifers: Empirical and mechanistic models of tree-ring width, number of cells, cell size, cell-wall thickness and wood density // Clim. Res. 1991. V. 1. № 2. P. 97–116. https://www.jstor.org/stable/24863301
Fritts H.C. Tree Rings and Climate. 2nd ed. Caldwell: The Blackburn Press, 2001. 567 p.
Wang T., Ren H., Ma K. Climatic signals in tree ring of Picea schrenkiana along an altitudinal gradient in the central Tianshan Mountains, northwestern China // Trees. 2005. V. 19. № 6. P. 736–742. https://doi.org/10.1007/s00468-005-0003-9
Lloyd A.H., Bunn A.G. Responses of the circumpolar boreal forest to 20th century climate variability // Environ. Res. Lett. 2007. V. 2. № 4:045013. P. 1–14. https://doi.org/10.1088/1748-9326/2/4/045013
Cochran P.H. Examples of mortality and reduced annual increments of white fir induced by drought, insects, and disease at different stand densities // USDA Forest Service, Pacific Northwest Experiment Station, Res. Note PNW-525. Portland, 1998. 19 p. https://doi.org/10.2737/PNW-RN-525
Speer J.H., Swetnam T.W., Wickman B.E., Youngblood A. Changes in pandora moth outbreak dynamics during the past 622 years // Ecology. 2001. V. 82. P. 679–697. https://doi.org/10.1890/0012-9658(2001)082[0679:CI-PMOD]2.0.CO;2
Babushkina E.A., Zhirnova D.F., Belokopytova L.V et al. Response of four tree species to changing climate in a moisture-limited area of South Siberia // Forests. 2019. V. 10. № 11:999. P. 1–18. https://doi.org/10.3390/f10110999
D’Arrigo R., Frank D., Pederson N. et al. 1738 years of Mongolian temperature variability inferred from a tree-ring width chronology of Siberian pine // Geophys. Res. Lett. 2001. V. 28. № 3. P. 543‒546. https://doi.org/10.1029/2000GL011845
Cooke B.J., Nealis V.G., Régnière J. Insect defoliators as periodic disturbances in northern forest ecosystems // Plant Disturbance Ecology, 2nd ed. / Eds Johnson E.A., Miyanishi K. Amsterdam, The Netherlands: Acad. Press, 2020. P. 423–461. https://doi.org/10.1016/B978-0-12-818813-2.00012-5
Rozenberg P., Chauvin T., Escobar-Sandoval M. et al. Climate warming differently affects Larix decidua ring formation at each end of a French Alps elevational gradient // Ann. For. Sci. 2020. V. 77. № 54. https://doi.org/10.1007/s13595-020-00958-w
Belokopytova L., Zhirnova D., Kostyakova T., Babushkina E. Dynamics of moisture regime and its reconstruction from a tree-ring width chronology of Pinus sylvestris in the downstream basin of the Selenga River, Russia // J. Arid Land. 2018. V. 10. № 6. P. 877–891. https://doi.org/10.1007/s40333-018-0025-y
Belokopytova L.V., Zhirnova D.F., Meko D.M. et al. Tree rings reveal the impact of soil temperature on larch growth in the forest-steppe of Siberia // Forests. 2021. V. 12. № 12:1765. P. 1–16. https://doi.org/10.3390/f12121765
Kostyakova T.V., Touchan R., Babushkina E.A., Belokopytova L.V. Precipitation reconstruction for the Khakassia region, Siberia, from tree rings // Holocene. 2018. V. 28. № 3. P. 377–385. https://doi.org/10.1177/0959683617729450
Kostyakova T.V., Belokopytova L.V., Zhirnova D.F. et al. Dendrochronological indication of phyllophages’ outbreaks by larch radial growth in the forest-steppe zone of the Republic of Tyva // Contemp. Probl. Ecol. 2021. V. 14. № 1. P. 37–48. https://doi.org/10.1134/S1995425521010054
Demina A.V., Belokopytova L.V., Zhirnova D.F. et al. Degree of connectivity in reconstructed precipitation dynamics and extremes for semiarid regions across South Siberia // Dendrochronologia. 2022. V. 71. № 125903. P. 1–12. https://doi.org/10.1016/j.dendro.2021.125903
Demidko D.A., Sultson S.M., Mikhaylov P.V., Verkhovets S.V. Influence of weather conditions and climate oscillations on the pine looper Bupalus piniaria (L.) outbreaks in the forest-steppe of the West Siberian Plain // Forests. 2022. V. 13. № 1:15. P. 1–18. https://doi.org/10.3390/f13010015
Babushkina E.A., Vaganov E.A., Grachev A.M. et al. The effect of individual genetic heterozygosity on general homeostasis, heterosis and resilience in Siberian larch (Larix sibirica Ledeb.) using dendrochronology and microsatellite loci genotyping // Dendrochronologia. 2016. V. 38. P. 26–37. https://doi.org/10.1016/j.dendro.2016.02.005
Johnson J.S., Gaddis K.D., Cairns D.M., Krutovsky K.V. Seed dispersal at alpine treeline: An assessment of seed movement within the alpine treeline ecotone // Ecosphere. 2017. V. 8. № 1: e01649. P. 1–15. https://doi.org/10.1002/ecs2.1649
Johnson J.S., Chhetri P., Krutovsky K.V., Cairns D.M. Growth and its relationship to individual genetic diversity of mountain hemlock (Tsuga mertensiana) at alpine treeline in Alaska: combining dendrochronology and genomics // Forests. 2017. V. 8. № 11: 418. P. 1–15. https://doi.org/10.3390/f8110418
Johnson J.S., Krutovsky K.V., Rajora O.P. et al. Advancing biogeography through population genomics // Population Genomics: Concepts, Approaches and Applications / Ed. Om Rajora. Cham: Springer, 2019. P. 539–585. https://doi.org/10.1007/13836_2018_39
Heer K., Behringer D., Piermattei A. et al. Linking dendroecology and association genetics in natural populations: Stress responses archived in tree rings associate with SNP genotypes in silver fir (Abies alba Mill.) // Mol. Eco. 2018. V. 27. № 6. P. 1428–1438. https://doi.org/10.1111/mec.14538
Piotti A., Garbarino M., Avanzi C. et al. Influence of spatiotemporal dynamics on the fine-scale spatial genetic structure of differently managed Picea abies stands // Forests. 2018. V. 9. № 10:622. https://doi.org/10.3390/f9100622
Housset J.M., Nadeau S., Isabel N. et al. Tree rings provide a new class of phenotypes for genetic associations that foster insights into adaptation of conifers to climate change // New Phytol. 2018. V. 218. № 2. P. 630–645. https://doi.org/10.1111/nph.14968
Evans M.E.K., Gugger P.F., Lynch A.M. et al. Dendroecology meets genomics in the common garden: new insights into climate adaptation // New Phytol. 2018. V. 218. № 2. P. 401–403. https://doi.org/10.1111/nph.15094
Trujillo-Moya C., George J.-P., Fluch S. et al. Drought sensitivity of Norway spruce at the species’ warmest fringe: Quantitative and molecular analysis reveals high genetic variation among and within provenances // G3: Genes Genom. Genet. 2018. V. 8. № 4. P. 1225–1245. https://doi.org/10.1534/g3.117.300524
Avanzi C., Piermattei A., Piotti A. et al. Disentangling the effects of spatial proximity and genetic similarity on individual growth performances in Norway spruce natural populations // Sci. Total Environ. 2019. V. 650. Part 1. P. 493–504. https://doi.org/10.1016/j.scitotenv.2018.08.348
Avanzi C., Heer K., Büntgen U. et al. Individual reproductive success in Norway spruce natural populations depends on growth rate, age and sensitivity to temperature // Heredity. 2020. V. 124. P. 685–698. https://doi.org/10.1038/s41437-020-0305-0
Depardieu C., Gérardi S., Nadeau S. et al. Connecting tree-ring phenotypes, genetic associations and transcriptomics to decipher the genomic architecture of drought adaptation in a widespread conifer // Mol. Ecol. 2021. V. 30. P. 3898–3917. https://doi.org/10.1111/mec.15846
Fasanella M., Suarez M.L., Hasbun R., Premoli A.C. Individual-based dendrogenomic analysis of forest dieback driven by extreme droughts // Can. J. For. Res. 2021. V. 51. № 3. P. 420–432. https://doi.org/10.1139/cjfr-2020-0221
Venegas-González A., Gibson-Capintero S., Anholetto-Junior C. et al. Tree-ring analysis and genetic associations help to understand drought sensitivity in the Chilean endemic forest of Nothofagus macrocarpa // Front. For. Glob. Change. 2022. V. 5. № 762347. P. 1–13. https://doi.org/10.3389/ffgc.2022.762347
Cappa E.P., Klutsch J.G., Sebastian-Azcona J. et al. Integrating genomic information and productivity and climate-adaptability traits into a regional white spruce breeding program // PLoS One. 2022. V. 17. № 3: e0264549. P. 1–28. https://doi.org/10.1371/journal.pone.0264549
Laverdière J.-P., Lenz P., Nadeau S. et al. Breeding for adaptation to climate change: Genomic selection for drought response in a white spruce multi-site polycross test // Evol. Appl. 2022. V. 15. № 3. P. 383–402. https://doi.org/10.1111/eva.13348
Lloret F., Keeling E.G., Sala A. Components of tree resilience: effects of successive low-growth episodes in old ponderosa pine forests // Oikos. 2011. V. 120. № 12. P. 1909–1920. https://doi.org/10.1111/j.1600-0706.2011.19372.x
Nikinmaa L., Lindner M., Cantarello E. et al. Reviewing the use of resilience concepts in forest sciences // Curr. For. Rep. 2020. V. 6. № 2. P. 61–80. https://doi.org/10.1007/s40725-020-00110-x
Vicente-Serrano S.M., Quiring S.M., Peña-Gallardo M. et al. A review of environmental droughts: Increased risk under global warming? // Earth-Sci. Rev. 2020. V. 201. № 102953. P. 1–23. https://doi.org/10.1016/j.earscirev.2019.102953
Schwarz J., Skiadaresis G., Kohler M. et al. Quantifying growth responses of trees to drought – a critique of commonly used resilience indices and recommendations for future studies // Curr. For. Rep. 2020. V. 6. № 3. P. 185–200. https://doi.org/10.1007/s40725-020-00119-2
Van Meerbeek K., Jucker T., Svenning J.-C. Unifying the concepts of stability and resilience in ecology // J. Ecol. 2021. V. 109. № 9. P. 3114–3132. https://doi.org/10.1111/1365-2745.13651
Yi C., Jackson N. A review of measuring ecosystem resilience to disturbance // Environ. Res. Lett. 2021. V. 16. № 5: 053008. P. 1–24. https://doi.org/10.1088/1748-9326/abdf09
Castagneri D., Vacchiano G., Hacket-Pain A. et al. Meta-analysis reveals different competition effects on tree growth resistance and resilience to drought // Ecosystems. 2022. V. 25. № 1. P. 30–43. https://doi.org/10.1007/s10021-021-00638-4
Pretzsch H., Río M., Giammarchi F. et al. Changes of tree and stand growth: review and implications // Climate-Smart Forestry in Mountain Regions. Managing Forest Ecosystems. V. 40 / Eds Tognetti et al. Cham, Switzerland: Springer Nature Switzerland AG, 2022. P. 189–222. https://doi.org/10.1007/978-3-030-80767-2_6
Vilonen L., Ross M., Smith M.D. What happens after drought ends: synthesizing terms and definitions // New Phytol. 2022. Online first. https://doi.org/10.1111/nph.18137
Zlobin I.E. Linking the growth patterns of coniferous species with their performance under climate aridization // Sci. Total Environ. 2022. V. 851. № 154971. https://doi.org/10.1016/j.scitotenv.2022.154971
Haberstroh S., Werner C. The role of species interactions for forest resilience to drought // Plant Biol. J. 2022. Online first. https://doi.org/10.1111/plb.13415
Hartmann H., Bastos A., Das A.J. et al. Climate change risks to global forest health: Emergence of unexpected events of elevated tree mortality worldwide // Annu. Rev. Plant Biol. 2022. V. 73. № 25. P. 1–30. https://doi.org/10.1146/annurev-arplant-102820-012804
Serra-Maluquer X., Mencuccini M., Martínez-Vilalta J. Changes in tree resistance, recovery and resilience across three successive extreme droughts in the northeast Iberian Peninsula // Oecologia. 2018. V. 187. № 1. P. 343‒354. https://doi.org/10.1007/s00442-018-4118-2
Van Mantgem P.J., Kerhoulas L.P., Sherriff R.L., Wenderott Z.J. Tree-ring evidence of forest management moderating drought responses: Implications for dry, coniferous forests in the southwestern United States // Front. For. Glob. Change. 2020. V. 3. № 41. P. 1–7. https://doi.org/10.3389/ffgc.2020.00041
Zhirnova D.F., Babushkina E.A., Belokopytova L.V., Vaganov E.A. To which side are the scales swinging? Growth stability of Siberian larch under permanent moisture deficit with periodic droughts // For. Ecol. Manag. 2020. V. 459. № 1:117841. P. 1–10. https://doi.org/10.1016/j.foreco.2019.117841
Mihai G., Alexandru A.M., Stoica E., Birsan M.V. Intraspecific growth response to drought of Abies alba in the Southeastern Carpathians // Forests. 2021. V. 12. № 4:387. P. 1–23. https://doi.org/10.3390/f12040387
Belokopytova L.V., Zhirnova D.F., Krutovsky K.V. et al. Species- and age-specific growth reactions to extreme droughts of the keystone tree species across forest-steppe and sub-taiga habitats of South Siberia // Forests. 2022. (submitted)
Bohner T., Diez J. Tree resistance and recovery from drought mediated by multiple abiotic and biotic processes across a large geographic gradient // Sci. Total Environ. 2021. V. 789. № 147744. P. 1–10. https://doi.org/10.1016/j.scitotenv.2021.147744
Campôa J., Calvão T., Firmino P.N., Pimentel C.S. Disentangling the effects of climate and defoliation on forest growth: The case of an outbreak of a Thaumetopoea pityocampa population with a shifted phenology in a Pinus pinaster monoculture // For. Ecol. Manag. 2021. V. 498. № 119548. https://doi.org/10.1016/j.foreco.2021.119548
Weigandt M., Villacide J., Bianchi E. et al. Growth response of Pinus contorta to the synergy of stress factors: Successive extreme drought events and a population outbreak of Sirex noctilio in NW Patagonia // New Forests. 2022. Online first. https://doi.org/10.1007/s11056-022-09907-z
DeSoto L., Cailleret M., Sterck F. et al. Low growth resilience to drought is related to future mortality risk in trees // Nat. Commun. 2020. V. 11. № 545. P. 1–9. https://doi.org/10.1038/s41467-020-14300-5
Mosca E., Cruz F., Gómez-Garrido J. et al. A reference genome sequence for the European silver fir (Abies alba Mill.): A community-generated genomic resource // G3: Genes Genom. Genet. 2019. V. 9. № 7. P. 2039–2049. https://doi.org/10.1534/g3.119.400083
Sun C., Xie Y.-H., Li Z. et al. The Larix kaempferi genome reveals new insights into wood properties // J. Integr. Plant Biol. 2022. Online first. https://doi.org/10.1111/jipb.13265
Niu S., Li J., Bo W. et al. The Chinese pine genome and methylome unveil key features of conifer evolution // Cell. 2022. V. 185. № 1. P. 204–217. https://doi.org/10.1016/j.cell.2021.12.006
Kuzmin D.A., Feranchuk S.I., Sharov V.V. et al. Stepwise large genome assembly approach: A case of Siberian larch (Larix sibirica Ledeb) // BMC Bioinformatics. 2019. V. 20 (Suppl. 1). № 37. P. 1–12. https://doi.org/10.1186/s12859-018-2570-y
Белоконь М.М., Политов Д.В., Мудрик Е.А. и др. Разработка микросателлитных маркеров сосны кедровой сибирской (Pinus sibirica Du Tour) по результатам полногеномного de novo секвенирования // Генетика. 2016. Т. 52. № 12. С. 1418–1427.
Орешкова Н.В., Путинцева Ю.А., Шаров В.В. и др. Разработка микросателлитных маркеров лиственницы сибирской (Larix sibirica Ledeb.) на основе полногеномного de novo секвенирования // Генетика. 2017. Т. 53. № 11. С. 1278–1284. https://doi.org/10.7868/S0016675817110091
Орешкова Н.В., Бондар Е.И., Путинцева Ю.А. и др. Разработка ядерных микросателлитных маркеров с длинными (трех-, четырех-, пяти- и шестинуклеотидными) мотивами для трех видов лиственницы на основе полногеномного de novo секвенирования лиственницы сибирской (Larix sibirica Ledeb.) // Генетика. 2019. Т. 55. № 4. С. 418–425.
Krutovsky K.V., Putintseva Y.A., Oreshkova N.V. et al. Postgenomic technologies in practical forestry: Development of genome-wide markers for timber origin identification and other applications // For. Eng. J. 2019. V. 9. № 1. P. 9–16. https://doi.org/10.12737/article_5c92016b64af27.15390296
Stevens K.A., Wegrzyn J.L., Zimin A. et al. Sequence of the sugar pine megagenome // Genetics. 2016. V. 204. № 4. P. 1613–1626. https://doi.org/10.1534/genetics.116.193227
Dasgupta M.G., Dharanishanthi V., Agarwal I., Krutovsky K.V. Development of genetic markers in Eucalyptus species by target enrichment and exome sequencing // PLoS One. 2015. V. 10. № 1: e0116528. P. 1–30. https://doi.org/10.1371/journal.pone.0116528
Dasgupta M.G., Parveen A.B.M., Shanmugavel S. et al. Targeted re-sequencing and genome-wide association analysis for wood property traits in breeding population of Eucalyptus tereticornis × E. grandis // Genomics. 2021. V. 113. № 6. P. 4276–4292. https://doi.org/10.1016/j.ygeno.2021.11.013
Lu M., Krutovsky K.V., Nelson C.D. et al. Exome genotyping, linkage disequilibrium and population structure in loblolly pine (Pinus taeda L.) // BMC Genomics. 2016. V. 17. № 730. P. 1–11. https://doi.org/10.1186/s12864-016-3081-8
Lu M., Krutovsky K.V., Nelson C.D. et al. Association genetics of growth and adaptive traits in loblolly pine (Pinus taeda L.) using whole exome-discovered polymorphisms // Tree Genet. Genomes. 2017. V. 13. № 3:57. P. 1–18. https://doi.org/10.1007/s11295-017-1140-1
Parchman T.L., Jahner J.P., Uckele K.A. et al. RADseq approaches and applications for forest tree genetics // Tree Genet. Genomes. 2018. V. 14. № 39. P. 1–25. https://doi.org/10.1007/s11295-018-1251-3
Aguirre N.C., Filippi C.V., Zaina G. et al. Optimizing ddRADseq in non-model species: A case study in Eucalyptus dunnii Maiden // Agronomy. 2019. V. 9. № 9:484. P. 1–21. https://doi.org/10.3390/agronomy9090484
Jia K.-H., Zhao W., Maier P.A. et al. Landscape genomics predicts climate change-related genetic offset for the widespread Platycladus orientalis (Cupressaceae) // Evol. Appl. 2020. V. 13. № 4. P. 666–677. https://doi.org/10.1111/eva.12891
Ulaszewski B., Meger J., Burczyk J. Comparative analysis of SNP Discovery and genotyping in Fagus sylvatica L. and Quercus robur L. using RADseq, GBS, and ddRAD methods // Forests. 2021. V. 12. № 2:222. P. 1–17. https://doi.org/10.3390/f12020222
Varas-Myrik A., Sepúlveda-Espinoza F., Fajardo A. et al. Predicting climate change-related genetic offset for the endangered southern South American conifer Araucaria araucana // Forest Ecol. Management. 2022. V. 504. № 119856.
Grace J., Berninger F., Nagy L. Impacts of climate change on the tree line // Ann. Bot. 2002. V. 90. № 4. P. 537–544. https://doi.org/10.1093/aob/mcf222
Wang Y., Li D., Ren P. et al. Heterogeneous responses of alpine treelines to climate warming across the Tibetan Plateau // Forests. 2022. V. 13. № 5:788. P. 1–13. https://doi.org/10.3390/f13050788
Kruse S., Kolmogorov A.I., Pestryakova L.A., Herzschuh U. Long-lived larch clones may conserve adaptations that could restrict treeline migration in northern Siberia // Ecol. Evol. 2020. V. 10. № 18. P. 10017–10030. https://doi.org/10.1002/ece3.6660
Zacharias M., Pampuch T., Heer K. et al. Population structure and the influence of microenvironment and genetic similarity on individual growth at Alaskan white spruce treelines // Sci. Total Environ. 2021. V. 798. № 149267. P. 1–11. https://doi.org/10.1016/j.scitotenv.2021.149267
Santini F., Shestakova T.A., Dashevskaya S. et al. Dendroecological and genetic insights for future management of an old-planted forest of the endangered Mediterranean fir Abies pinsapo // Dendrochronologia. 2020. V. 63. № 125754. P. 1–11. https://doi.org/10.1016/j.dendro.2020.125754
Matallana-Ramirez L.P., Whetten R.W., Sanchez G.M., Payn K.G. Breeding for climate change resilience: A case study of loblolly pine (Pinus taeda L.) in North America // Front. Plant Sci. 2021. V. 12. № 606908. P. 1–22. https://doi.org/10.3389/fpls.2021.606908
Hu H., Xiong L. Genetic engineering and breeding of drought-resistant crops // Annu. Rev. Plant Biol. 2014. V. 65. № 1. P. 715–741. https://doi.org/10.1146/annurev-arplant-050213-040000
Singh P.K., Indoliya Y., Agrawal L. et al. Genomic and proteomic responses to drought stress and biotechnological interventions for enhanced drought tolerance in plants // Curr. Plant Biol. 2022. V. 29. № 100239. P. 1–13. https://doi.org/10.1016/j.cpb.2022.100239
Wang H., Qin F. Genome-wide association study reveals natural variations contributing to drought resistance in crops // Front. Plant Sci. 2017. V. 8. № 110. P. 1–12. https://doi.org/10.3389/fpls.2017.01110
Pfenninger M., Reuss F., Kiebler A. et al. Genomic basis for drought resistance in European beech forests threatened by climate change // eLife. 2021. V. 10. № e65532. P. 1–17. https://doi.org/10.7554/eLife.65532
Lebedev V.G., Lebedeva T.N., Chernodubov A.I., Shestibratov K.A. Genomic selection for forest tree improvement: methods, achievements and perspectives // Forests. 2020. V. 11. № 11:1190. P. 1–36. https://doi.org/10.3390/f11111190
Joshi R.K., Bharat S.S., Mishra R. Engineering drought tolerance in plants through CRISPR/Cas genome editing // 3 Biotech. 2020. V. 10. № 400. P. 1–14. https://doi.org/10.1007/s13205-020-02390-3
Cao H.X., Vu G.T.H., Gailing O. From genome sequencing to CRISPR-based genome editing for climate-resilient forest trees // Int. J. Mol. Sci. 2022. V. 23. № 2:966. P. 1–26. https://doi.org/10.3390/ijms23020966
Дополнительные материалы отсутствуют.