

Генетика, 2022, T. 58, № 12, стр. 1349-1366
Особенности структуры генофонда и генетическая история саха (якутов)
С. А. Федорова 1, 2, *, Э. К. Хуснутдинова 1, 3
1 Северо-Восточный федеральный университет им. М.К. Аммосова
677013 Якутск, Россия
2 Якутский научный центр комплексных медицинских проблем
677000 Якутск, Россия
3 Институт биохимии и генетики Уфимского федерального исследовательского центра
Российской академии наук
450054 Уфа, Россия
* E-mail: sardaanafedorova@mail.ru
Поступила в редакцию 09.09.2021
После доработки 30.04.2022
Принята к публикации 11.05.2022
- EDN: FZYHOU
- DOI: 10.31857/S0016675822120037
Аннотация
В обзоре суммированы результаты молекулярно-генетических исследований структуры генофонда и генетической истории народа саха (якутов) в контексте данных по антропологии, лингвистике, истории и археологии. Описаны характерные особенности генетической структуры якутской популяции по линиям Y-хромосомы, митохондриальной ДНК и данным полногеномного анализа аутосомных локусов. Рассмотрены состав и соотношение компонентов генофонда различного происхождения и генетические взаимоотношения саха с народами Якутии и соседних регионов. Реконструкция генетической истории популяции позволила ответить на ряд вопросов, касающихся этногенеза якутов.
Изучение формирования народов Сибири, генетические портреты которых имеют уникальные черты, сложившиеся при длительном развитии в условиях относительной изоляции, представляет особый интерес для специалистов в области популяционной генетики. Исследования структуры генофонда и генетических взаимоотношений саха (якутов) с другими народами Сибири были предприняты нами начиная с 2002 г. в сотрудничестве с коллегами из Эстонского биоцентра (Тарту, Эстония) и Университета Поля Сабатье (Тулуза, Франция) [1–17]. К детальному изучению генетической структуры якутского этноса одновременно с нами приступили две группы ученых: 1) из Института медицинской генетики (г. Томск) [18–20]; 2) из Института эволюционной антропологии Общества Макса Планка (Лейпциг, Германия) совместно с Институтом здоровья АН РС(Я) (Якутск, Россия) [21–27]. В обзоре приведены результаты наших исследований в сравнении с данными других групп ученых и представлениями об этногенезе якутов, предложенными ранее антропологами, лингвистами, историками и археологами.
ЭТНОГЕНЕЗ НАРОДА САХА (ЯКУТОВ)
Саха (якуты) – тюркоязычный народ, освоивший субарктические и арктические территории северо-востока Евразии. Общепринято считать, что формирование народа саха на территории современной Якутии происходило путем постепенного смешения тюркоязычных племен, переселившихся с южных регионов на Среднюю Лену, с местными племенами в течение длительного периода времени [28–39]. В отличие от соседних народов Восточной Сибири (тундровые юкагиры, чукчи, кочевые коряки, эвенки, эвены, долганы, нганасаны), занимающихся оленеводством, традиционным занятием для якутов является коневодство и разведение крупного рогатого скота.
По антропологическим показателям саха отличаются от других народов Арктики – юкагиров, эвенов и эвенков (байкальский антропологический тип), чукчей, эскимосов и коряков (арктический антропологический тип) и объединяются вместе с северными бурятами Иркутской области в ангаро-ленский локальный тип центральноазиатской группы североазиатских монголоидов [40, 41]. Согласно исследованиям Л.Ф. Томтосовой по антропологическим признакам с якутами наиболее сходны западные буряты, затем монголы, тункинские буряты и тувинцы [42].
В научной литературе уже давно утвердилось мнение о раннем отделении и обособлении якутского языка от основного тюркского языкового ствола. Предлагаются различные датировки этого размежевания: от II до VIII в. н. э. [30, 32, 43–47]. Известный российский тюрколог А.В. Дыбо считает, что отделение языка саха от общетюркского ствола, вероятно, произошло вскоре после начала нашей эры (примерно во II в. н. э.) [45]. Анализ основного словарного фонда якутского и древнетюркского языков, проведенный Г.Г. Левиным, позволил предположить, что расхождение якутского с древними тюркскими языками началось в IV–V вв., а окончательное отделение от языка орхоно-енисейских памятников датируется началом VIII в. [32]. Согласно расчетам А. Савельева и М. Робитс, якутская ветвь отделилась от пратюркского субстрата ~1500 лет назад [47]. Из современных тюркских языков как более близкие к якутскому рассматриваются “сибирские тюркские языки” (алтайский, хакасский, шорский, сарыг-югурский, тувинский, тофаларский) и киргизский [43–46]. По одной из более поздних лингвистических классификаций якутский язык, наряду с долганским, наиболее близок к подгруппе, включающей хакасский, тувинский, тофаларский и шорский языки [46].
Из нетюркских компонентов в якутском языке отмечается значительный вклад слов монгольского происхождения [30]. Кроме того, в языке народа саха присутствует большой пласт слов, не имеющих соответствий ни в каких современных языках [48], что позволило исследователям предположить существование неизвестного народа, оставившего заметный след в якутском языке. Эвенкийских слов, по мнению А.М. Певнова, специалиста в области документирования и сравнительно-исторического изучения тунгусо-маньчжурских языков, в якутском языке чрезвычайно мало, самодийские, кетские и юкагирские заимствования если и есть, то их и того меньше [48].
По вопросу о времени миграции тюркоязычных предков саха на север во мнениях историков и археологов имеются значительные расхождения. Г.В. Ксенофонтов считал, что саха представляют собой народ смешанного происхождения, включивший в себя три волны тюркоязычных переселенцев. По его представлению, заселение предками народа саха бассейна р. Вилюй началось в конце I в. н. э., вторая волна переселилась на Вилюй и западную половину бассейна Лены из Приангарья начиная с VII–VIII вв. н. э.; наконец, последний этап переселения был вызван падением уйгурского ханства в середине IX в. и усилением монгольских племен, вытеснивших предков саха из прибайкальских степей [28]. “В эпоху возвышения монголов Чингисхана “саха”-якутов в Приангарском крае уже не было. Очевидно, якуты Тыгына переселились в пределы отдаленного Якутского округа незадолго до появления Чингисхана” [28]. А.П. Окладников предполагал два “решающих этапа” в расселении южных предков якутов на север: первый, начавшийся в раннем железном веке, заканчивается к X–XI вв., второй этап относится к XV–XVI вв. [33]. По мнению археолога И.В. Константинова переселение предков саха с Прибайкалья произошло в XV в. в виде компактной группы, представлявшей вполне сложившуюся этническую общность [34]. А.И. Гоголев считает, что массовое переселение тюркоязычных южных предков саха на север произошло в начале II тыс. н. э. и документируется появлением в XIII в. поселений культуры “малых домов”, которую позднее сменила кулун-атахская археологическая культура XIV–XVI вв. [35]. На поселениях кулун-атахцев археологи в большом количестве находят кости крупного рогатого скота и лошадей. Однако археологические исследования последних лет показывают, что эпизодические проникновения скотоводческих групп на Олекму начались гораздо раньше – с III–IV вв. н. э. и в X–XVI вв. в бассейне Средней Лены существовала уже сложившаяся, адаптированная к местным условиям культура кочевников, успешно разводивших лошадей и крупный рогатый скот [38].
До сих пор в академической среде общепринятым является представление, что непосредственными южными тюркоязычными предками народа саха были прибайкальские курыканы [33, 34]. Впервые теория о расселении курыкан из Прибайкальской области на север, вниз по р. Лене и участие их в формировании якутской народности выдвинута А.П. Окладниковым [33]. Курыкане были самым дальним и самым северным из телесских племен, проживавших на окраине древнетюркских каганатов, отдельные упоминания о которых зафиксированы в древнетюркских и китайских летописях [33, 49]. С курыканами связывают курумчинскую культуру циркумбайкальского региона (V–XIV вв.), называемую иначе культурой “курумчинских кузнецов” из-за большого количества предметов железоделательного производства, обнаруженных в их поселениях [33, 35, 49]. Помимо курумчинцев, как гипотетических южных предков народа саха позднее стали также рассматривать носителей усть-талькинской культуры Прибайкалья XII–XIV вв. [50, 51].
Долгое время среди исследователей господствовало сформированное историографией середины XIX в. мнение, что автохтонные племена, с которыми встретились южные предки якутов, были представлены предками современных юкагиров и тунгусов, хотя лингвистические данные указывают на слабое влияние эвенкийского и юкагирского языка на якутский [30–32, 48]. В настоящее время существуют иные точки зрения, согласно которым кроме указанных этносов наиболее древний аборигенный пласт в этногенезе якутов могли составлять прасамодийские и другие протоазиатские племена, этнонимы которых неизвестны [36, 38, 52]. Археолог А.Н. Алексеев считает, что в становление генофонда якутского народа значительный вклад был внесен древним дотунгусским населением Якутии, отуреченным в конечном счете в результате миграции полиэтнических групп южных скотоводов [36, 38]. Исторические предания свидетельствуют о достаточном разнообразии аборигенного и мигрировавшего населения той эпохи [53].
Таким образом, при рассмотрении этногенеза народа саха специалистами в области гуманитарных наук недостаточно изученными являются вопросы о времени миграции тюркоязычных предков саха на север, количестве миграционных волн, соотношении пришлых и автохтонных элементов в генофонде якутов, идентификации племен, принимавших участие в сложении якутского народа и степени генетического родства с другими современными этносами Сибири. Наиболее перспективным подходом для решения некоторых из этих вопросов являются исследования в области этногеномики (популяционной геномики), позволяющие реконструировать генетическую историю этноса на основе изучения полиморфизма генома человека в различных популяциях.
ОСОБЕННОСТИ СТРУКТУРЫ ГЕНОФОНДА ЯКУТОВ ПО ЛИНИЯМ МИТОХОНДРИАЛЬНОЙ ДНК И Y-ХРОМОСОМЫ
Пристальное внимание генетиков к истории якутского этноса было вызвано прежде всего уникальными особенностями генофонда, обнаруженными в 1997 г. английской исследовательницей Татьяной Зерджал [54]. Т. Зерджал впервые показано необычайно высокое содержание гаплогруппы N3 Y-хромосомы (86%) у якутов в небольшой выборке из 21 человека. В дальнейшем исследователями НИИ медицинской генетики (г. Томск) было обнаружено, что к гаплогруппе N3 относится 87% Y-хромосом мужчин из п. Чэриктэй Усть-Алданского улуса Центральной Якутии (n = 46) [18, 19]. Доминирование гаплогруппы N3 для якутского этноса в целом было установлено в работах Б. Пакендорф (Лейпциг, Германия) и соавт. [21, 23] на расширенной выборке якутов из девяти улусов РС(Я) (n = 184). Одновременно с Б. Пакендорф нами был проведен анализ линий Y-хромосомы коренных жителей 16 улусов РС(Я) (n = = 216), который подтвердил высокие частоты гаплогруппы N3 в центральной (86%), вилюйской (93%) и северной (71%) этногеографических группах [5]. В филогенетической сети N3-линий, построенной на основе анализа шести STR-маркеров (DYS19-DYS388-DYS390-DYS391-DYS392-DYS393), выделялся доминантный гаплотип 14-12-23-11-16-14 (Ht1), частота которого в изученной нами выборке составила 64%, что позволило распространить утверждение об эффекте основателя у мужчин п. Чэриктэй Усть-Алданского улуса, впервые высказанное В.А. Степановым в 2002 г., на структуру этноса в целом [5]. Частоты гаплогрупп Y-хромосомы в различных этногеографических группах якутов в сравнении с данными двух других групп ученых приведены в табл. 1 [5, 8, 20, 23]. Помимо доминирующей N3, в мужском генофонде якутов были обнаружены в меньшем количестве и другие азиатские гаплогруппы: N2 (характерна для самодийцев Западной Сибири – нганасан, ненцев, энцев), C3* и С3с (распространены в большей степени среди тунгусоязычных народов) и С3d (обнаружена у народов Южной Сибири – бурятов, телеутов, тувинцев, сойотов, монголов, хамниган). Линии преимущественно европейского происхождения R1a1*, R1a1a7, R1b1b2, J и I1 выявлены у якутов в небольшом количестве: их общая доля составляет от 2 до 11% в различных популяционных выборках [5, 8, 20, 23] (табл. 1).
Таблица 1.
Частоты гаплогрупп Y-хромосомы у центральных (ЦЯ), вилюйских (ВЯ) и северных (СЯ) якутов
| Гаплогруппа | Популяции | |||||||
|---|---|---|---|---|---|---|---|---|
| ЦЯ1 n = 109 |
ЦЯ2 n = 49 |
ЦЯ3 n = 47 |
ЦЯ4 n = 92 |
ВЯ5 n = 56 |
ВЯ6 n = 58 |
СЯ7 n = 32 |
СЯ8 n = 66 |
|
| N3 (ТАТ) | 89 | 93.9 | 93.6 | 85.9 | 98.2 | 93.1 | 87.5 | 71.2 |
| N2 (P43) | 2.8 | 2.0 | 0 | 2.2 | 0 | 0 | 0 | 10.6 |
| N* (DYF155S2) | 0.9 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| C3* (M217) | 1.8 | 2.0 | 2.1 | 0 | 0 | 0 | 3.1 | 3.0 |
| C3c (M48) | 1.8 | 0 | 0 | 0 | 0 | 3.4 | 3.1 | 6.1 |
| C3d (M407) | 0 | 0 | 0 | 1.1 | 0 | 0 | 0 | 4.5 |
| Q (P36) | 0 | 0 | 0 | 0 | 0 | 0 | 3.1 | 0 |
| I1 (M253) | 0 | 0 | 0 | 2.2 | 0 | 1.7 | 0 | 0 |
| J (12f2) | 0 | 0 | 0 | 0 | 1.8 | 0 | 0 | 0 |
| R1a1* (М198) | 3.7 | 2.0 | 4.3 | 4.3 | 0 | 1.7 | 3.1 | 4.5 |
| R1a1a7 (М458) | 0 | 0 | 0 | 2.2 | 0 | 0 | 0 | 0 |
| R1b1b2 (М269) | 0 | 0 | 0 | 2.2 | 0 | 0 | 0 | 0 |
Примечание. 1 Центральные якуты Усть-Алданского улуса (п. Бээди, п. Чэриктэй, п. Дупсээ) [20]; 2 центральные якуты “акающие”; 3 центральные якуты “окающие”, пробы собраны в Намском, Таттинском, Чурапчинском, Мегино-Кангаласском, Алданском улусах [23]; 4 центральные якуты Амгинского, Горного, Кобяйского, Мегино-Кангаласского, Намского, Таттинского, Усть-Алданского, Хангаласского, Чурапчинского улусов [5, 8]; 5 вилюйские якуты Cунтарского, Нюрбинского, Вилюйского улусов [23]; 6 вилюйские якуты Верхневилюйского, Вилюйского, Мирнинского, Нюрбинского, Сунтарского улусов [5, 8]; 7 северные якуты Верхоянского улуса [23]; 8 северные якуты Абыйского, Верхнеколымского, Верхоянского, Жиганского, Момского, Среднеколымского, Томпонского улусов [5, 8]. Гаплогруппы азиатского происхождения выделены серым цветом. Обозначения ветвей гаплогруппы N соответствуют филогенетической номенклатуре, использованной в работе Karmin et al., 2015 [11], которая была далее преобразована в Ilumäe et al., 2016 [12].
Параллельно с изучением линий Y-хромосомы нами были проведены исследования митохондриального генофонда у коренных жителей 25 улусов РС(Я) (n = 191) и впервые обнаружено, что спектр линий мтДНК якутов имеет достаточно высокие показатели генетического разнообразия [1]. Установлено, что более 90% мт-линий якутов относятся к азиатским гаплогруппам A, B, C, D, F, G, M7, M13а, Y, Z, остальные – к европейским кластерам H, HV1, J, T, U, W [1]. Характерной особенностью пула митохондриальной ДНК якутов является преобладание двух гаплогрупп – С и D, широко распространенных в Северной Азии. При этом ~18% якутов из центральной и вилюйской группы улусов являются потомками женщины с линией D5a2а2 [1, 5, 8]. Суммарная частота линий митохондриального генофонда якутов, совпадающих с мт-линиями тюркоязычных народов Средней Азии (казахи, киргизы и уйгуры) и монголов, составила 56%, что указывает на существование общего генетического субстрата, сложившегося до формирования этих этносов [1]. При сравнении с соседними палеоазиатскими народами северо-востока Сибири (чукчи, коряки, ительмены) было установлено, что всего три линии гаплогрупп А8 и С у якутов суммарной частотой 9% пересекаются с линиями митохондриального генофонда коряков. Таким образом, сравнительный анализ показал близкое генетическое родство саха с народами Средней Азии и монголами и дистанцированность от соседних народов Чукотки и Камчатки. При изучении генетических взаимоотношений якутов с народами Средней Азии, Южной Сибири и Камчатки Б. Пакендорф и соавт. пришли к тем же выводам [22, 23]. Частоты гаплогрупп мтДНК в трех этногеографических группах якутов в сравнении с данными двух других групп ученых приведены в табл. 2 [5, 8, 19, 20, 23].
Таблица 2.
Частоты гаплогрупп митохондриальной ДНК у центральных (ЦЯ), вилюйских (ВЯ) и северных (СЯ) якутов
| Гаплогруппа | Популяции | |||||||
|---|---|---|---|---|---|---|---|---|
| ЦЯ1 n = 83 |
ЦЯ2 n = 45 |
ЦЯ3 n = 46 |
ЦЯ4 n = 164 |
ВЯ5 n = 32 |
ВЯ6 n = 111 |
СЯ7 n = 55 |
СЯ8 n = 148 |
|
| A | 1.2 | 2.2 | 2.2 | 2.4 | 7.3 | 3.6 | 0 | 0 |
| B | 3.6 | 4.4 | 4.3 | 0.6 | 3.6 | 0 | 9.4 | 2.7 |
| С | 48.2 | 40 | 37 | 45.7 | 32.7 | 36.0 | 46.9 | 40.5 |
| D | 32.5 | 33.4 | 32.6 | 33.0 | 34.5 | 29.7 | 15.7 | 25.7 |
| F | 3.6 | 4.4 | 8.7 | 3.7 | 3.6 | 5.4 | 15.6 | 4.7 |
| G | 3.6 | 4.4 | 2.2 | 4.9 | 5.5 | 3.6 | 6.3 | 6.8 |
| М | 1.2 | 2.2 | 0 | 1.8 | 1.8 | 2.7 | 0 | 8.2 |
| Y | 0 | 4.4 | 0 | 1.2 | 0 | 2.7 | 0 | 0 |
| Z | 0 | 0 | 2.2 | 0 | 0 | 0 | 0 | 2.7 |
| R1b | 0 | 0 | 2.2 | 0 | 0 | 0 | 0 | 1.4 |
| H | 2.4 | 2.2 | 0 | 3.7 | 3.6 | 6.3 | 3.1 | 1.4 |
| HV1 | 0 | 0 | 0 | 0 | 0 | 1.8 | 0 | 2.0 |
| T | 0 | 0 | 2.2 | 0.6 | 1.8 | 3.6 | 0 | 0 |
| J | 1.2 | 2.2 | 0 | 0.6 | 0 | 2.7 | 0 | 1.4 |
| U | 2.4 | 0 | 4.3 | 0.6 | 0 | 0.9 | 0 | 0.7 |
| W | 0 | 0 | 2.2 | 1.2 | 1.8 | 0.9 | 0 | 2.0 |
| Н.о. | 0 | 0 | 0 | 0 | 3.6 | 0 | 3.1 | 0 |
Примечание. 1 Якуты Усть-Алданского улуса (п. Чэриктэй) [19]; 2 центральные якуты “окающие”; 3 центральные якуты “акающие”, пробы собраны в Намском, Таттинском, Чурапчинском, Мегино-Кангаласском, Алданском улусах [23]; 4 центральные якуты Амгинского, Горного, Кобяйского, Мегино-Кангаласского, Намского, Таттинского, Усть-Алданского, Хангаласского, Чурапчинского улусов [5, 8]; 5 вилюйские якуты Cунтарского, Нюрбинского, Вилюйского улусов [23]; 6 вилюйские якуты Верхневилюйского, Вилюйского, Мирнинского, Нюрбинского, Сунтарского улусов [5, 8]; 7 северные якуты Верхоянского улуса [23]; 8 северные якуты Абыйского, Верхнеколымского, Верхоянского, Жиганского, Момского, Среднеколымского, Томпонского улусов [5, 8]. Гаплогруппы азиатского происхождения выделены серым цветом. Н.о. – гаплогруппы не определены. При обозначении гаплогрупп мтДНК использована номенклатура согласно http://www.phylotree.org/ и Van Oven, Kayser, 2009 [55].
Cубпопуляции якутов отличаются низкой генетической гетерогенностью: хотя северные якуты в большей степени отличаются от центральных и вилюйских, степень межпопуляционных различий между тремя группами саха по Y-хромосоме составляет 4.1%, по мтДНК всего 0.5% [9].
На следующем этапе с целью решения вопроса о соотношении южного и автохтонного компонентов анализ линий мтДНК был проведен на расширенных выборках якутов (n = 423) в сравнении с соседними этносами (эвенками, эвенами и юкагирами) [5, 8]. По спектру гаплогрупп мтДНК якуты оказались более близки к эвенкам и отдалены от юкагиров и эвенов [5, 8]. С помощью более глубокого филогенетического анализа было установлено, что в отличие от автохтонов более характерными для якутов являются азиатские субкластеры С4а1с, С4а2, С5b1b, D5а2а2, G2a5, Z3, а также бόльшее разнообразие линий европейского и ближневосточного происхождения [8]. Доля типичных для якутов субкластеров мтДНК (D5а2а2, С4а1с, С4а2, С5b1b, G2a, Z3, H20a, HV1a1a, U4d2) у центральных и вилюйских якутов составляет ~48% [8], у северных якутов, генетически более близких к автохтонам, ~38%.
К линиям, более характерным для локальных популяций, относятся C4b3a, C4b7, C5d1, Z1a, распространенные у юкагиров и эвенов, а также “корякские” A8, C4b2, C5a2a, G1b и “эвенкийские” C4b9. К автохтонным также, очевидно, относятся линии R1b, D4b1c, A4b, C4b*, выявленные у семи индивидов из древних погребений Якутии, датируемых временем от 16.9 до 3.1 тыс. лет назад: Хайыргас, Онньес, Кердюген, Каменка, Помазкино [4, 56]. Все эти линии были, по-видимому, привнесены в митохондриальный генофонд якутского этноса в результате браков мужчин-саха с женщинами из соседних племен на территории Якутии.
Таким образом, в генетических исследованиях было установлено, что отличительной особенностью якутской популяции являются низкое разнообразие линий Y-хромосомы и высокое разнообразие линий митохондриальной ДНК. Как в мужском, так и в женском генофонде якутов наблюдаются эффекты основателя. Сравнительный анализ линий мтДНК якутов с линиями южных народов (казахи, киргизы, уйгуры, монголы) и территориально соседних этносов (эвенки, эвены, юкагиры, чукчи, коряки, ительмены) позволил нам предположить, что около половины мт-линий прибыли на современную территорию проживания народа саха вместе с южными предками, а другая половина имеет автохтонное происхождение.
Низкий уровень разнообразия линий Y-хромосомы, как правило, наблюдается у народов, имеющих небольшую численность [57]. Высокая частота одной N3-гаплогруппы (в среднем 83%) и, более того, одного N3-гаплотипа (Ht1, 64%) в мужском генофонде якутов – необычное явление для достаточно многочисленного этноса. По данным Всероссийской переписи 2010 г. численность якутов в РФ составляет 478 085 человек [58]. К генетическим процессам, которые могут определять высокие частоты отдельных гаплотипов Y‑хромосомы и мтДНК в генофонде этноса, помимо эффекта основателя также относят эффект бутылочного горлышка, который подразумевает значительное снижение численности популяции из-за неблагоприятных условий (эпидемии, войны, голод) с уменьшением уровня генетического разнообразия и преимущественного “выживания” некоторых гаплотипов. Многочисленные эпидемии и войны в истории народа саха [38, 59], таким образом, могли даже способствовать увеличению доли N3-хромосом в генофонде популяции за счет эффекта генетического дрейфа в том случае, если в исходной малочисленной предковой популяции доминировали мужчины с гаплогруппой N311.
Кроме этих факторов, не исключено также влияние естественного отбора отдельных линий Y-хромосомы, обусловливающих генетическое преимущество для выживания мужчин в условиях холодного климата. В пользу данного предположения свидетельствует преимущественное распространение носителей гаплогруппы N3 в северных широтах Евразийского континента [3]. Однако принимая во внимание то, что в других северных популяциях, соседних с якутами (эвенки, эвены, юкагиры, долганы), частота гаплогруппы N3 не достигает столь высоких значений (26–33%) [5, 8], мы считаем, что этот фактор не мог быть определяющим.
По нашему мнению, на формирование особенностей структуры современного генофонда якутов в большей степени повлияли брачные традиции народа саха: патрилинейность якутских кланов, строгая экзогамия (выбор жены из другого рода, из отдаленного региона) и ограничения в притоке линий в мужской генофонд популяции от соседних этносов. В этнографических исследованиях отмечалось, что мужчины-якуты часто и охотно женились на тунгусках, в то время как женщины-саха редко выходили замуж за тунгусов [61]. Эта же тенденция сохраняется по настоящее время: в исследованиях А.Н. Кучер и А.Л. Даниловой при анализе структуры браков в трех улусах со смешанным национальным составом было отмечено, что мужчины-якуты чаще вступают в межнациональные браки с представителями других коренных национальностей, чем женщины-якутки [62, 63]. Проведенный нами сравнительный анализ показал, что с тунгусоязычными эвенками и эвенами совпадают от 51% мт-линий у центральных якутов до 66% у северных [5], тогда как в мужском генофонде саха наблюдается низкое содержание линий, характерных для тунгусов (С* и С3с), – от 2% у центральных якутов до 9% у северных [5, 8]. Поток генов от тунгусов к якутам, по-видимому, происходил главным образом через заключение смешанных браков мужчин-саха с женщинами-тунгусками, тогда как в направлении от якутов к тунгусам перенос генов через женщин был незначителен из-за строгих национальных брачных традиций, ограничивающих браки женщин-саха с мужчинами из других этносов [61, 62].
Низкая частота линий Y-хромосомы западноевразийского происхождения в мужском генофонде якутов также указывает на незначительный процент браков между женщинами-саха и европейцами. Общее содержание линий Y-хромосомы, имеющих недавнее, в рамках эволюционного времени, происхождение вследствие метисации с европейцами (I1, R1b1b2, R1a1a7), в изученной нами выборке составляет всего 3%. В генетико-демографических исследованиях населения административных центров пяти улусов РС(Я), проведенных А.Н. Кучер и соавт., отмечается, что по настоящее время межнациональные браки в большинстве случаев заключаются между представителями коренных национальностей Республики Саха (Якутия) – якутами, эвенами и эвенками, тогда как браки между пришлым и коренным населением не нашли широкого распространения [63, 64].
Следует также отметить значительный разрыв в показателях смертности мужчин и женщин-саха, который, по-видимому, был характерен для населения Якутии до начала XX в. По данным переписи 1926 г. смертность женщин в возрастных группах от 15–50 лет была в 1.5–2 раза выше, чем у мужчин, в возрасте от 20–29 лет – более чем в 2 раза [65] соответственно было больше мужчин, которые женились в течение жизни несколько раз. Возможно, этот фактор в какой-то степени внес вклад в значимые различия по разнообразию линий Y-хромосомы и мтДНК в генофонде народа саха.
Помимо вышеперечисленных, нельзя также исключить вклад такого фактора, как многоженство, хотя этнографами отмечалось, что “…Фактически многоженство было доступно очень немногим и никогда не могло стать преобладающей у якутов формой брака” вследствие высоты калыма [61]. На основании изучения архивных материалов установлено, что в XIX–начале XX в. у якутов преобладала моногамная форма семьи. Для внутреннего строя якутской семьи было характерно господство патриархальных традиций, регулировавших весь уклад семейной жизни [66].
ПРОИСХОЖДЕНИЕ СПЕЦИФИЧЕСКИХ N3-ЛИНИЙ (М46, ТАТ) Y-ХРОМОСОМЫ ЯКУТОВ
Происхождение специфических N3-линий Y‑хромосомы якутов является наиболее интригующим вопросом в изучении генетической истории народа саха. На начальном этапе решение данного вопроса было крайне затруднительно, так как гаплогруппа N3 встречается по всей территории Северной Евразии, в том числе во всех популяциях Якутии [3, 5, 57, 67–69], в Средней Азии [57, 70], Скандинавии, Северном Китае, Японии [3] и на Кавказе [71, 72].
В ранних исследованиях изучение происхождения N3-линий якутов проводилось главным образом путем анализа филогенетических сетей микросателлитных STR-гаплотипов. К недостаткам данного подхода относится то, что структура филогенетической сети может сильно варьировать в зависимости от использованных для построения сети популяционных выборок и наборов STR-маркеров, что затрудняет определение наиболее близких предковых гаплотипов по отношению к “якутским”. Определение времени происхождения наименее древнего общего предка (TMRCA – time of the most recent common ancestor), в свою очередь, зависит от правильного выбора предкового гаплотипа и метода определения скорости возникновения мутаций в использованных STR-локусах. Структура сети также усложняется гомоплазией и обратными мутациями, однако этот подход до сих пор является актуальным в силу своей относительной простоты и дешевизны, а также широкого использования STR-наборов в судебной медицине.
На основании сравнения STR-линий якутов с восточными эвенками, бурятами и тувинцами учеными НИИ медицинской генетики (г. Томск) было сделано заключение, что N3-линии якутов наиболее близки к таковым у восточных эвенков [18, 19]. Авторы предположили, что мужской генофонд якутов сложился на базе автохтонного компонента, а тюркский язык был приобретен в результате культурного доминирования тюркоязычной элиты, не оставившей значительного следа в пуле Y-хромосом. Позднее, на основе более глубокого филогенетического анализа N3-линий из 18 популяций Северной и Восточной Европы, Чукотки, Западной и Южной Сибири, и оценки времени генерации N3-линий в якутской популяции в 4.45 ± 1.96 тыс. лет В.Н. Харьковым и др. было высказано мнение, что эти линии были приобретены якутами опосредованно через эвенков от еще более древних автохтонных племен (возможно, протоюкагиров или других палеоазиатских племен), проживавших ранее на территории современной Якутии и ассимилированных тунгусами [20]. Необычайно высокую частоту N3-линий в мужском генофонде якутов (89%), в отличие от эвенков (15–40%), авторы объяснили значительным увеличением численности якутов за последние века, которое привело к случайному резкому увеличению частот отдельных линий с преобладанием гаплотипа-основателя в популяции. Таким образом, при интерпретации полученных данных исследователи Института медицинской генетики (г. Томск) склонились к версии автохтонного происхождения N3-линий якутов и незначительности вклада южного тюркского компонента в генофонд современных якутов.
В работе Б. Пакендорф и соавт. было установлено, что в филогенетической сети STR-гаплотипов русских, латвийцев, финнов, саами, хантов, манси, карелов, марийцев, бурятов, юкагиров, западных эвенков и тувинцев N3-линии якутов образуют отдельную ветвь и перекрываются только с линиями эвенков [21, 23]. Время генерации микросателлитного разнообразия N3-гаплотипов якутов по оценке Б. Пакендорф и соавт. – 880 ± ± 440 лет, что по утверждению авторов, соответствовало гипотезе о вытеснении якутов на север в XII в. из региона оз. Байкал под давлением монголов. С точки зрения Б. Пакендорф и соавт. полученные результаты свидетельствовали о южном происхождении якутского этноса и незначительном смешении пришлых тюркоязычных племен с автохтонными [23]. Авторы отметили загадочность происхождения N3-линий якутов, не похожих на N3-линии других исследованных ими популяций за исключением эвенков.
В наших исследованиях N3-линии якутов были рассмотрены в сравнении с 27 популяциями Южной и Западной Сибири, Северной и Восточной Европы, Чукотки и Камчатки [8]. В структуре расширенной филогенетической сети N3-линии якутов формировали отдельную ветвь, что соответствовало данным других авторов, но результаты филогенетического анализа получили другую интерпретацию. Структура “якутской” N3-ветви, в которой доминировали два гаплотипа (более древний Ht2 и более “молодой” Ht1, отличающийся от первого мутацией по маркеру DYS392), по нашему мнению, свидетельствовала о двух последовательных экспансиях численности якутской популяции (рис. 1). Дивергенция более древнего гаплотипа Ht2, рассчитанная нами по скорости мутирования STR-маркеров, началась ~1600 лет назад, а гаплотипа Ht1 ~900 лет назад [8]. При этом если в IV в. в предковом племени, очевидно, доминировали мужчины с гаплотипом Ht2, то в популяции якутов, увеличение численности которой началось ~900 лет назад, существовали носители обоих доминантных гаплотипов (Ht1 и Ht2). В отличие от В.Н. Харькова и соавт. [20] мы считаем более вероятным возникновение “якутской” N3-ветви в Восточно-Саянском регионе, чем на территории современной Якутии, так как близкие к якутским STR-гаплотипы присутствуют у народов, проживающих к западу от Байкала, – тувинцев, тофаларов и сойотов.
Рис. 1.
Филогенетическая сеть N3-гаплотипов Y-хромосомы народов Якутии и соседних регионов на основе 5 STR-маркеров [8]. Кругами обозначены микросателлитные гаплотипы, площадь круга и секторов внутри круга пропорциональна частоте гаплотипа. Доминантные “якутские” гаплотипы Ht1 и Ht2 обозначены стрелками. Черной стрелкой указан корневой гаплотип (Root).
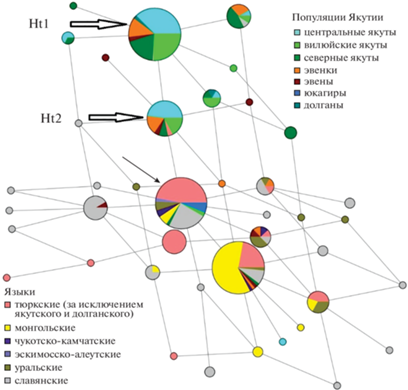
N3-линии якутов действительно перекрываются с N3-линиями эвенков, что было показано в параллельных работах двух других групп исследователей [18–20, 23], и кроме того с N3-линиями тунгусоязычных эвенов и тюркоязычных долган [8], но основываясь на данных этнографов и на изучении брачных традиций, мы не можем согласиться с утверждением, что “мужской генофонд якутов сложился на базе локального (эвенкийского) компонента” [18–20] и считаем более вероятным обратный процесс – перенос генов от якутов к эвенкам, эвенам и долганам.
Более поздние исследования позволили в значительной степени уточнить структуру филогенетического дерева Y-хромосомы [11, 12] и в какой-то мере прояснить вопрос о происхождении N3-линий якутов. На основе анализа полных последовательностей Y-хромосом у 94 индивидов из различных популяций Евразии в гаплогруппе N3 было выделено три ветви, каждая из которых имеет специфическое территориальное распределение: N3a имеет наиболее широкое распространение в Северной Евразии – от Скандинавии до Чукотки, N3b – в Южной Сибири и Монголии, N3c – в Японии, Китае и других регионах Восточной Азии. В свою очередь N3a разделяется на шесть субгаплогрупп (N3a1–N3a6), возникших в период ~3.3–8.3 тыс. лет назад. Субгаплогруппа N3a1 встречается у народов Волго-Уральского региона, в Южной и Западной Сибири, N3a2 – в Якутии, Южной и Западной Сибири, Средней Азии и на Кавказе, N3a3 и N3a4 – в Северо-Восточной Европе, Волго-Уральском регионе и Западной Сибири, N3a5 – на Чукотке, Камчатке, в Южной Сибири, Монголии, Средней Азии и на Кавказе, N3a6 – на Дальнем Востоке (у нанайцев).
Филогеографический анализ структуры и распространения ветвей гаплогруппы N3 показал, что линии якутов относятся к субгаплогруппе N3a2 и не являются эволюционными производными от линий, доминирующих у монголов и бурятов (N3a5-F4205), чукчей, эскимосов и коряков (N3a5-B202), нанайцев (N3a6), японцев (N3c), а также отличны от N3-линий народов Волго-Уральского региона, Западной Сибири, Восточной Европы и Скандинавии (N3a1, N3a3 и N3a4).
Субгаплогруппа N3a2, в состав которой входят линии якутов, отделилась от кластера N3a2'6 ~7.1 тыс. лет назад [11, 12]. Самая древняя ее ветвь N3a2-B508, возраст которой ~4.5 тыс. лет, включает линии индивидов из Ливанской Республики и Китая. N3a2-линии были обнаружены также у отдельных индивидов из Турции и Королевства Бутан, расположенного между Индией и КНР [73]. Единичные N3a2-линии были выявлены в популяциях сибирских татар, тувинцев, шорцев, монголов, казахов, узбеков, караногайцев, каракалпаков, хантов и манси [12]. Таким образом, эта субгаплогруппа, возникшая в эпоху неолита, имеет очень широкий ареал распространения, включает представителей народов разных языковых семей и помимо Сибири, Средней Азии и Кавказа встречается также на Ближнем Востоке и в Южной Азии.
“Якутская” ветвь субгаплогруппы N3a2 определяется маркерами М1982, M1993, M1987, M2032, M2077, M2103, M2108, M2011, M2122 и имеет относительно небольшой возраст. Время дивергенции этой ветви, обозначенной как N3a2-M1982, по данным полного секвенирования Y-хромосом четырех мужчин составило ~1.7 ± 0.8 тыс. лет [11, 12], что вполне соответствовало нашим расчетам, проведенным до этого на основе анализа STR-локусов (~1.6 ± 0.6 тыс. лет) [5, 8]. По последним данным, доступным в настоящее время из базы https://www.yfull.com/tree/, “якутская” N3a2-1982-ветвь сформировалась ~2300 лет назад (95% ДИ 3000–1500 лет назад). При этом время возникновения наименее древнего общего предка, рассчитанное на основе полного секвенирования Y-хромосом 20 мужчин-саха, составляет ~1.4 тыс. лет (95% ДИ 2100–850 лет назад), что в пределах доверительного интервала соответствует полученным нами ранее значениям. Эти генетические датировки, очевидно, будут скорректированы в ближайшем будущем с увеличением количества полных сиквенсов N3-M1982-хромосом, что позволит уменьшить границы доверительного интервала и определить TMRCA с большей точностью. В период увеличения численности якутской популяции (~1.4 тыс. лет назад) N3а2-М1982-ветвь делится на две крупные подветви, определяемые мутациями M1932 и M1991. Судя по комбинациям STR у мужчин-саха, опубликованным в базе данных https:// www.yfull.com/tree/, более древний гаплотип Ht2 соотносится с подветвью M1932, а гаплотип Ht1 – c подветвью M1991. Дальнейшая дивергенция этих подветвей происходит начиная с XI в., при этом значимое вторичное увеличение численности популяции якутов с наибольшей вероятностью приходится на период 450–500 лет назад.
Таким образом, время дивергенции N3-линий якутов, рассчитанное на основе микросателлитного разнообразия STR-гаплотипов и по доступным в настоящее время данным полного секвенирования N3а2-1982-хромосом, указывает на IV–VII вв. как наиболее вероятный период первичной экспансии предковой популяции, за которым последовала вторичная экспансия в период с XI по XX в. с наиболее усиленным ростом численности якутов начиная с XVI в.
Любопытно то, что дата первой экспансии численности популяции (IV–VII вв.) соответствует времени расхождения якутского языка c древними тюркскими языками (в IV–V вв.) по Г.Г. Левину [32], А. Савельеву и М. Робитс [47] и археологическим данным, указывающим на появление скотоводческих групп в Южной Якутии в III–IV вв. [38]. Вторая дата (~900 лет назад) совпадает с предполагаемым временем миграции последней, наиболее обширной волны тюркоязычных предков якутов в бассейн Средней Лены по Г.В. Ксенофонтову [28]. Основываясь на времени дивергенции гаплотипов Ht2 и Ht1, связанных с последовательными увеличениями численности популяции, нами было выдвинуто предположение, что южные предки якутов могли перекочевать на Север несколькими волнами, первая из которых произошла в период распада империи хуннов, а вторая – чуть раньше создания империи Чингисхана [14]. Косвенным подтверждением раннего обособления и переселения южных предков якутов на Север до возникновения Монгольской империи может служить тот факт, что у якутов не обнаружена так называемая “С3-линия чингизидов”, распространенная на территориях, находившихся под властью Чингисхана [8]. Следует также отметить, что по глоттохронологическим датировкам язык предков саха сформировался примерно 1500–1800 лет назад [32, 45, 47], а все сохранившееся разнообразие монгольской группы языков сложилось всего около 1000 лет назад [46] и соответственно язык саха намного древнее любого из известных современных монгольских языков. Присутствие монгольского компонента в якутском языке, таким образом, может объясняться не столько поздними заимствованиями, сколько намного более древним его происхождением.
ПРОИСХОЖДЕНИЕ ЗАПАДНОЕВРАЗИЙСКИХ ЛИНИЙ В ГЕНОФОНДЕ ЯКУТОВ
Проведенный нами анализ спектра линий Y‑хромосомы показал, что западноевразийский компонент генофонда якутов представлен гаплогруппами I1, R1a1*, R1a1a7, R1b1b2 [5, 8] (табл. 1). Кроме них, единичные линии гаплогруппы J были обнаружены у вилюйских якутов в исследованиях Б. Пакендорф и соавт. [23]. Общее содержание линий Y-хромосомы западноевразийского происхождения для объединенной выборки численностью 509 мужчин составляет в среднем 7% (табл. 1) [5, 8, 20, 23].
Гаплогруппа I1 предположительно возникла в Северо-Западной Европе, распространилась с максимальной частотой в странах Скандинавии и далее проникла на территории восточных славян [74]. Гаплогруппа R1b1b2 более характерна для Западной Европы и Ближнего Востока [75]. Учитывая тот факт, что у славян гаплогруппы I1 и R1b1b2 встречаются повсеместно с умеренными частотами [69, 74, 76], можно утверждать, что происхождение этих линий в генофонде якутов связано с процессами метисации с русскими и, возможно, другими восточноевропейскими этносами (украинцы, белорусы, татары), заселившими территорию Якутии начиная со времени вхождения в состав Российской империи в XVII в.
Гаплогруппа R1a1a7 распространена главным образом в популяциях Центральной, Восточной и Северной Европы [69, 77, 78]. Анализ разнообразия микросателлитов гаплогруппы R1a1a7 показал наибольшее значение в популяции поляков, что предполагает возникновение данной гаплогруппы на территории современной Польши [77]. Линии гаплогруппы R1a1a7 обнаружены нами у двух центральных и одного вилюйского якута. Происхождение этих линий в генофонде якутов, возможно, связано с событиями переселения в Якутию ссыльных поляков [79].
Что касается линии R1а1*, которая встречается с небольшими частотами во всех изученных субпопуляциях якутов, и гаплогруппы J, обнаруженной помимо якутов у долган и эвенов, предполагается более древнее происхождение этих линий из южных регионов Алтае-Саянского нагорья. Линии R1а1* Y-хромосомы широко распространены в популяциях Южной Сибири и Монголии – у алтайцев, хакасов, шорцев, сойотов, тувинцев, тофаларов, бурятов, уйгуров, монголов [57, 67–69], гаплогруппа J встречается у алтайцев-кижи, телеутов, уйгуров, монголов, казахов, узбеков, тувинцев [57, 67–69]. Присутствие этих линий в генофонде народов Южной Сибири и Монголии объясняется существованием древнего потока генов с запада на восток по степному поясу Евразии. Анализ ДНК древних популяций указывает на то, что в бронзовом и железном веках европеоиды проживали в Южной Сибири, в том числе на Алтае [80–82], а индоевропейская миграция достигала даже Северо-Восточной Монголии [83].
Таким образом, содержание западноевразийских линий Y-хромосомы, имеющих недавнее, в рамках эволюционного времени, происхождение со времени вхождения территории Якутии в состав Российской империи в XVII в. (I1, R1b1b2, R1a1a7), в изученной нами выборке составляет всего 3%. Доля линий Y-хромосомы древнего европейского происхождения (R1a1*, J), очевидно мигрировавших из Южной Сибири вместе с южными предками якутов, в среднем ~4%.
Анализ мтДНК показал, что западноевразийский компонент митохондриального генофонда якутов представлен единичными линиями гаплогрупп Н*, H1*, H1a, H20a, U4d2, U5b1b1a, HV1a1a, J1c5, T2, W суммарным содержанием 9% от общей выборки (n = 423) [5, 8]. Источником большинства этих митохондриальных линий (Н*, H1*, H1a, U5b1b1a, T2, W) были, по-видимому, русские и, возможно, другие восточноевропейские этносы, так как они распространены у восточных славян и народов Волго-Уральского региона [84–87]. Суммарная частота этих линий у якутов составляет 6%. Из исторических данных известно, что численность женщин русской национальности в Якутии в XVII–XVIII вв. была крайне низкой [88]. Русское население в Якутии в XVII в. составляли главным образом мужчины, которые брали в жены местных женщин. Только с конца XVIII в. началось более массовое переселение в Якутскую губернию семей крестьян-земледельцев и почтовых работников, обслуживающих ямщицкие тракты [89]. Численность русского населения росла до XX в. очень медленно: по данным Первой всеобщей переписи Российской империи 1897 г., русские Якутской области составляли 30 807 человек, или 11.4% от общей численности населения [90].
Меньшая часть линий митохондриального генофонда якутов (H20a, U4d2, HV1a1a, J1c5) имеет, по-видимому, более древнее происхождение, не связанное с переселением русских женщин на территорию Якутии начиная с XVIII в. Гаплогруппа J мтДНК была обнаружена в останках из якутского погребения начала XVII в. [91], задолго до миграций семей русских крестьян в Якутию в XVIII в. [89]. Происхождение линий H20a и HV1a1a у якутов также предполагает поток генов не от русских, потому что эти гаплогруппы редки (H20a) или даже отсутствуют (HV1a1a) у них [84, 85, 92–94], но распространены у южносибирских народов, а также на Кавказе, Ближнем и Среднем Востоке [94–97]. Кроме того, ГВСI-линии H20a и HV1a1a якутов в точности совпадают с гаплотипами бурят из Республики Бурятия [95]. Линия U4d2, обнаруженная нами у одного вилюйского якута, возможно, имеет западносибирское происхождение, так как U4d2-линии в большей степени характерны для народов Западной Сибири и данная ГВСI-линия (16189–16356) выявлена у нганасан и манси [98, 99]. Соответственно, общая доля митохондриальных линий древнего западноевразийского происхождения (H20a, U4d2, HV1a1a, J1c5) в генофонде якутов составляет всего ~3%.
СТРУКТУРА ГЕНОМА ЯКУТОВ ПО ДАННЫМ ПОЛНОГЕНОМНОГО АНАЛИЗА
Полногеномный анализ более чем 600 тыс. SNPs у 25 якутов в числе 938 человек из различных этнических групп Европы, Азии, Африки и Америки впервые был проведен в работе J.Z. Li и соавт. (2008) [100]. Результаты первого полногеномного исследования показали преобладание азиатского (>90%) и низкое содержание европейского компонента (<10%) в геноме якутов, что полностью коррелировало с данными, полученными ранее при изучении спектра линий Y-хромосомы и мтДНК [1, 5, 18–20, 23].
В 2010 г. в исследованиях генома древнего палеоэскимоса из Гренландии возрастом ~4000 лет с целью поиска генетически близких к нему индивидов из современных популяций Евразии был осуществлен анализ структуры генома других народов Якутии – эвенков, долган и юкагиров [6]. При этом было отмечено генетическое сходство саха с эвенками и глубокие отличия якутов от народов крайнего Северо-Востока Евразии (чукчей и коряков), что также коррелировало с результатами исследовательских работ, осуществленных ранее с применением гаплоидных генетических систем [1, 5, 23].
Далее для более детального изучения генетической истории заселения северо-востока Евразии нами был проведен полногеномный анализ 758 человек из 55 популяций [8]. Помимо 40 новых образцов из популяций Якутии, Южной и Западной Сибири, в выборку были включены опубликованные к тому времени данные по популяциям Европы, Азии и Америки из работы D. Li et al. (2008) [100]. При изучении генетических взаимоотношений между народами Якутии и соседних регионов были использованы PC-анализ и алгоритм ADMIXTURE, который дает количественные оценки различных генетических компонентов при определении происхождения от предполагаемого числа К предковых популяций. Установлено, что главной особенностью генома якутов является доминирование центрально-сибирского компонента, объединяющего их, в первую очередь, с долганами Таймыра и эвенками (рис. 2). Другие сибирские генетические компоненты, ярко представленные у народов арктической зоны Якутии и Красноярского края – эвенов, юкагиров, нганасан, кетов и селькупов, едва присутствуют у якутов. В геноме якутов также почти не выражен “палеоазиатский” компонент, общий с коряками и чукчами. Результаты исследования показали тесное генетическое родство якутов с долганами и эвенками Якутии, меньшее сходство с эвенами, юкагирами и нганасанами, отдаленность от кетов и селькупов Западной Сибири и глубокие генетические отличия от соседних палеоазиатских народов Камчатки и Чукотки (рис. 2) [8].
Рис. 2.
Соотношение генетических компонентов у 758 индивидов из 55 популяций с использованием программы ADMIXTURE при предполагаемом числе предковых популяций K = 3, K = 4, K = 6, K = 8, K = 13 [8].

Генетическая близость тюркоязычных якутов и долган неудивительна: долганы – один из самых молодых этносов Сибири, который сформировался в период летописной истории в течение XVII–XIX вв., вобрав линии якутов, эвенков и русских старожилов, а также энцев и ненцев [101]. К приходу русских в Восточную Сибирь такой народности не существовало: в XVII в. по Лене, Вилюю и побережью Охотского моря были встречены лишь отдельные тунгусские роды с названием Долган. Но к концу XIX в. по данным переписи 1897 г. численность этноса составляла 2633 человек [101]. Долганский язык является древним языком якутов в период их продвижения на север и активного взаимодействия с тунгусами, поэтому долгое время рассматривался как один из диалектов якутского [101].
Генетическое сходство якутов с эвенками, по-видимому, объясняется не только процессами смешения территориально близких этносов в Якутии в течение длительного периода времени, но и происхождением из общего южносибирского генетического субстрата (причем если происхождение народа саха по генетическим данным связано с регионами западнее Байкала, то тунгусов – с территориями восточнее Байкала). Точка зрения о местонахождении прародины тунгусов к востоку от Байкала поддерживается многими учеными, в том числе археологическими, лингвистическими и генетическими исследованиями [27, 102–104]. Анализируя материалы археологических памятников Забайкалья, Приамурья и особенно среднего Амура (ныне район Благовещенска), а также Приморья, археологи пришли к выводу, что характерные для тунгусской культуры таежные черты сложились в этом регионе на границе тайги и степи [102]. Анализ “диагностической” лексики названий лиственных деревьев (в частности, ясеня и дуба), лососевых рыб, потамонимов (названия рек) и домашних животных, проведенный лингвистом А.М. Певновым, позволил предположить, что “родина языковых предков тунгусо-маньчжурских народов находилась не в равнинных районах бассейна Среднего Амура, а в гористой местности с хвойно-широколиственными лесами” (возможно, южная часть Буреинского хребта и Малый Хинган, параллельно р. Амур) [103]. Результаты генетических исследований коррелируют с данными археологов и лингвистов: анализ структуры генома эвенков Таймыра и Подкаменной Тунгуски, а также эвенов из других этногеографических групп Якутии и Камчатки [27] свидетельствует в пользу гипотезы о местонахождении прародины тунгусов в бассейне р. Амур. С помощью комплексного подхода, объединяющего лингвистические, археологические и генетические данные, было установлено, что прародина тунгусоязычных этносов, вероятнее всего, находится на правобережье нижнего течения р. Амур [104].
Любопытно, что у эвенков Якутии нами был обнаружен “гаплотип Чингисхана” [8], встречающийся иногда у народов, бывших под властью чингизидов [105], что указывает на относительно недавний поток генов через мужчин от монголов к эвенкам. Полученные генетические аргументы свидетельствуют в пользу гипотезы о сравнительно недавнем (в начале II тыс. н. э.) появлении тунгусов на территории Якутии, поддерживаемой А.Н. Алексеевым [36]. Масштабные экспансии тунгусов Забайкалья и тюркоязычных якутов Прибайкалья по территории Восточной Сибири в пределах последних 2000 лет, по-видимому, привели к ассимиляции и/или вытеснению древнего населения на еще более северные территории. Поэтому неудивительно, что по сравнению с эвенками Якутии народы Арктического побережья (юкагиры, эвены и нганасаны) в большей степени отличаются от саха по составу компонентов генофонда и содержанию специфических линий мтДНК и Y-хромосомы [5, 8].
Из современных народов, проживающих в соседних с Якутией регионах, по степени генетической близости к якутам оказались более близки буряты и тувинцы, а также алтайцы, монголы и хакасы [8]. Полногеномные исследования, проведенные позднее на расширенных выборках из популяций Сибири, подтвердили полученные нами данные [27, 106, 107]. При сопоставлении якутов с алтайцами, тувинцами и бурятами с помощью IBD-анализа было показано более близкое генетическое сходство якутов с бурятами [27]. С другой стороны, согласно данным M. Robbeets et al. (2021) по генетическим дистанциям к якутам более близки тюркоязычные тувинцы, чем монголоязычные буряты [104]. В отличие от вышеперечисленных народов Южной Сибири геном якутов более однороден и “обеднен” по составу компонентов, что еще раз подтверждает малочисленность предковой популяции, отделившейся от генетического субстрата, общего с южными тюркоязычными народами [8].
ГЕОГРАФИЧЕСКОЕ РАСПОЛОЖЕНИЕ ПРАРОДИНЫ ЯКУТОВ
К настоящему времени в результате исследований древних популяций Южной Сибири и Восточной Азии получены новые сведения, касающиеся географического расположения прародины саха. В недавних исследованиях E.H.M. Wong и соавт. (2017) [108], P.B. Damgaard и соавт. (2018) [109], С. Jeong и соавт. (2020) [110], G.M. Kılınç и соавт. (2021) [56] и M. Robbeets и соавт. (2021) [104] были проанализированы геномы более чем 300 индивидов из древних погребений Южной Сибири, Монголии, Китая, Кореи и Японии, датированных в широком интервале времени – от неолита до средневековья. По полногеномным данным к современным саха оказались более близки древние индивиды глазковской культуры бронзового века из могильника Шаманка II, расположенного на южной оконечности о. Байкал (Иркутская область), а также из некоторых погребений хунну железного века, локализованных в Бурятии и Монголии, и монголов средневековья [56, 104, 109, 110]. Эти данные полностью коррелируют с результатами проведенных ранее краниологических исследований, сближающих якутов с глазковцами Приангарья и древними хунну [42].
Но при этом следует отметить, что изученные к настоящему времени древние популяции хунну отличаются от якутов по спектру линий Y-хромосомы преобладанием гаплогрупп R1a и Q [110, 111]. Частота N3-линий у древних хунну из 29 погребений, дисперсно расположенных на территории Монголии, – 3% (n = 29) [110], из могильника Тамирын-Улан-Хошу Центральной Монголии – 8% (n = 25) [111]. Сравнительный анализ N3-линий хунну с другими древними популяциями показал их сходство с STR-гаплотипами древних аварцев из нескольких погребений VII–VIII вв. на территории Венгрии и одного якута из погребения Сергеляхское шоссе (XV–XVI вв.). Однако по комбинации STR-маркеров N3-линии древних хунну и аварцев более близки между собой и отличаются от N3-линии cредневекового конного воина-саха [111]. Стоит также подчеркнуть, что N3-линия индивида из погребения Сергеляхское шоссе ближе к корневому гаплотипу и не является филогенетически близкой к гаплотипам Ht1 и Ht2, доминирующим в якутской популяции [8, 16]. Соответственно, утверждение о прямой связи саха с хунну по отцовским линиям является преждевременным и скорее всего ошибочным.
Линии Y-хромосом охотников и рыболовов бронзового века из погребения Шаманка II, останки которых датированы периодом 4000–3800 лет назад, относятся к гаплогруппе Q [109]. Несоответствие данных по Y-хромосоме с результатами полногеномного анализа, указывающими на генетическое сходство якутов с индивидами глазковской культуры, возможно определяется присутствием автохтонного компонента, содержание которого в геноме современных саха, по нашему предположению, составляет около 50%. К тому же следует учесть, что в палеогенетических исследованиях до сих пор не были проанализированы останки древних индивидов из могильников курумчинской и усть-талькинской археологических культур, непосредственно связываемых с южными предками народа саха, и оценка степени генетической близости современных якутов с древними глазковцами и хунну является предварительной из-за ограниченного числа исследованных археологических культур Южной Сибири. Расширенный ДНК-анализ древних популяций Прибайкалья и Алтае-Саянского нагорья позволит в будущем более точно локализовать географическое расположение прародины саха. Результаты полногеномных исследований современных и древних популяций Сибири полностью согласуются с идеей, что предки якутов жили когда-то в непосредственной близости от предков бурят и тувинцев на берегах оз. Байкал.
ЗАКЛЮЧЕНИЕ
Согласно генетическим реконструкциям, в предковой популяции якутов доминировали мужчины с гаплогруппой N3а2-М1982, преимущественно одного рода (Ht1), и существовало достаточно высокое разнообразие линий женщин, среди которых преобладал род с гаплогруппой D5a2a2. На формирование особенностей структуры современного генофонда якутов, очевидно, повлияли брачные традиции народа саха – экзогамия (выбор супруги из другого рода и/или отдаленного региона), браки с женщинами из соседних автохтонных племен и ограниченность притока линий от соседних народов в мужской генофонд популяции. Сравнительный анализ линий мтДНК якутов с линиями южных народов (казахи, киргизы, уйгуры, монголы) и территориально соседних этносов (эвенки, эвены, юкагиры, чукчи, коряки, ительмены) позволяет предположить, что около половины мт-линий прибыли на современную территорию проживания народа саха вместе с южными предками, а другая половина имеет автохтонное происхождение. В отличие от других групп генетиков, преувеличивающих долю автохтонного [18, 19] или южного тюркоязычного [23] компонента, мы считаем, что их соотношение в генофонде центральных и вилюйских якутов примерно равное. У северных якутов доля автохтонного компонента несколько повышена (до ~60% по линиям мтДНК).
Содержание европейского компонента в генофонде якутов по данным всех трех генетических систем (Y-хромосома, мтДНК, полногеномный анализ SNP-маркеров) ≤10%. Генетические исследования доказали двойственное происхождение этого компонента [5, 8]: часть линий была привнесена в генофонд якутов относительно недавно со времени вхождения Якутии в состав Российской империи, 3–4% линий имеют более древнее происхождение, не связанное с миграцией русскоязычного населения в Сибирь начиная с XVII в. Происхождение древних западноевразийских линий в генофонде якутского этноса, по-видимому, связано с доэтническим этапом формирования якутов как отдельного народа. Некоторые из этих линий возникли на Ближнем Востоке, мигрировали по степному евразийскому коридору через Среднюю Азию в Южную Сибирь и появились на территории Якутии, по нашему предположению, с южными тюркоязычными предками якутов [8].
Результаты генетических исследований соответствуют версиям историков о более тесном взаимодействии якутов с эвенками, чем с юкагирами, эвенами и нганасанами, и указывают на незначительность вклада соседних палеоазиатских (чукчи, коряки) и западносибирских (кеты, селькупы) племен в генофонд якутов [5, 8]. По составу генетических компонентов якуты наиболее близки к эвенкам Центральной Якутии, что обусловлено, по-видимому, как общим происхождением из древнего южносибирского генетического субстрата, так и достаточно интенсивным обменом генов между соседними народами, преимущественно через женщин. Обмен генов между соседними популяциями якутов и эвенков привел к стиранию генетических различий между ними, что особенно ярко проявляется по линиям мтДНК и результатам полногеномного анализа, в то же время отмечается высокая степень генетических различий по линиям Y-хромосомы. Из народов соседних с Якутией регионов по своим генетическим характеристикам к якутам более близки народы Южной Сибири – прежде всего буряты и тувинцы, а также алтайцы, хакасы и монголы.
Анализ географического распределения линий Y-хромосомы и мтДНК и результаты полногеномного анализа указывают на южносибирское происхождение якутов к западу от Байкала в Восточно-Саянском регионе [8, 27, 104, 109, 110], что соответствует традиционным представлениям историков, лингвистов и этнографов об этногенезе народа саха. Результаты генетических исследований в целом хорошо согласуются и с антропологическими данными, указывающими на сходство якутов с тувинцами, бурятами Иркутской области и монголами, а также на близость якутов к древним глазковцам и хунну [40–42].
Следует отметить, что время возникновения (~2300 лет назад) и дивергенции специфических “якутских” линий Y-хромосомы, по-видимому, отражает реальные факты демографической истории популяции. Время дивергенции N3-M1982-линий якутов, рассчитанное на основе микросателлитного разнообразия и полного секвенирования Y-хромосом, указывает на первичное увеличение численности предковой популяции в IV–VII вв., за которым последовала вторичная экспансия в период с XI по XX в. с наиболее усиленным ростом численности начиная с XVI в.
На основании того, что у якутов не была обнаружена так называемая “С3-линия чингизидов” [8], ранее нами было выдвинуто предположение, что тюркоязычные предки якутов перекочевали на Север задолго до создания империи Чингисхана [14]. Это предположение подтверждается археологическими данными, полученными в последние годы. Наиболее ранние следы проникновения скотоводов на Среднюю Лену были датированы III–IV вв. Исследования археологических памятников раннего этапа кулун-атахской культуры последних лет показали, что в X–XIV вв. в бассейне Средней Лены существовала уже сложившаяся, адаптированная к местным условиям культура кочевников, успешно разводивших лошадей и крупный рогатый скот [38].
Обобщая вышеизложенное, отметим, что к настоящему времени на основе данных этногеномики (популяционной геномики) определены генетические особенности, состав компонентов генофонда и генетические взаимоотношения саха (якутов) с народами Якутии и соседних регионов. Полученные данные позволили частично реконструировать генетическую историю и ответить на ряд вопросов, касающихся этногенеза народа саха. Генетические данные подтверждают версию о местонахождении прародины тюркоязычных предков якутов к западу от Байкала в Восточно-Саянском регионе, но окончательный ответ на этот вопрос может дать в дальнейшем только сравнительный анализ древней ДНК носителей курумчинской (V–XIV вв.), усть-талькинской (XII–XIV вв.) и других археологических культур циркумбайкальского региона с линиями современных саха. Вопрос о точном времени появления “якутских” N3а2-М1982-линий на северных территориях также остается открытым, пока не будет проведен ДНК-анализ древних останков из погребений Якутии от неолита до раннего средневековья.
Из предложенных ранее гипотез по этногенезу народа саха наиболее обоснованными нам представляются версии этнографа Г.В. Ксенофонтова: колонизация Ленского края южными предками якутов с начала I тыс. н. э., существование нескольких волн миграций, переселение южных предков народа саха на Среднюю Лену до создания империи Чингис-хана, тесное взаимодействие с тунгусами [28], а также археологов А.Н. Алексеева и Р.И. Бравиной: проникновение скотоводческих групп на Среднюю Лену в первой половине I тыс. н. э., присутствие тюркоязычных носителей кулун-атахской культуры в бассейне Средней Лены в X–XIV вв. [38] и лингвистов А.В. Дыбо, Г.Г. Левина, А.В. Савельева и М. Роббитс: отмежевание якутского языка от основного пратюркского языкового ствола в IV–V вв. [32, 47] или даже ранее – во II в. [45].
Следует подчеркнуть, что одним из центральных направлений в современной медицинской геномике является поиск полигенных шкал риска предрасположенности прежде всего к распространенным мультифакториальным, а также к редким заболеваниям. Адекватное знание генетической структуры населения имеет важное значение не только для решения вопросов происхождения и генетической истории этноса, но и является основой для молекулярной эпидемиологии наследственных и мультифакториальных болезней.
Работа выполнена в рамках Государственного задания Министерства науки и высшего образования РФ (FSRG-2020-0016) “Широкогеномные исследования генофонда коренного населения арктического побережья Якутии”, НИР ЯНЦ КМП “Изучение генетической структуры и груза наследственной патологии в популяциях Республики Саха (Якутия)”, а также при поддержке гранта РФФИ (№ 20-015-00328_A).
Настоящая статья не содержит каких-либо исследований с использованием в качестве объекта животных.
Настоящая статья не содержит каких-либо исследований с участием в качестве объекта людей.
Авторы заявляют, что у них нет конфликта интересов.
Список литературы
Федорова С.А., Бермишева М.А., Виллемс Р. и др. Анализ линий митохондриальной ДНК в популяции якутов // Мол. биология. 2003. Т. 37. № 4. С. 643–653.
Федорова С.А., Хусаинова Р.И., Кутуев И.А. и др. Полиморфизм CTG-повторов гена миотонинпротеинкиназы в популяциях Республики Саха (Якутия) и Средней Азии // Мол. биология. 2005. Т. 39. № 3. С. 385–393.
Rootsi S., Zhivotovsky L.A., Baldovic M. et al. A counter clockwise northern route of the Y-chromosome haplogroup N from Southeast Asia towards Europe // Eur. J. Hum. Genet. 2007. V. 15. P. 204–211. https://doi.org/10.1038/sj.ejhg.5201748
Федорова С.А., Степанов А.Д., Адоаян М. и др. Анализ линий древней митохондриальной ДНК в Якутии // Мол. биология. 2008. Т. 42. № 3. С. 445–453.
Федорова С.А. Генетические портреты народов Республики Саха (Якутия): анализ линий митохондриальной ДНК и Y-хромосомы. Якутск: Изд-во ЯНЦ СО РАН, 2008. 235 с.
Rasmussen M., Li Y., Lindgreen S. et al. Ancient human genome sequence of an extinct Palaeo-Eskimo // Nature. 2010. V. 463. P. 757–762. https://doi.org/10.1038/nature08835
Федорова С.А. Якуты: генетические реконструкции в сравнении с историческими // Этногенез и культурогенез в Байкальском регионе / Под ред. Коновалова П.Б. Улан-Удэ: Изд-во БНЦ СО РАН, 2011. С. 151–176.
Fedorova S.A., Reidla M., Metspalu E. et al. Autosomal and uniparental portraits of the native populations of Sakha (Yakutia): implications for the peopling of Northeast Eurasia // BMC Evol. Biology. 2013. V. 13:127. https://doi.org/10.1186/1471-2148-13-127
Федорова С.А., Барашков Н.А., Ушницкий В.В. и др. Этнотерриториальные группы якутов: особенности генетической структуры // Якутский мед. журн. 2014. № 2. С. 28–31.
Yunusbayev B., Metspalu M., Metspalu E. et al. The genetic legacy of the expansion of Turkic-speaking nomads across Eurasia // PLoS Genet. 2015. V. 11(4): e1005068. https://doi.org/10.1371/journal.pgen.1005068
Karmin M., Saag L., Vicente M. et al. A recent bottleneck of Y chromosome diversity coincides with a global change in culture // Genome Res. 2015. V. 25. P. 459–466. https://doi.org/10.1101/gr.186684.114
Ilumäe A.M., Reidla M., Chukhryaeva M. et al. Human Y chromosome haplogroup N: A non-trivial time-resolved phylogeography that cuts across language families // Am. J. Hum. Genet. 2016. V. 99. P. 163–173. https://doi.org/10.1016/j.ajhg.2016.05.025
Zvénigorosky V., Crubézy E., Gibert M. et al. The genetics of kinship in remote human groups // Forensic Sci. Int. Genet. 2016. V. 25. P. 52–62.
Федорова С.А. Происхождение якутов: молекулярно-генетические реконструкции в сравнении с гипотезами историков // Якутский мед. журн. 2017. № 3(59). С. 60–64.
Zvénigorosky V., Nikolaeva D., Romanov G. et al. Persistence and disappearance of traditional patrilocality: matrimonial strategies and post-nuptial residence patterns in two Eastern Siberian communities of the twenty-first century // Sibirica. 2019. V. 18. Issue 1. P. 53–70. https://doi.org/10.3167/sib.2019.180104
Zvénigorosky V., Duchesne S., Romanova L. et al. The genetic legacy of legendary and historical Siberian chieftains // Communication Biology. 2020. V. 3(1): 581. https://doi.org/10.1038/s42003-020-01307-3
Zvénigorosky V., Sabbagh A., Gonzalez A. et al. The limitations of kinship determinations using STR data in ill-defined populations // Intern. J. Legal Med. 2020. V. 134(6). P. 1981–1990. https://doi.org/10.1007/s00414-020-02298-w
Степанов В.А. Этногеномика населения Северной Евразии. Томск: Печатная мануфактура, 2002. 243 с.
Пузырев В.П., Степанов В.А., Голубенко М.В. и др. Линии мтДНК и Y хромосомы в популяции якутов // Генетика. 2003. Т. 39. № 7. С. 975–981.
Харьков В.Н., Степанов В.А., Медведева О.Ф. и др. Происхождение якутов: анализ гаплотипов Y‑хромосомы // Мол. биология. 2008. Т. 42. № 2. С. 226–237.
Pakendorf B., Morar B., Tarskaia L.A. et al. Y-chromosomal evidence for a strong reduction in male population size of Yakuts // Hum. Genet. 2002. V. 110. P. 198–200. https://doi.org/10.1007/s00439-001-0664-4
Pakendorf B., Wiebe V., Tarskaia L.A. et al. Mitochondrial DNA evidence for admixed origins of Central Siberian populations // Am. J. Phys. Anthropol. 2003. V. 120. P. 211–224. https://doi.org/10.1002/ajpa.10145
Pakendorf B., Novgorodov I.N., Osakovskij V.L. et al. Investigating the effects of prehistoric migrations in Siberia: genetic variation and the origins of Yakuts // Hum. Genet. 2006. V. 120. P. 334–353. https://doi.org/10.1007/s00439-006-0213-2
Тарская Л.А., Мелтон Ф. Сравнительный анализ митохондриальной ДНК якутов и других монголоидных популяций // Генетика. 2006. Т. 42. № 12. С. 1703–1711.
Тарская Л.А., Гоголев А.И., Ельчинова Г.И. и др. Этническая геномика якутов (народа саха). М.: Наука, 2009. 271 с.
Zlojutro M., Tarskaia L.A., Sorensen M. et al. Coalescent simulations of Yakut mtDNA variation suggest small founding populationy // Am. J. Phys. Anthropol. 2009. V. 139(4). P. 474–482. https://doi.org/10.1002/ajpa.21003
Pugach I., Matveev R., Spitsyn V. et al. The complex admixture history and recent southern origins of Siberian populations // Mol. Biol. Evol. 2016. V. 33(7). P. 1777–1795. https://doi.org/10.1093/molbev/msw055
Ксенофонтов Г.В. Ураангхай-сахалар: Очерки по древней истории якутов. Иркутск: ОГИЗ Восточносибирское областное изд-во, 1937. Т. I. 576 с.
Бетлингк О.Н. О языке якутов. Новосибирск: Наука, 1989. 646 с.
Убрятова Е.И. Якутский язык в его отношении к другим тюркским языкам, а также к языкам монгольским и тунгусо-маньчжурским. М.: Вост. лит-ра, 1960. 13 с.
Новгородов И.Н. Якутско-эвенкийские языковые взаимосвязи: Дис. … докт. филог. наук. Казань: КГУ, 2009. 582 с.
Левин Г.Г. Исторические связи якутского языка с древними тюркскими языками VII–IX вв. (в сравнительно-сопоставительном аспекте с восточно-тюркскими и монгольскими языками). Якутск: Изд-во СВФУ, 2013. 439 с.
Окладников А.П. История Якутской АССР. В 3-х томах. Т. 1: Якутия до присоединения к русскому государству. М.; Л.: Изд-во АН СССР, 1955. 432 с.
Константинов И.В. Происхождение якутского народа и его культуры // Якутия и ее соседи в древности. Якутск, 1975. С. 106–173.
Гоголев А.И. Якуты: проблемы этногенеза и формирования культуры. Якутск: Национальное изд-во РС(Я), 1993. 136 с.
Алексеев А.Н. Древняя Якутия. Железный век и эпоха средневековья. Новосибирск: Изд-во ИАЭТ СО РАН, 1996. 95 с.
Ушницкий В.В. Проблема происхождения народа саха // Народ cаха – от века к веку. Новосибирск: Наука, 2003. С. 39–61.
Алексеев А.Н., Бравина Р.И. Формирование якутского народа // История Якутии. Новосибирск: Изд-во Наука, 2020. Т. 1. С. 223–230.
Боякова С.И. Традиционная экономика и система жизнеобеспечения // Якуты. Саха. Серия. “Народы и культуры”. М.: Наука, 2013. С. 141–150.
Левин М.Г. Антропологические типы Сибири и Дальнего Востока // Сов. этнография. 1950. № 2. С. 53–64.
Аксянова Г.А. Антропология якутов // Якуты (Саха). Серия “Народы и культуры”. М.: Наука, 2012. С. 64–78.
Томтосова Л.Ф. Новые материалы по краниологии современных якутов // Исследования по палеоантропологии и краниологии СССР: Сб. МАЭ. Л.: Наука, 1980. Т. 36. С. 121–129.
Мудрак О.А. Классификация тюркских языков и диалектов с помощью методов глоттохронологии на основе вопросов по морфологии и исторической фонетике. Серия “Orientalia и Classica”. Вып. 23. М.: РГГУ, 2009. 186 с.
Широбокова Н.Н. Отношение якутского языка к тюркским языкам Южной Сибири. Новосибирск: Наука, 2005. 269 с.
Дыбо A.B. Хронология тюркских языков и лингвистические контакты ранних тюрков. М.: Академия, 2004. 811 с.
Robbeets M., Bouckaert R. Bayesian phylolinguistics reveals the internal structure of the Transeurasian family // J. Language Evol. 2018. V. 3(2). P. 145–162. https://doi.org/10.1093/jole/lzy007
Savelyev A., Robbeets M. Bayesian phylolinguistics infers the internal structure and the time-depth of the Turkic language family // J. Language Evol. 2020. V. 5(1). P. 39–53. https://doi.org/10.1093/jole/lzz010
Певнов А.М. Лингвистические свидетельства истории народов Якутии // История Якутии. Т. 1. Новосибирск: Наука, 2020. С. 273–280.
Дашибалов Б.Б. Археологические памятники курыкан и хори. Улан-Удэ: Изд-во БНЦ СО РАН, 1995. 189 с.
Николаев В.С. Средневековые погребальные комплексы по обряду захоронения человека и коня в Предбайкалье и проблема этногенеза якутов // Этноистория и археология Северной Евразии: теория, методология и практика исследования. Иркутск; Эдмонтон: ИрГТУ, 2007. С. 156–163.
Бравина Р.И., Дьяконов В.М., Николаев Е.Н. и др. Комплексное исследование раннеякутского Сергеляхского погребения середины XV–начала XVI в. // Вестник археологии, антропологии и этнографии. 2016. № 4(35). С. 90–109.
Савинов Д.Г. Дотюркский пласт в палеоэтнографии якутов // Сибирский сборник. СПб.: МАЭ РАН, 2010. С. 68–81.
Эргис Г.У. Исторические предания и рассказы якутов: в 2 частях. М.; Л.: Изд-во АН СССР, 1960. Ч. 1. 322 с.
Zerjal T., Dashnyam B., Pandya A. et al. Genetic relationships of Asians and Northern Europeans, revealed by Y-chromosomal DNA analysis // Am. J. Hum. Genet. 1997. V. 60. P. 1174–1183.
Van Oven M., Kayser M. Updated comprehensive phylogenetic tree of global human mitochondrial DNA variation // Hum. Mutat. 2009. V. 30. E386–E394. https://doi.org/10.1002/humu.20921
Kılınç G.M., Kashuba N., Koptekin D. et al. Human population dynamics and Yersinia pestis in ancient northeast Asia // Sci. Adv. 2021. V. 7. eabc4587. https://doi.org/10.1126/sciadv.abc4587
Karafet T.M., Osipova L.P., Gubina M.A. et al. High levels of Y-chromosome differentiation among native Siberian populations and the genetic signature of a boreal hunter-gatherer way of life // Hum. Biol. 2002. V. 74. P. 761–789. https://doi.org/10.1353/hub.2003.0006
Информационные материалы об окончательных итогах Всероссийской переписи населения 2010 года http://www.gks.ru/free_doc/new_site/perepis2010/perepis_itogi1612.htm
Dabernat H., Thèves C., Bouakaze C. et al. Tuberculosis epidemiology and selection in an autochthonous Siberian population from the 16th–19th century // PLoS One. 2014. V. 9(2): e89877. https://doi.org/10.1371/journal.pone.0089877
Долгих Б.О. Родовой и племенной состав народов Сибири в XVII веке. М.: Изд-во АН СССР, 1960. 622 с.
Серошевский В.Л. Якуты. Опыт этнографического исследования. Т. 1. Санкт-Петербург: Типография Главного Управления Уделов, 1896. 720 с.
Брагина Д.Г. Об этнической идентичности якутов в Республике Саха (Якутия) // The Scientific Heritage. 2021. № 76. P. 59–62.
Кучер А.Н., Данилова А.Л., Конева Л.А., Ноговицина А.Н. Структура браков в якутских популяциях: национальный состав и инбридинг по изонимии // Генетика. 2010. Т. 46. № 3. С. 408–416.
Данилова А.Л. Генетико-демографическое исследование народонаселения Республики Саха (Якутия): Дис. … канд. биол. наук. Томск: НИИ медицинской генетики, 2009. 198 с.
Игнатьева В.Б. Этногеографические группы. Демография и территориальное расселение якутов // Якуты. Саха. Серия “Народы и культуры”. М.: Наука, 2013. С. 20–31.
Слепцов П.А. Семья и семейная обрядность у якутов (XIX–начало XX века): Дис. … канд. ист. наук. Ленинград: ЛГУ, 1984. 238 с.
Derenko M., Malyarchuk B., Denisova G.A. et al. Contrasting patterns of Y-chromosome variation in South Siberian populations from Baikal and Altai-Sayan regions // Hum. Genet. 2006. V. 118(5). P. 591–604. https://doi.org/10.1007/s00439-005-0076-y
Derenko M., Malyarchuk B., Denisova G. et al. Y-chromosome haplogroup N dispersals from south Siberia to Europe // J. Hum. Genet. 2007. V. 52(9). P. 763–770. https://doi.org/10.1007/s10038-007-0179-5
Харьков В.Н. Структура и филогеография генофонда коренного населения Сибири по маркерам Y-хромосомы: Дис. … докт. биол. наук. Томск: НИИ мед. генетики, 2012. 440 с.
Wells R.S., Yuldasheva N., Ruzibakiev R. et al. The Eurasian heartland: A continental perspective on Y‑chromosome diversity // Proc. Natl Acad. Sci. USA. 2001. V. 98. P. 10244–10249. https://doi.org/10.1073/pnas.171305098
Кутуев И.А. Генетическая структура и молекулярная филогеография народов Кавказа. Дис. … докт. биол. наук. Уфа, 2010. 305 с.
Yunusbayev B., Metspalu M., Järve M. et al. The Caucasus as an asymmetric semipermeable barrier to ancient human migrations // Mol. Biol. Evol. 2012. V. 29(1). P. 359–365. https://doi.org/10.1093/molbev/msr221
Hallast P., Batini C., Zadik D. et al. The Y-chromosome tree bursts into leaf: 13 000 high-confidence SNPs covering the majority of known clades // Mol. Biol. Evol. 2015. V. 32 (3). P. 661–673. https://doi.org/10.1093/molbev/msu327
Rootsi S., Magri C., Kivisild T. et al. Phylogeography of Y-chromosome haplogroup I reveals distinct domains of prehistoric gene flow in Europe // Am. J. Hum. Genet. 2004. V. 75(1). P. 128–137. https://doi.org/10.1086/422196
Semino O., Passarino G., Oefner P.J. et al. The genetic legacy of Paleolithic Homo sapiens in extant Europeans: a Y-chromosome perspective // Science. 2000. V. 290. P. 1155–1159. https://doi.org/10.1126/science.290.5494.1155
Balanovsky O., Rootsi S., Pshenichnov A. et al. Two sources of the Russian patrilineal heritage in their Eurasian context // Am. J. Hum. Genet. 2008. V. 82(1). P. 236–250. https://doi.org/10.1016/j.ajhg.2007.09.019
Underhill P.A., Myres N.M., Rootsi S. et al. Separating the post-Glacial coancestry of European and Asian Y chromosomes within haplogroup R1a // Eur. J. Hum. Genet. 2010. V. 18 (4). P. 479–484. https://doi.org/10.1038/ejhg.2014.50
Кушнеревич Е.И., Сивицкая Л.Н., Богачева А.В. и др. Гаплогруппа R1a1a7 (М-458) Y-хромосомы современных белорусов и миграции предков славян на территории Беларуси // Генетика популяций и эволюция. 2011. Т. 9(1). С. 44–52.
Казарян П.Л. Итоги и задачи исследования пребывания поляков в якутской ссылке (XVII–начало XX вв.) // Ссыльные поляки в Якутии: итоги, задачи, исследование пребывания. Cб. науч. трудов. Якутск: ИГИ АН РС(Я), 1999. С. 6–30.
Keyser C., Bouakaze C., Crubézy E. et al. Ancient DNA provides new insights into the history of south Siberian Kurgan people // Hum. Genet. 2009. V. 126(3). P. 395–410. https://doi.org/10.1007/s00439-009-0683-0
González-Ruiz M., Santos C., Jordana X. et al. Tracing the origin of the east-west population admixture in the Altai region (Central Asia) // PLoS One. 2012. V. 7(11). e48904. https://doi.org/10.1371/journal.pone.0048904
Pilipenko A.S., Trapezov R.O., Cherdantsev S.V. et al. Maternal genetic features of the Iron Age Tagar population from Southern Siberia (1st millennium BC) // PLoS One. 2018. V. 13(9): e0204062. https://doi.org/10.1371/journal.pone.0204062
Kim K., Brenner C.H., Mair V.H. et al. A western Eurasian male is found in 2000-year-old elite Xiongnu cemetery in Northeast Mongolia // Am. J. Phys. Anthropol. 2010. V. 142(3). P. 429–440. https://doi.org/10.1002/ajpa.21242
Malyarchuk B., Grzybowski T., Derenko M. et al. Mitochondrial DNA variability in Poles and Russians // Ann. Hum. Genet. 2002. V. 66. P. 261–283. https://doi.org/10.1016/j.fsigen.2017.06.003
Malyarchuk B., Derenko M., Grzybowski T. et al. Differentiation of mitochondrial DNA and Y-chromosomes in Russian populations // Hum. Biol. 2004. V. 76. P. 877–900. https://doi.org/10.1353/hub.2005.0021
Бермишева М.А., Тамбетс К., Виллемс Р., Хуснутдинова Э.К. Разнообразие гаплогрупп митохондриальной ДНК у народов Волго-Уральского региона // Мол. биология. 2002. Т. 36. С. 802–812.
Belyaeva O., Bermisheva M., Khrunin A. et al. Mitochondrial DNA variations in Russian and Belorussian populations // Hum. Biol. 2003. V. 75. P. 647–660. https://doi.org/10.1353/hub.2003.0069
Фишер И. Сибирская история с самого открытия Сибири до завоевания сей земли Российским оружием. СПб.: Имперская Акад. наук, 1774. 632 с.
Сафронов Ф.Г. Распространение земледелия на северо-востоке Сибири в XVII–начале ХХ века // Исторические связи народов Якутии с русским народом: Сб. науч. трудов. Якутск: Якутское кн. изд-во, 1987. С. 28–39.
Романов И.Г. Формирование русского населения Якутии (1917–1941 гг.). Якутск: Изд-во ИГИ АН РС(Я), 1998. 217 с.
Ricaut F.-X., Kolodesnikov S., Keyser-Tracqui C. et al. Molecular genetic analysis of 400-year-old human remains found in two yakut burial sites // Am. J. Phys. Anthropol. 2006. V. 129. P. 55–63. https://doi.org/10.1002/ajpa.20195
Grzybowski T., Malyarchuk B.A., Derenko M.V. et al. Complex interactions of the Eastern and Western Slavic populations with other European groups as revealed by mitochondrial DNA analysis // Forensic Sci. Int. Genet. 2007. V. 1. P. 141–147. https://doi.org/10.1016/j.fsigen.2007.01.010
Morozova I., Evsyukov A., Kon’kov A. et al. Russian ethnic history inferred from mitochondrial DNA diversity // Am. J. Phys. Anthropol. 2012. V. 147(3). P. 341–351. https://doi.org/10.1002/ajpa.21649
Loogvali E.-L., Roostalu U., Malyarchuk B.A. et al. Disuniting uniformity: A pied cladistic canvas of mtDNA haplogroup H in Eurasia // Mol. Biol. Evol. 2004. V. 21. P. 2012–2021. https://doi.org/10.1093/molbev/msh209
Derenko M., Malyarchuk B., Grzybowski T. et al. Phylogeographic analysis of mitochondrial DNA in Northern Asian populations // Am. J. Hum. Genet. 2007. V. 81. P. 1025–1041. https://doi.org/10.1086/522933
Roostalu U., Kutuev I., Loogväli E.L. et al. Origin and expansion of haplogroup H, the dominant human mitochondrial DNA lineage in West Eurasia: the Near Eastern and Caucasian perspective // Mol. Biol. Evol. 2007. V. 24(2). P. 436–448. https://doi.org/10.1093/molbev/msl173
Behar D.M., Metspalu E., Kivisild T. et al. Counting the founders: the matrilineal genetic ancestry of the Jewish Diaspora // PLoS One. 2008. V. 3(4): e2062. https://doi.org/10.1371/journal.pone.0002062
Derbeneva O.A., Starikovskaya E.B., Wallace D.C., Sukernik R.I. Traces of Early Eurasians in the Mansi of Northwest Siberia revealed by mitochondrial DNA analysis // Am. J. Hum. Genet. 2002. V. 70. P. 1009–1014. https://doi.org/10.1086/339524
Гольцова Т.В., Осипова Л.П., Жаданов С.И., Виллемс Р. Влияние брачной миграции на генетическую структуру популяции нганасан Таймыра: генеалогический анализ по маркерам митохондриальной ДНК // Генетика. 2005. Т. 41. № 7. С. 954–965.
Li J.Z., Absher D.M., Tang H. et al. Worldwide human relationships inferred from genome-wide patterns of variation // Science. 2008. V. 319. P. 1100–1104. https://doi.org/10.1126/science.1153717
Долгих Б.О. Происхождение долган // Сиб. этнографический сборник. 1963. Т. 5. С. 92–141.
Окладников А.П., Деревянко А.П. Далекое прошлое Приморья и Приамурья. Владивосток: Дальневост. кн. изд-во, 1973. 440 с.
Певнов А.М. Лингвистические пути решения тунгусо-маньчжурской проблемы // Вопр. языкознания. 2008. № 5. С. 64–81.
Robbeets M., Bouckaert R., Conte M. et al. Triangulation supports agricultural spread of the Transeurasian languages // Nature. 2021. V. 599. P. 616–621. https://doi.org/10.1038/s41586-021-04108-8
Zerjal T., Xue Y., Bertorelle G. The genetic legacy of the Mongols // Am. J. Hum. Genet. 2003. V. 72(3). P. 717–721. https://doi.org/10.1086/367774
Cardona A., Pagani L., Antao T. et al. Genome-wide analysis of cold adaptation in indigenous Siberian populations // PLoS One. 2014. V. 9(5): e98076. https://doi.org/10.1371/journal.pone.0098076
Zhernakova D.V., Brukhin V., Malov S. et al. Genome-wide sequence analyses of ethnic populations across Russia // Genomics. 2020. V. 112(1). P. 442–458. https://doi.org/10.1016/j.ygeno.2019.03.007
Wong E.H.M., Khrunin A., Nichols L. et al. Reconstructing genetic history of Siberian and Northeastern European populations // Genome Res. 2017. V. 27(1). P. 1–14. https://doi.org/10.1101/gr.202945.115
Damgaard P.B., Martiniano R., Kamm J. et al. The first horse herders and the impact of early Bronze Age steppe expansions into Asia // Science. 2018. V. 360(6396): eaar7711. https://doi.org/10.1126/science.aar7711
Jeong C., Wang K., Wilkin S. et al. A dynamic 6000-year genetic history of Eurasia’s Eastern steppe // Cell. 2020. V. 183(4). P. 890–904. https://doi.org/10.1016/j.cell.2020.10.015
Keyser C., Zvénigorosky V., Gonzalez A. et al. Genetic evidence suggests a sense of family, parity and conquest in the Xiongnu Iron Age nomads of Mongolia // Hum. Genet. 2020. V. 140(2). P. 349–359. https://doi.org/10.1007/s00439-020-02209-4
Дополнительные материалы отсутствуют.