

Генетика, 2022, T. 58, № 3, стр. 332-342
Популяционно-генетическая характеристика тувинских короткожирнохвостых овец
С. В. Бекетов 1, *, Е. А. Коноров 1, А. К. Пискунов 1, В. Н. Воронкова 1, С. В. Каштанов 1, Т. Е. Денискова 2, О. А. Кошкина 2, М. И. Селионова 3, Ю. А. Столповский 1
1 Институт общей генетики им. Н.И. Вавилова Российской академии наук
119991 Москва, Россия
2 Федеральный исследовательский центр животноводства им. академика Л.К. Эрнста
142132 п. Дубровицы, г/о Подольск, Московская область, Россия
3 Российский государственный аграрный университет – Московская сельскохозяйственная академия
им. К.А. Тимирязева
127550 Москва, Россия
* E-mail: svbeketov@gmail.com
Поступила в редакцию 05.07.2021
После доработки 13.09.2021
Принята к публикации 21.09.2021
- EDN: SZFHBB
- DOI: 10.31857/S0016675822030031
Аннотация
С использованием 11 микросателлитных маркеров получены данные о генетической структуре, уровнях изменчивости и степени дифференциации 23 популяций (1110 голов) тувинской грубошерстной короткожирнохвостой овцы разных районов Республики Тыва. Установлено, что в настоящее время эта автохтонная порода представлена двумя основными типами (горным и степным), а также многочисленными местными промежуточными популяциями. Несмотря на обнаруженное аллельное разнообразие от 7502 до 10 114, во всех исследованных популяциях был выявлен достоверный дефицит гетерозигот – от 15.3 до 35.6%. При этом межпопуляционные различия в группе тувинских овец степного типа (FST = 0.013–0.153, Jost’s D = 0.029–0.461) оказались более выражены, чем в группе тувинских овец горного типа (FST = 0.01–0.071, Jost’s D = 0.029–0.247). Одной из возможных причин неоднородной дифференциации популяционной структуры тувинской короткожирнохвостой овцы является адаптация к различным природно-климатическим условиям, включая типологический состав и продуктивность пастбищ на территории Тувы.
Сельское хозяйство Республики Тыва традиционно ориентировано на отгонно-пастбищное скотоводство, в котором овцеводство является основной подотраслью, которая обеспечивает две трети объема валовой продукции животноводства [1]. При этом 30.8% грубошерстных мясных овец Российской Федерации сосредоточено в Туве [2], где при общей численности овец более 880 тыс. голов тувинская грубошерстная короткожирнохвостая овца является фактически моно-породой [3, 4], а остальные 60% поголовья овец приходятся на долю полугрубошерстных помесей, получаемых от скрещивания пород-улучшателей (преимущественно сараджинской) с местной тувинской овцой [3].
Грубошерстная тувинская короткожирнохвостая овца – это древняя аборигенная порода, которая характеризуется исключительной жизнеспособностью и приспособленностью к суровым природно-климатическим условиям данного региона Южной Сибири [5].
Сложно сказать, когда именно сформировался современный тип тувинской овцы, однако курганные находки дают вполне ясное представление о том, что уже у древних кочевников на территории современной Тувы в составе стад преобладали мясные жирнохвостые овцы [6]. Развитию отгонно-пастбищного животноводства на этой территории в значительной степени способствовали горно-котловинный рельеф местности, а также природно-климатические условия [7, 8].
Так как горы покрывают 82% Республики Тыва, естественные пастбища сосредоточены здесь преимущественно в межгорных равнинных впадинах [9], предгорных степях, а также на низких террасах крупных рек, причем с учетом выраженной высотной зональности и разнообразия почв зачастую даже в пределах одного животноводческого хозяйства отмечаются разные почвенно-климатические и кормовые условия [10].
Всего на кормовые угодья приходится до 25% площади Тувы или до 99% всех естественных пастбищ [7, 8]. При этом в силу своеобразия рельефа местности наибольшее поголовье овец сосредоточено в западном (46.72%) и южном (30.83%) районах республики [11].
Климат Тувы – ультраконтинентальный со значительной годовой амплитудой температур и малым количеством осадков, при этом отмечаются ежегодные засухи и зимнее глубокое промерзание почвы [12].
То есть можно сказать, что на формирование тувинских овец существенное влияние оказали три основных фактора: внешняя среда, кормовые условия и традиционная технология круглогодичного отгонно-пастбищного содержания.
Местные тувинские грубошерстные короткожирнохвостые овцы отличаются выносливостью, скороспелостью и способностью к быстрому накоплению жировых отложений после зимы [13]. Помимо этого, важной экологической приспособленностью является невысокая плодовитость овцематок (105–110%) при выраженном материнском инстинкте, направленном на максимальное сохранение потомства [14].
По своей конституции тувинские овцы – крепкие, со слега облегченным костяком, копыта прочные, голова небольшая, длинная, узкая, с часто встречающейся горбоносостью, окрашена в черный, иногда бурый и рыжий цвета с переходом окраски у некоторых животных почти на всю шею. Большинство баранов имеет массивные спиральные рога; матки, как правило, комолые. Животные чаще всего белые по окраске руна. Форма хвоста варьирует от жировой до клинообразной [1, 13–15].
Тувинские короткожирнохвостые овцы имеют сравнительно небольшую живую массу и характеризуются невысокой шерстной продуктивностью [16]. Шерсть – грубая, с большим содержанием сухих и мертвых волокон [3].
Неравномерный состав травостоя, непостоянство урожайности пастбищ и контрастность природно-климатических условий территории Тувы наложили свои особенности на селекционно-племенную работу с тувинской овцой. Это привело к тому, что к настоящему времени имеются два внутрипородных типа тувинских овец: горный и степной, которые отличаются морфометрическими показателями и продуктивностью. Оба типа были утверждены в 2010 г. [17].
Тем не менее, несмотря на изменения, которые произошли с тувинской овцой за последние десятилетия, опубликовано всего лишь две работы, отражающие степень генетического разнообразия местных овец. В одной из них впервые было применено ISSR-маркирование тувинской короткожирнохвостой овцы [18], во второй изучали полиморфизм генов-кандидатов хозяйственно полезных признаков овец: CAST, GDF9, IGFBP-3, GHo [19].
Еще две недавние публикации были посвящены полногеномному генотипированию отдельной выборочной популяции тувинских овец в сравнении с другими отечественными породами [20, 21].
И хотя имеется большой интерес исследователей к изучению тувинской грубошерстной короткожирнохвостой овцы, работ, отражающих современное состояние генетической структуры этой уникальной породы, до сих пор не проводили.
Одним из эффективных методов исследования генетической изменчивости популяций является микросателлитное ДНК-профилирование групп и отдельных организмов. В частности, микросателлитные маркеры широко используются для генетического анализа популяций автохтонных пород овец [22–25], а также диких сородичей домашней овцы [26]. В связи с чем основной целью настоящего исследования стало проведение сравнительного микросателлитного анализа нескольких популяций тувинских грубошерстных короткожирнохвостых овец разных районов Республики Тыва.
МАТЕРИАЛЫ И МЕТОДЫ
Материалом для исследований были образцы биоматериала (кровь) 23 популяций тувинских грубошерстных короткожирнохвостых овец из отдельных хозяйств девяти административных районов (кожуунов) Республики Тыва (n = 1110) (табл. 1).
Таблица 1.
Местоположение и координаты овцеводческих хозяйств
| № п/п | Наименование популяции | Хозяйство, населенный пункт | Координаты | Высота над уровнем моря, м | |
|---|---|---|---|---|---|
| северная широта | восточная долгота | ||||
| 1 | Aylyg-Oy | СПК “Айлыг-Ой” (с. Теве-Хая, Дзун-Хемчикский кожуун) | 51°22′00′′ | 91°27′00′′ | 729 |
| 2 | Ak-Khaya | СПК “Ак-Хая” (с. Шеми, Дзун-Хемчикский кожуун) | 51°13′00′′ | 91°17′60′′ | 966 |
| 3 | Dag-Uzu | СПК “Даг-Ужу” (с. Хондергей, Дзун-Хемчикский кожуун) | 51°07′00′′ | 91°37′00′′ | 1042 |
| 4 | Bai-Tal | СПК “Бай-Тал” (с. Хондергей, Дзун-Хемчикский кожуун) | 51°07′00′′ | 91°37′00′′ | 1042 |
| 5 | Mongush | СПК “Монгуш” (с. Теве-Хая, Кызылский кожуун) | 51°43′00′′ | 94°25′60′′ | 607 |
| 6 | Irtysh | СПК “Иртиш” (п. г. т. Каа-Хем, Кызылский кожуун) | 51°42′00′′ | 94°32′60′′ | 621 |
| 7 | Alaak | СПК “Алаак” (с. Суг-Аксы, Сут-Хольский кожуун) | 51°23′60′′ | 91°16′60′′ | 713 |
| 8 | Amyk | СПК “Амык” (с. Алдан-Маадыр, Сут-Хольский кожуун) | 51°17′60′′ | 90°58′00′′ | 756 |
| 9 | Murgustug | СПК “Мургустуг” (с. Хандагайты, Овюрский кожуун) | 50°43′60′′ | 92°04′00′′ | 1157 |
| 10 | Torgalyg | МУП “Торгалыг” (п. Дус-Даг, Овюрский кожуун) | 50°48′53′′ | 92°40′54′′ | 1260 |
| 11 | Adargan | МУП “Адарган” (с. Саглы, Овюрский кожуун) | 50°30′00′′ | 91°19′60′′ | 1574 |
| 12 | Bai-Hol | Племенное СПК “Бай-Хол” (с. Бай-Даг, Эрзинский кожуун) | 50°17′60′′ | 95°05′60′′ | 1074 |
| 13 | Yamaalyk | СПК “Ямаалык” (с. Эрзин, Эрзинский кожуун) | 50°16′00′′ | 95°10′00′′ | 1102 |
| 14 | Mandala | СПК “Мандала” (с. Морен, Эрзинский кожуун) | 50°19′00′′ | 95°22′60′′ | 1193 |
| 15 | Sai-Khonash | МУП “Сай-Хонаш” (с. Кызыл-Мажалык, Барун-Хемчикский район) | 51°07′60′′ | 90°34′00′′ | 846 |
| 16 | Duza | СПК “Дуза” (с. Кызыл-Мажалык, Барун-Хемчикский кожуун) | 51°07′60′′ | 90°34′00′′ | 846 |
| 17 | Oorug | СПК “Ооруг” (с. Кызыл-Даг, Бай-Тайгинский кожуун) | 51°01′60′′ | 90°05′60′′ | 967 |
| 18 | Sukpak | СПК “Сукпак” (с. Тээли, Бай-Тайгинский кожуун) | 51°01′00′′ | 90°13′00′′ | 982 |
| 19 | Despen | МУП “Племенное овцеводческое хозяйство “Деспен” (с. О-Шимаа, Тес-Хемский кожуун) | 50°40′00′′ | 93°45′00′′ | 817 |
| 20 | Choduraa | ГУП РТ “Чодураа” (с. Бельдир-Арыг, Тес-Хемский кожуун) | 50°34′60′′ | 94°58′00′′ | 1201 |
| 21 | Beldir | СПК “Белдир” (с. Самагалтай, Тес-Хемский кожуун) | 50°37′00′′ | 95°00′00′′ | 1285 |
| 22 | Malchyn | ГУП РТ “Малчын” (с. Мугур-Аксы, Монгун-Тайгинский кожуун) | 50°22′60′′ | 90°25′60′′ | 1837 |
| 23 | Mogen-Buren | ГУП РТ “Моген-Бурен” (с. Кызыл-Хая, Монгун-Тайгинский кожуун) | 50°02′60′′ | 89°52′00′′ | 2031 |
Выделение ДНК из образцов крови проводили с использованием наборов “ДНК-Экстран” (ЗАО “Синтол”, Россия) согласно протоколу фирмы-изготовителя. Количественную и качественную оценку полученных препаратов ДНК определяли на микроспектрофотометре Nano Drop 8000 (Thermo Fisher, США): концентрацию ДНК – по степени поглощения при 260 нм и чистоту ДНК – по соотношению поглощения при длине волн 260 и 280 нм.
Исследование тувинских овец осуществляли по 11 STR-маркерам (авторская панель ОНИС БиоТехЖ) с использованием двух мультиплексных ПЦР-реакций шести (панель 1 – ОarCP49, INRA063, HSC, OarAE129, MAF214, OarFCB11) и пяти (панель 2 – INRA005, SPS113, INRA23, MAF65, McM527) локусов микросателлитов.
После начальной денатурации (95°С, 240 с) проводили 41 (панель 1) и 35 (панель 2) циклов амплификации в следующем температурно-временном режиме: 95°С, 20 с; 63°С (панель 1) и 55°С (панель 2), 30 с; 72°С, 60 с.
Капиллярный электрофорез осуществляли на генетическом анализаторе ABI 3130xl (Applied Biosystems, США) с последующим определением длин аллелей микросателлитов в программном обеспечении GeneMapper v 4.0 (Applied Biosystems).
Для оценки структуры популяций тувинской овцы использовали следующие показатели: наблюдаемая (HO) и несмещенная ожидаемая гетерозиготность (HE(u)) [27], среднее число аллелей на локус (NA), число эффективных аллелей на локус (NE), количество информативных аллелей с частотой более 5% (NA ≥ 5%), аллельное разнообразие (AR) [28], коэффициент инбридинга (FIS).
Степень генетической дифференциации исследуемых популяций тувинской овцы определяли на основании попарных генетических различий (индекс фиксации FST) [29] и попарных генетических дистанций по Jost’s D [30]. Филогенетические отношения устанавливали путем определения построения Neighbor-net дендрограмм [31].
Оценку генетической структуры популяций осуществляли с помощью адмикс-модели. Анализ базы данных STR-вариабельности популяций тувинской овцы проводили для k от 1 до 10 при длине burn-in периода – 100 000, модель Марковских цепей Монте-Карло (MCMC) – 1 000 000 повторов.
Для формирования исходных файлов и проведения анализа использовали R-версию 3.3.2 [32], программное обеспечение GenAlEx 6.5 [33], R-пакет “diveRsity”, STRUCTURE 2.3.4 [34] с последующей визуализацией в “pophelper” [35].
РЕЗУЛЬТАТЫ
В результате проведенных исследований с использованием STR-маркеров были определены генотипы 1110 голов тувинских овец. Как показано в табл. 2, число аллелей в расчете на локус микросателлитов в проанализированных популяциях овец варьировало от 8.364 ± 0.607 (Mandala) до 11.727 ± 0.875 (Ak-Khayа), аллельное разнообразие – от 7.502 ± 0.541 до 10.114 ± 0.702 соответственно. При этом минимальное число эффективных аллелей было обнаружено в популяциях Sukpak – 4.615 ± 0.476 и Sai-Khonash – 4.678 ± ± 0.598 (табл. 2). Во всех исследованных популяциях был выявлен достоверный дефицит гетерозигот (родственное спаривание) – от 15.3% в группе животных Sukpak, до 35.6% в популяции Beldir (табл. 3). Кроме того, сравнение значений наблюдаемой гетерозиготности (HO) – 0.520–0.642 и несмещенной ожидаемой гетерозиготности (HE(u)) – 0.753–0.832 (табл. 3) указывает на то, что в рассматриваемых популяциях преобладают инбредные спаривания.
Таблица 2.
Параметры аллельного разнообразия в изучаемых популяциях тувинских овец
| Популяция | n | NA | NE | NA ≥ 5% | AR |
|---|---|---|---|---|---|
| Mongush | 50 | 10.455 ± 1.003 | 5.503 ± 0.478 | 6.091 ± 0.530 | 9.114 ± 0.833 |
| Irtysh | 50 | 10.273 ± 0.662 | 5.403 ± 0.600 | 5.455 ± 0.623 | 8.820 ± 0.553 |
| Alaak | 51 | 10.000 ± 0.915 | 5.742 ± 0.700 | 5.727 ± 0.428 | 8.526 ± 0.823 |
| Aylyg-Oy | 40 | 9.455 ± 0.994 | 4.928 ± 0.586 | 5.818 ± 0.553 | 8.341 ± 0.916 |
| Amyk | 49 | 10.636 ± 1.038 | 5.540 ± 0.847 | 5.818 ± 0.658 | 8.632 ± 0.992 |
| Despen | 40 | 9.273 ± 1.335 | 4.800 ± 0.760 | 5.455 ± 0.743 | 8.103 ± 1.177 |
| Beldir | 50 | 10.636 ± 0.993 | 5.265 ± 0.613 | 5.636 ± 0.491 | 8.872 ± 0.886 |
| Duza | 50 | 11.273 ± 1.121 | 5.588 ± 0.609 | 5.636 ± 0.664 | 9.222 ± 0.857 |
| Ak-Khayа | 48 | 11.727 ± 0.875 | 6.282 ± 0.750 | 6.000 ± 0.588 | 10.114 ± 0.702 |
| Oorug | 50 | 10.000 ± 0.934 | 5.255 ± 0.627 | 5.636 ± 0.607 | 8.817 ± 0.824 |
| Sukpak | 50 | 9.818 ± 0.711 | 4.615 ± 0.476 | 5.727 ± 0.541 | 8.039 ± 0.747 |
| Dag-Uzu | 48 | 10.364 ± 0.778 | 5.835 ± 0.463 | 5.909 ± 0.392 | 8.793 ± 0.706 |
| Bai-Tal | 50 | 11.091 ± 0.879 | 6.337 ± 0.730 | 6.818 ± 0.615 | 9.714 ± 0.777 |
| Bai-Hol | 50 | 10.545 ± 1.048 | 5.933 ± 0.760 | 6.273 ± 0.557 | 9.026 ± 0.907 |
| Yаmaalyk | 50 | 10.727 ± 0.964 | 5.488 ± 0.645 | 5.818 ± 0.377 | 9.121 ± 0.838 |
| Murgustug | 42 | 10.545 ± 0.846 | 5.438 ± 0.597 | 5.091 ± 0.392 | 8.941 ± 0.685 |
| Mandala | 50 | 8.364 ± 0.607 | 4.346 ± 0.359 | 4.909 ± 0.315 | 7.502 ± 0.541 |
| Choduraa | 50 | 10.273 ± 0.727 | 5.324 ± 0.725 | 6.000 ± 0.603 | 8.811 ± 0.740 |
| Torgalyg | 50 | 10.273 ± 0.915 | 5.194 ± 0.525 | 5.182 ± 0.400 | 8.463 ± 0.809 |
| Adargan | 50 | 11.273 ± 0.810 | 5.217 ± 0.659 | 5.455 ± 0.623 | 9.180 ± 0.740 |
| Malchyn | 50 | 11.545 ± 0.846 | 5.843 ± 0.543 | 5.909 ± 0.436 | 9.710 ± 0.690 |
| Mogen-Buren | 50 | 9.818 ± 0.893 | 4.950 ± 0.456 | 5.455 ± 0.390 | 7.947 ± 0.649 |
| Sai-Khonash | 42 | 9.000 ± 1.000 | 4.678 ± 0.598 | 4.545 ± 0.474 | 7.775 ± 0.905 |
Таблица 3.
Параметры генетического разнообразия в изучаемых популяциях тувинских овец
| Популяция | n | HO | HE(u) | FIS (FIS 95%CI) |
|---|---|---|---|---|
| Mongush | 50 | 0.609 ± 0.058 | 0.804 ± 0.028 | 0.246 (0.128–0.364) |
| Irtysh | 50 | 0.581 ± 0.074 | 0.791 ± 0.034 | 0.268 (0.106–0.430) |
| Alaak | 51 | 0.567 ± 0.089 | 0.802 ± 0.029 | 0.308 (0.111–0.505) |
| Aylyg-Oy | 40 | 0.581 ± 0.071 | 0.769 ± 0.034 | 0.233 (0.031–0.435) |
| Amyk | 49 | 0.549 ± 0.077 | 0.769 ± 0.048 | 0.284 (0.100–0.468) |
| Despen | 40 | 0.545 ± 0.085 | 0.692 ± 0.071 | 0.215 (0.041–0.389) |
| Beldir | 50 | 0.520 ± 0.080 | 0.786 ± 0.033 | 0.356 (0.194–0.518) |
| Duza | 50 | 0.607 ± 0.064 | 0.794 ± 0.032 | 0.238 (0.103–0.373) |
| Ak-Khay | 48 | 0.617 ± 0.061 | 0.832 ± 0.025 | 0.263 (0.137–0.389) |
| Oorug | 50 | 0.576 ± 0.065 | 0.784 ± 0.036 | 0.277 (0.161–0.393) |
| Sukpak | 50 | 0.653 ± 0.060 | 0.768 ± 0.028 | 0.153 (0.019–0.287) |
| Dag-Uzu | 48 | 0.538 ± 0.071 | 0.824 ± 0.021 | 0.352 (0.194–0.510) |
| Bai-Tal | 50 | 0.625 ± 0.074 | 0.826 ± 0.025 | 0.250 (0.095–0.405) |
| Bai-Hol | 50 | 0.608 ± 0.045 | 0.804 ± 0.034 | 0.250 (0.182–0.318) |
| Yamaalyk | 50 | 0.598 ± 0.067 | 0.805 ± 0.025 | 0.272 (0.144–0.400) |
| Murgustug | 42 | 0.603 ± 0.060 | 0.794 ± 0.033 | 0.252 (0.147–0.357) |
| Mandala | 50 | 0.642 ± 0.070 | 0.768 ± 0.022 | 0.164 (–0.002–0.330) |
| Choduraa | 50 | 0.620 ± 0.061 | 0.785 ± 0.033 | 0.220 (0.117–0.323) |
| Torgalyg | 50 | 0.572 ± 0.062 | 0.789 ± 0.026 | 0.280 (0.146–0.414) |
| Adargan | 50 | 0.584 ± 0.060 | 0.761 ± 0.048 | 0.252 (0.175–0.329) |
| Malchyn | 50 | 0.580 ± 0.065 | 0.815 ± 0.024 | 0.290 (0.142–0.438) |
| Mogen-Buren | 50 | 0.620 ± 0.072 | 0.776 ± 0.036 | 0.218 (0.070–0.366) |
| Sai-Khonash | 42 | 0.561 ± 0.066 | 0.753 ± 0.036 | 0.266 (0.135–0.397) |
Значения попарных генетических различий и дистанций, рассчитанных на основании значений FST и Jost’s D, обобщены на рис. 1.
Рис. 1.
Генетические дистанции между исследуемыми популяциями тувинских овец. Попарные генетические дистанции по значениям Jost’s D показаны над диагональю; по FST – под диагональю. Расшифровку аббревиатур для популяций овец см. в табл. 1.

Как показано на рис. 2 и 3, тувинские овцы из разных хозяйств и районов республики сформировали два кластера, один из которых образован популяциями Oorug, Bai-Hol, Choduraa, Yаmaalyk, Aylyg-Oy, Malchyn, Murgustug, Mandala, а другой объединил популяции Adargan, Alaak, Bai-Tal, Mogen-Buren, Sukpak, Duza, Amyk, Beldir, Sai-Khonash, Torgalyg, Mongush, Despen, Ak-Khaya, Irtysh, что соответствует внутрипородному подразделению тувинской короткожирнохвостой овцы на два селекционно-заводских типа: горный и степной. Некоторые расхождения можно отметить лишь для популяции овец Dag-Uzu, которая в одном случае попадает в кластер животных горного типа (рис. 3), а в другом – в группу овец степного типа, являясь, в то же время, более близкой по генетическим расстояниям овцам горного типа (рис. 2).
Рис. 2.
Neighbor-net дендрограмма взаимоотношений исследуемых популяций овец тувинской короткожирнохвостой породы, построенная на основе матрицы попарных генетических дистанций FST.

Рис. 3.
Neighbor-net дендрограмма взаимоотношений исследуемых популяций овец тувинской короткожирнохвостой породы, построенная на основе матрицы попарных генетических дистанций Jost’s D.
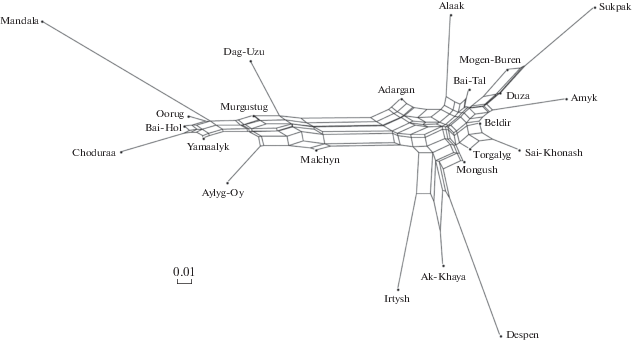
Причем, если на дендрограмме Neighbor-net, построенной на основе матрицы FST (рис. 2), граница между кластерами сравнительно нечеткая, то на дендрограмме на основе матрицы попарных генетических дистанций Jost’s D (рис. 3) градация рассматриваемых популяций на две большие группы более выражена.
При этом в кластере, объединяющем животных горного типа, наиболее генетически дивергентными оказались популяции Mandala, Choduraa, Aylyg-Oy (рис. 2, 3), Dag-Uzu (рис. 3), которые сформировали относительно длинные ветви, в то время как остальные популяции этой группы располагались на ребрах сети, что указывает на их смешанное происхождение (рис. 2, 3). Во втором кластере, включающем овец степного типа, сильная дивергенция была характерна для популяций Sukpak, Alaak, Amyk, Sai-Khonash, Despen, Irtysh (рис. 2, 3), Adargan, Mogen-Buren (рис. 2) и Ak-Khaya (рис. 3).
При этом генетические связи между популяциями Malchyn (ГУП РТ “Малчын” – хозяйство-оригинатор тувинских овец горного типа) и Despen (МУП “Племенное овцеводческое хозяйство “Деспен” – хозяйство-оригинатор тувинских овец степного типа) (FST = 0.108, Jost’s D = 0.3), относящимися к разным группам кластеризации, оказались на уровне генетических дистанций между наиболее удаленными популяциями Malchyn и Mandala (FST = 0.059, Jost’s D = 0.247) внутри кластера “овцы горного типа” и популяциями Despen и Sukpak (FST = 0.109, Jost’s D = 0.294) внутри кластера “овцы степного типа”. И хотя в рассматриваемых группах овец отмечается достаточно слабая или средняя дифференциация популяций, в целом межпопуляционные различия в группе тувинских овец “степного типа” (FST = = 0.013–0.153, Jost’s D = 0.029–0.461) оказались несколько бóльшими, чем в группе “горного типа” (FST = 0.01–0.071, Jost’s D = 0.029–0.247) (рис. 1), что отчасти подтверждается и в ходе анализа генетической структуры популяций тувинских короткожирнохвостых овец (рис. 4).
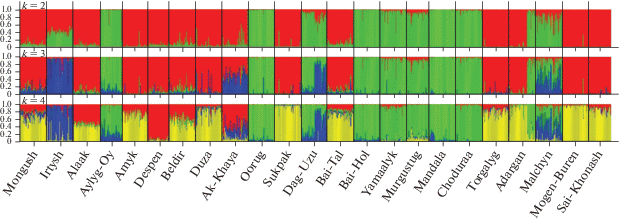
Так, если при k = 2 можно видеть разделение популяций тувинских овец на две группы, соответствующие горному и степному типам, то при k = 3 и k = 4 популяция Irtysh, относящаяся согласно Государственному племенному регистру (2017) [36] к степному типу тувинских короткожирнохвостых овец, формирует дополнительный кластер с неполной адмиксией.
ОБСУЖДЕНИЕ
Республика Тыва расположена в переходной зоне от центрально-азиатских пустынь к сибирской тайге и на ее территории выделяют четыре природно-экономических сельскохозяйственных района: центральный, или подтаежно-степной с лесостепной подзоной (Пий-Хемский, Кызылский, Улуг-Хемский, Чеди-Хольский кожууны); западный – горно-степной (Монгун-Тайгинский, Бай-Тайгинский, Барун-Хемчикский, Сут-Хольский, Дзун-Хемчикский, Чаа-Хольский, Овюрский кожууны), южный – сухих степей (Эрзинский, Тес-Хемский, Тандинский кожууны) и восточный – горно-таежный (Тоджинский, Тере-Хольский, Каа-Хемский кожууны) [37].
Проведенный по STR-маркерам молекулярно-генетический анализ выявил наличие двух групп тувинских овец (рис. 2, 3), которые объединяют, соответственно, животных горного и степного типов. При этом группа “горного типа” включала популяции тувинских овец Монгун-Тайгинского, Дзун-Хемчикского, Тес-Хемского, Бай-Тайгинского, Эрзинского и Овюрского кожуунов, а группу “степного типа” сформировали популяции тувинских овец Дзун-Хемчикского, Кызылского, Монгун-Тайгинского, Бай-Тайгинского, Барун-Хемчикского, Сут-Хольского, Овюрского и Тес-Хемского кожуунов, т.е. как минимум в трех районах: Монгун-Тайгинском, Дзун-Хемчикском и Тес-Хемском кожуунах встречаются животные обоих типов.
Следовательно, наблюдается некоторое несоответствие между распределением отдельных исследованных популяций тувинских овец горного и степного типов и зональной принадлежностью административных районов.
Выявленное расхождение, по-видимому, объясняется особенностями формирования современных типов тувинской грубошерстной овцы, определяющими факторами которых явились направление селекции и уровень обеспечения питательными веществами при пастбищном содержании.
В частности, горный тип тувинских овец был получен в ГУП РТ “Малчын” Монгун-Тайгинского кожууна путем многоступенчатого отбора из местных популяций [17, 38]. Животные этого типа консолидированы по экстерьерным особенностям, окраске руна, кроющего волоса и приспособлены к круглогодовому пастбищному содержанию в горно-степной зоне республики, где недостаточное увлажнение (100–187 мм) сочетается с пониженным температурным фоном (среднегодовая температура –2.5°С), безморозный период длится в среднем 95 дней, а пастбищная растительность изрежена и до 30% состоит из кустарников [17]. На одну овцу в год здесь приходится в 2 раза меньше заготовленных кормов, чем в хозяйствах степной зоны [10], что негативно сказывается на их живой массе и мясной продуктивности.
По сравнению с горно-степными районами зона сухих степей отличается менее жесткими природно-климатическими условиями. При средней температуре –1.5°С среднегодовое количество осадков составляет 180–230 мм, безморозный период длится 105–120 дней, травостой пастбищ – осоко-злаковый. Соответственно, при выведении тувинских овец степного типа применяли совершенно иную стратегию, чем при создании тувинских короткожирнохвостых овец горного типа. Для этого в хозяйствах ООО “Сельскохозяйственная корпорация “Кызылская” (Кызылский кожуун) и МУП “Племенное овцеводческое хозяйство “Деспен” (Тес-Хемский кожуун) путем скрещивания тувинских грубошерстных овцематок с баранами баядской короткожирнохвостой породы сначала получали помесей и лишь затем проводили отбор помесных животных с последующим закреплением полезных признаков в потомстве путем однородного подбора [17, 38].
И хотя тувинские овцы степного типа сходны по окраске с животными горного типа, по экстерьерным особенностям они имеют много общего с монгольскими овцами Говь-алтая и дархадскими [39]. Овцы степного типа лучше приспособлены к использованию равнинных сухостепных пастбищ с относительно высокой продуктивностью травостоя и проявляют потенциал мясной улучшающей породы [14]. Так, в отличие от овец горного типа овцематки и бараны-производители степного типа на 16.85 и 25.3 кг имеют бóльшую живую массу и на 0.4 и 0.1 кг дают больше шерсти [40, 41].
Разведением овец степного типа тувинской короткожирнохвостой породы кроме вышеназванных хозяйств-оригинаторов занимаются также СПК “Иртиш”, “Бай-Хол” и “Белдир”. Однако в нашем случае согласно построенным дендрограммам по FST и Jost’s D (рис. 2, 3) из этих трех хозяйств только популяции Irtysh и Beldir попадают в кластер “степного типа”, к тому же последующий структурный анализ выделяет популяцию Irtysh в самостоятельную группу, что в целом может свидетельствовать о неоднородной генетической дифференциации тувинских овец даже в пределах одного внутрипородного типа.
Важно отметить, что кроме увеличения поголовья новых типов в республике одновременно продолжается плановая селекционно-племенная работа по улучшению местных популяций тувинских грубошерстных овец. Главную роль в этом играют базовые овцеводческие хозяйства Тувинского НИИСХ: МУП “Племенное овцеводческое хозяйство “Деспен” (Тес-Хемский кожуун), СПК “Амык” (Сут-Хольский кожуун), СПК “Бай-Хол” (Эрзинский кожуун), “Бай-Тал” (Дзун-Хемчикский кожуун), ГУП РТ “Малчын” и ГУП РТ “Моген-Бурен” (Монгун-Тайгинский кожуун), МУП “Сай-Хонаш” (Барун-Хемчикский кожуун) [40]. Ю.А. Юлдашбаев отмечал, что наиболее ценное поголовье тувинских короткожирнохвостых овец находится в племенных репродукторах “Ак-Бедик” и “Кызыл-Тук” Овюрского кожууна республики [42].
В этом направлении задействован также потенциал крупных овцеводческих предприятий. Например, в ООО “Сельскохозяйственная корпорация “Кызылская” (Кызылский район) были получены три линии тувинской овцы, различающиеся между собой по одному или нескольким признакам [39]. Возможно, что частичной разнонаправленностью селекционно-племенных мероприятий и объясняется генетическое расхождение между отдельными популяциями тувинских овец, отмечаемое даже в границах одного административного района. Так, согласно результатам проведенных исследований в разные кластеры (рис. 2, 3) попали популяции Mongush и Irtysh (Кызылский кожуун) (FST = 0.051, Jost’s D = = 0.148), Dag-Uzu и Bai-Tal (Дзун-Хемчикский кожуун) (FST = 0.059, Jost’s D = 0.177), Malchyn и Mogen-Buren (Могун-Тайгинский кожуун) (FST = = 0.079, Jost’s D = 0.297).
Таким образом, типологический состав пастбищ и их продуктивность, обусловленные комплексом природно-климатических факторов, определяют территориальные особенности ведения селекционнно-племенной работы по тувинской грубошерстной короткожирнохвостой овце, которая на сегодняшний день представлена горным и степным внутрипородными типами, а также многочисленными местными популяциями, в большей или меньшей степени тяготеющими к тому или иному типу.
Исследования выполнены при финансовой поддержке РНФ в рамках проекта № 19-76-20061.
Все применимые международные, национальные и/или институциональные принципы ухода и использования животных были соблюдены.
Авторы заявляют, что у них нет конфликта интересов.
Список литературы
Донгак М.И. Продуктивность тувинских короткожирнохвостых овец с разным строением руна: Автореф. … канд. с.-х. наук. М., 2011. 20 с.
Григорян Л.Н., Хататаев С.А. Численность и племенная база полугрубошерстных и грубошерстных пород овец, разводимых в России // Овцы, козы, шерстяное дело. 2015. № 1. С. 9–12.
Монгуш С.С. Сравнительная оценка настрига и свойств шерсти тувинско-сараджинских полугрубошерстных и местных тувинских овец // Овцы, козы, шерстяное дело. 2018. № 1. С. 24–26.
Чысыма Р.Б., Кузьмина Е.Е. Инновационное развитие животноводства Республики Тыва // Природные ресурсы, среда и общество. 2019. Т. 1. № 1. С. 59–65.
Цырендондоков Н.Д. Основы овцеводства: биологические особенности овец. Технология производства. М.: Росагропромиздат, 1989. 175 с.
Кызыл-оол И.Т. Овцеводство Тувинской АССР: историко-зоотехнический очерк. Кызыл: Тувкнигоиздат, 1975. 263 с.
Варварин Б.Г. Пастбища и сенокосы Тувинской автономной области // Тр. Тувинской с.-х. опытной станции. Кызыл, 1950. Вып. 2. С. 7–85.
Лысенков А.А. Пастбища Тувы // Тр. Тувинской государственной с.-х. опытной станции. Кызыл, 1969. Вып. 4. 83 с.
Самбуу А.Д., Титлянова А.А. Влияние социально-экономического развития Республики Тыва на ее растительный покров // Фундаментальные исследования. 2014. № 11. Ч. 3. С. 550–555.
Конгар Н.М. Условия и некоторые вопросы развития овцеводства в Туве. Кызыл: Новости Тувы, 1995. 96 с.
Иргит Р.Ш., Лущенко А.Е., Оюн С.М. Характеристика овцеводства Республики Тыва по породному составу и зонально-территориальному размещению поголовья овец // Успехи современного естествознания. 2011. № 5. С. 148–151.
Государственный доклад “О состоянии окружающей среды Республики Тыва в 2000 г.”. Кызыл: Гос. Ком. природных ресурсов по Республике Тыва, 2001. 26 с.
Ондар С.Н. Формирование шерстной и овчинно-шубной продуктивности тувинских короткожирнохвостых овец: Автореф. … канд. с.-х. наук. Дубровицы, 1997. 26 с.
Билтуев С.И., Матханова А.В., Монгуш Б.Б. Экологическая пластичность тувинских короткожирнохвостых овец // Зоотехния. 2010. Т. 20. № 3. С. 28–33.
Коротков В.И., Рогожников А.М. Сараджинская порода овец в Сибири // Овцеводство. 1974. № 4. С. 23.
Двалишвили В.Г., Шимит Л.Д. Овцеводство Тывы на рубеже веков // Материалы научно-произв. конф. “Состояние и задачи обеспечения устойчивого развития агропромышленного производства Республики Тыва на 2001–2005 гг.”. Новосибирск, 2002. С. 119–122.
Юлдашбаев Ю.А., Донгак М.И., Куликова К.А. Хозяйственно-полезные признаки у овец тувинской короткожирнохвостой породы и перспективы изучения полиморфизма генов // Изв. Санкт-Петербургского гос. аграрного ун-та. 2016. № 42. С. 141–148.
Столповский Ю.А., Шимиит Л.В., Кол Н.В. и др. Анализ генетической изменчивости и филогенетических связей у популяций тувинской короткожирнохвостой овцы с использованием ISSR-маркеров // С.-х. биология. 2009. № 6. С. 34–43.
Куликова К.С. Характеристика тувинской короткожирнохвостой породы овец по ДНК-маркерам продуктивно-биологических особенностей: Автореф. … канд. биол. наук. Москва, 2018. 21 с.
Deniskova T.E., Dotsev A.V., Selionova M.I. et al. Population structure and genetic diversity of 25 Russian sheep breeds based on whole-genome genotyping // Genet. Sel. Evol. 2018. V. 50. № 1. P. 29. https://doi.org/10.1186/s12711-018-0399-5
Deniskova T.E., Abdelmanova A.S., Dotsev A.V. et al. PSX-18 high-density genomic description of Russian native sheep breed of the Republic of Tyva // J. Animal Sci. 2020. V. 98. № S4. P. 453–454.
Arora R., Bhatia S., Mishra B.P., Joshi B.K. Population structure in Indian sheep ascertained using microsatellite information // Anim. Genet. 2011. V. 42. № 3. P. 242–250. https://doi.org/10.1111/j.1365-2052.2010.02147.x
Qwabe O.O., Köster E., Visser C. Genetic diversity and population structure of the endangered Namaqua Afrikaner sheep // Trop. Anim. Health. Prod. 2013. V. 45. № 2. P. 511–516. https://doi.org/10.1007/s11250-012-0250-x
Kunene N.W., Ceccobelli S., Di Lorenzo P. et al. Genetic diversity in four populations of Nguni (Zulu) sheep assessed by microsatellite analysis // Ital. J. Animal Sci. 2014. V. 13. № 1. P. 75–82. https://doi.org/10.4081/ijas.2014.3083
Gahlot K., Purva M., Maherchandani S., Kashyap S.K. Original research assessment of genetic diversity in Chokla sheep breed of Rajasthan using microsatellite markers // Intern. J. Livestock Research. 2018. V. 8. № 11. P. 291–299. https://doi.org/10.5455/ijlr.20171227075019
Lowrey B., Proffitt K.M., McWhirter D.E., White P.J. Characterizing population and individual migration patterns among native and restored bighorn sheep (Ovis canadensis) // Ecol. Evol. 2019. V. 9. № 4. P. 8829–8839. https://doi.org/10.1002/ece3.5435
Nei M. Estimation of average heterozygosity and genetic distance from small number of individuals // Genetics. 1978. V. 89. № 3. P. 583–590.
Kalinowski S.T. Counting alleles with rarefaction: Private alleles and hierarchical sampling designs // Conserv. Genet. 2004. № 5. P. 539–543.https://doi.org/10.1023/B:COGE.0000041021.91777.1a
Weir B.S., Cockerham C.C. Estimating F-statistics for the analysis of population structure // Evolution. 1984. V. 38. № 6. P. 1358–1370. https://doi.org/10.2307/2408641
Jost L. GST and its relatives do not measure differentiation // Mol. Ecol. 2008. № 17. P. 4015–4026. https://doi.org/10.1111/j.1365-294X.2008.03887.x
Huson D.H., Bryant D. Application of phylogenetic networks in evolutionary studies // Mol. Biol. Evol. 2006. V. 23. № 2. P. 254–267. https://doi.org/10.1093/molbev/msj030
R Core Team. R: A language and environment for statistical computing. R Foundation for Statistical Computing. Vienna. 2018. https://www.R-project.org.
Pritchard J.K., Stephens M., Donnelly P. Inference of population structure using multilocus genotype data // Genetics. 2000. V. 155. № 2. P. 945–959. https://doi.org/10.1093/genetics/155.2.945
Peakall R., Smouse P.E. GenAlEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research – an update // Bioinformatics. 2012. V. 28. № 19. P. 2537–2539. https://doi.org/10.1093/bioinformatics/bts460
Keenan K., McGinnity P., Cross T.F. et al. DiveRsity: An R package for the estimation and exploration of population genetics parameters and their associated errors // Methods Ecol. Evol. 2013. № 4. P. 782–788. https://doi.org/10.1111/2041-210X.12067
Государственный племенной регистр (по состоянию на 02.02.2021) www.mcx.gov.ru.
Андрейчик М.Ф., Калугина Г.В. Особенности синоптических условий загрязнения Республики Тывы. Кызыл: Изд-во ТывГУ, 2002. 70 с.
Амерханов X.А., Билтуев C.И., Орус-оол В.С., Ооржак Ч.М. Особенности селекции тувинских короткожирнохвостых овец в разных природно-климатических условиях Республики Тыва // Овцы, козы, шерстяное дело. 2010. № 2. С. 8–13.
Монгуш Ж.Н. Продуктивные качества овец степного типа тувинской короткожирнохвостой породы в зависимости от их линейной принадлежности: Дис. … канд. с.-х. наук. Улан-Удэ, 2014. 113 с.
Монгуш Б.Б. Сравнительная характеристика экстерьерно-продуктивных и некоторых биологических особенностей тувинских короткожирнохвостых овец рвзных типов: Автореф. … канд. с.-х. наук. Улан-Удэ, 2012. 21 с.
Чысыма Р.Б., Кузьмина Е.Е. Инновационное развитие животноводства Республики Тыва // Природные ресурсы, среда и общество. 2019. С. 59–65.
Юлдашбаев Ю.А., Биче-оол С.Х. Технологическое проектирование овцеводческих крестьянско-фермерских хозяйств. М.: РГАУ-МСХА, 2009. 26 с.
Дополнительные материалы отсутствуют.