

Генетика, 2022, T. 58, № 3, стр. 267-298
Структура полиморфизма гордеинов, контролируемых аллелями локусов Hrd A и Hrd B, в диком ячмене (Hordeum spontaneum C. Koch)
А. А. Поморцев 1, *, А. В. Рубанович 1, Е. В. Лялина 1
1 Институт общей генетики им. Н.И. Вавилова Российской академии наук
119991 Москва, Россия
* E-mail: pomortsev@vigg.ru
Поступила в редакцию 15.06.2021
После доработки 21.07.2021
Принята к публикации 21.07.2021
- EDN: AVNQLO
- DOI: 10.31857/S0016675822030110
Аннотация
Методом электрофореза в крахмальном геле изучен полиморфизм гордеинов, контролируемых аллелями локусов Hrd A и Hrd B, в 258 образцах H. spontaneum из 18 стран Северной Африки, Ближнего Востока и Азии. В результате было обнаружено 225 вариантов блоков компонентов, контролируемых аллелями локуса Hrd A, которые были сгруппированы в 16 семейств. Среди вариантов HRD A, найденных в H. spontaneum, 33 (14.7%) ранее были выявлены и в H. vulgare. В диком ячмене было обнаружено 308 вариантов, контролируемых аллелями локуса Hrd B, которые были сгруппированы в 18 семейств блоков компонентов гордеина. Среди вариантов HRD B также 33 (10.7%) варианта ранее были идентифицированы в культурном ячмене. Семейства существенно различаются по числу составляющих их вариантов блоков: для HRD A – от 4 (AVII) до 48 (AI), для HRD B – от 5 (BXI, BXIV) до 49 (BXIII). Показана мозаичность и неравномерность частот в распространении семейств вариантов HRD А и HRD B в исследованных образцах H. spontaneum. Обнаружено, что самое большое разнообразие вариантов блоков компонентов в целом и самое большое число “культурных” вариантов HRD А и HRD B присутствуют в популяциях H. spontaneum из Израиля. При этом в израильских образцах дикого ячменя были представлены все семейства HRD A, а также все семейства HRD B за исключением одного. С учетом этих данных и данных литературы по геномному анализу, исследованию экзома и анализу нуклеотидных последовательностей генов Btr1/btr1 и Btr2/btr2 у дикого и культурного ячменя сделано предположение, что самое первое введение ячменя в культуру произошло на севере Израиля.
Ранее нами были проведены исследования структуры полиморфизма электрофоретических вариантов блоков компонентов гордеина в 1197 образцах стародавних сортов ячменя из 22 стран, входящих в вавиловские центры разнообразия и граничащих с ними, а также 1047 селекционных сортов из стран Европы, Азии, Америки и Африки [1]. По гордеинам, контролируемым аллелями локуса Hrd A, было выявлено 156 вариантов блоков компонентов, локуса Hrd B – 271 вариант и локуса Hrd F – 5 вариантов. Основываясь на данных литературы о молекулярных механизмах формирования полиморфизма гордеинов, исследовании искусственных мутантов по гордеин-кодирующим локусам [1–7], среди вариантов гордеинов А и В были выделены дискретные группы фенотипически схожих блоков компонентов – семейства блоков компонентов. Для HRD А было выделено 12 семейств, для HRD B – 17 семейств, которые существенно различались по числу составляющих их вариантов блоков – от 3 до 60. Было сделано предположение, что появление собственно семейств HRD A и HRD B обусловлено, с одной стороны, спонтанной гибридизацией между H. vulgare и H. spontaneum при распространении культурного ячменя, с другой стороны, накоплением мутаций в гордеин-кодирующих локусах. Цель настоящей работы – изучение структуры полиморфизма гордеинов А и В в H. spontaneum и ее сравнение со структурой полиморфизма этих локусов в культурном ячмене.
МАТЕРИАЛЫ И МЕТОДЫ
Разнообразие запасных белков, контролируемых локусами Hrd A и Hrd В в H. spontaneum, изучали путем электрофоретического анализа гордеинов в 258 образцах из 18 стан Северной Африки, Ближнего Востока и Азии (табл. 1). Территории этих стран входят в ареал H. spontaneum [8]. Материал для исследований был получен из Генбанков: National Small Grains Research Collection USDA (Абердин, Айдахо, США), International Center for Agricultural Research in the Dry Areas – ICARDA (Алеппо, Сирия), Leibniz Institute of Plant Genetics and Crop Plant Research – (IPK) (Гатерслебен, Германия). Часть материала была собрана и предоставлена доктором H. Ozkan, Department of Field Crops, Faculty of Agriculture, University of Çukurova, Adana, Турция. Популяции дикого ячменя из Израиля представляли собой оригинальные сборы в основном из районов Галилейского моря (оз. Кинерет), а также Голанских высот и Хайфы. Материал был собран О. Раскиной, А. Беляевым и Е. Бадаевой в 2000–2008 гг. При работе с коллекционным материалом были обнаружены образцы, которые, по нашему мнению, нельзя отнести к H. spontantum. Например образец IG 38709 из Палестины был представлен не колосками, характерными для H. spontaneum, у которых при разламывании колоса при созревании остаются членики колосового стержня (рис. 1,а), а индивидуальными, хорошо выполненными зерновками, не сросшимися с члениками колосового стержня (рис. 1,б). Кроме этого, при электрофоретическом анализе гордеина в некоторых образцах дикого ячменя из Ирана была установлена их гетерозиготность по аллелям гордеин-кодирующих локусов, которые встречались как в H. vulgare, так и в H. spontaneum [9]. Не исключено, что это спонтанные гибриды между диким и культурным ячменем, возникшие при репродуцировании коллекционного материала. Такие образцы из исследований были исключены. Для каждого образца дикого ячменя анализировали гордеины в 5–10 индивидуальных зерновках.
Таблица 1.
Число исследованных образцов H. spontaneum из различных стран
| № п/п | Страна | Число образцов | № п/п | Страна | Число образцов |
|---|---|---|---|---|---|
| 1 | Ливия | 7 | 10 | Сирия | 23 |
| 2 | Египет | 1 | 11 | Иордания | 34 |
| 3 | Кипр | 3 | 12 | Ирак | 21 |
| 4 | Армения | 1 | 13 | Западный Иран | 43 |
| 5 | Азербайджан | 2 | 14 | Туркменистан | 18 |
| 6 | Юго-Восточная Турция | 37 | 15 | Узбекистан | 4 |
| 7 | Ливан | 10 | 16 | Таджикистан | 2 |
| 8 | Израиль | 30 | 17 | Афганистан | 3 |
| 9 | Палестина | 17 | 18 | Пакистан | 2 |
| Всего | 258 | ||||
Рис. 1.
а – колосок образца H. spontaneum IG 38677, б – зерновки, представляющие образец H. spontaneum IG 38709.

Электрофорез гордеинов проводили в столбиках 12–14%-ного крахмального геля с 3 М мочевиной в алюминий-лактатном буфере с рН 3.1 по методике А.А. Созинова и Ф.А. Поперели [10] с некоторыми модификациями [11]. Анализ сходства исследованных популяций H. spontaneum проводили методом главных компонент с помощью пакета программ SPSS 19.
РЕЗУЛЬТАТЫ
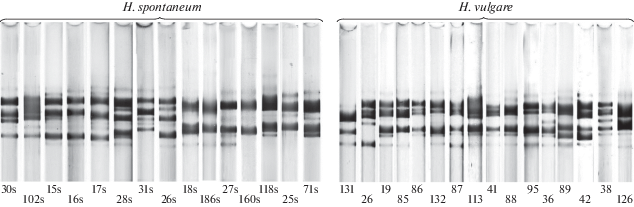
В результате электрофоретического анализа гордеина в 258 образцах H. spontantum было обнаружено 225 вариантов блоков компонентов, контролируемых аллелями локуса Hrd A, из которых 33 (14.7%) ранее были выявлены и в культурном ячмене (рис. 2, табл. 2). Из 308 идентифицированных в диком ячмене вариантов блоков компонентов, контролируемых аллелями локуса Hrd B, так же 33 (10.7%) варианта ранее были обнаружены в H. vulgare (рис. 3, табл. 2).
Рис. 2.
Варианты блоков компонентов гордеина, контролируемые аллелями локуса Hrd A, обнаруженные при анализе 258 образцов H. spontaneum. Символом “s” обозначены варианты блоков компонентов гордеина, обнаруженные исключительно в H. spontaneum.

Таблица 2.
Число вариантов блоков компонентов гордеинов А и В, объединяемых в различные семейства в H. spontaneum
| Число вариантов блоков компонентов гордеина А в семействах | |||||
|---|---|---|---|---|---|
| Семейство | Число вариантов | Семейство | Число вариантов | Семейство | Число вариантов |
| AI | 39/9 | AVIII | 15/0 | AXIV | 8/1 |
| AIII | 1/2 | AIX | 13/3 | AXV | 10/0 |
| AIV | 15/5 | AX | 8/0 | AXVI | 13/0 |
| AV | 25/5 | AXI | 3/2 | AXVII | 9/0 |
| AVI | 4/1 | AXII | 10/3 | ||
| AVII | 3/1 | AXIII | 16/1 | ||
| Всего | 192/33 | ||||
| Число вариантов блоков компонентов гордеина В в семействах | |||||
| BI | 27/1 | BVII | 18/0 | BXIII | 39/10 |
| BII | 14/3 | BVIII | 15/2 | BXIV | 5/0 |
| BIII | 10/2 | BIX | 20/2 | BXV | 14/1 |
| BIV | 21/5 | BX | 9/0 | BXVII | 8/2 |
| BV | 19/1 | BXI | 5/0 | BXVIII | 6/0 |
| BVI | 5/3 | BXII | 20/1 | BXIX | 20/0 |
| Всего | 275/33 | ||||
Рис. 3.
Варианты блоков компонентов гордеина, контролируемые аллелями локуса Hrd В, обнаруженные при анализе 258 образцов H. spontaneum. Символом “s” обозначены варианты блоков компонентов гордеина, обнаруженные исключительно в H. spontaneum.

По гордеину А в диком ячмене по аналогии с культурным ячменем мы выделили 16 групп фенотипически схожих вариантов блоков компонентов – семейств вариантов блоков компонентов (рис. 2, табл. 2). Ранее в культурном ячмене нами были выделены 12 семейств и один вариант HRD A21, который не относился ни к одному из семейств [1]. Сравнивая структуру полиморфизма гордеина А в диком и в культурном ячмене, в H. spontaneum мы обнаружили 11 семейств, включающих как варианты, характерные только для дикого ячменя, так и варианты, выявленные ранее в культурном ячмене. К семейству, обозначенному нами в H. spontaneum как AXIII (включающее 17 вариантов), относится вариант HRD A21 не вошедший ни в одно из семейств в H. vulgare (рис. 2). Кроме этого, в диком ячмене мы выделили отдельное семейство AXIV, в которое вошел вариант HRD A62. Этот вариант блока компонентов гордеина в культурном ячмене относили ранее к семейству AI [1]. Однако выявление в H. spontaneum восьми близких с HRD A62 вариантов позволило объединить их в отдельное семейство (рис. 2). Представители семейств AXV–AXVII, выделенные в диком ячмене, ранее в культурном ячмене обнаружены не были. В исследованных образцах H. spontaneum мы не выявили варианты гордеина А, которые можно было бы отнести к семейству AII, представленному в H. vulgare пятью вариантами [1]. Напротив, в семействе AX в диком ячмене мы не обнаружили варианты, идентичные с вариантами, вошедшими в это семейство в культурном ячмене (рис. 4, табл. 2). В образцах H. spontaneum наибольшее число вариантов включало семейство AI – 48 (21.3%) (табл. 2). К самым многочисленным семействам в диком ячмене относились также AV – 30 (13.3%), AIV – 20 (8.9%), AXIII – 17 (7.6%), AIX – 16 (7.1%) и AVIII – 15 (6.7%). Остальные десять семейств объединяли от трех (AIII) до тринадцати (AXII) вариантов. Таким образом, шесть семейств из 16 включали 64.9% вариантов блоков компонентов гордеина А, обнаруженных в диком ячмене. Заметим, что в культурном ячмене семейства AI, AV и AVIII также объединяли самое большое число вариантов блоков компонентов гордеина А: 60 (38.5%) и по 16 (10.3%) соответственно [1].
Рис. 4.
Варианты блоков компонентов, контролируемые аллелями локуса Hrd A, объединенные в семейство AVIII в H. spontaneum и H. vulgare.
В H. spontaneum 308 вариантов гордеина В были сгруппированы в 18 семейств (рис. 3, табл. 2). При этом в 12 семейств вошли варианты блоков компонентов, обнаруженные как в диком, так и в культурном ячмене. В то же время, в семействах BVII, BX, BXI и BXIV отсутствовали общие для H. spontaneum и H. vulgare варианты блоков компонентов гордеина. В диком ячмене нам не встретились варианты гордеина В, которые можно было бы отнести в семейство BXVI. В культурном ячмене в это семейство входили три варианта [1]. В H. spontaneum были выделены семейства BXVIII и BXIX, представителей которых в культурном ячмене мы не обнаружили. Наибольшее число вариантов блоков компонентов гордеина В включали семейства: BXIII – 49 (15.9%), BI – 28 (9.1%), BIV – 26 (8.4%), BIX – 22 (7.1%), BXII – 21 (6.8%), BV и BXIX – по 20 (6.5%). В состав остальных 11 семейств входили от 5 (ВXI) до 18 (BVII) вариантов гордеина В. Таким образом, семь семейств из 18 объединяют 60.3% вариантов блоков компонентов гордеина В, идентифицированных в H. spontaneum. В культурном ячмене семейства BXIII, BIV, BIX и BXII также включали наибольшее число вариантов блоков компонентов: BXIII – 41 (15.1%), BIV – 26 (9.6%), BIX – 19 (7.8%), BXII – 24 (8.9%) [1].
Присутствие вариантов блоков компонентов, входящих в семейства HRD А и HRD В в исследованных образцах дикого ячменя из различных стран оказалось неодинаковым. Так, в популяциях H. spontaneum из Израиля были обнаружены варианты гордеина А, представляющие все 16 семейств (табл. 3). В сирийских образцах были выявлены представители 12 семейств, в иранских – 11, а в Турецких – 10 семейств. В исследованных популяциях из других стран присутствовали представители от одного (Египет, Азербайджан) до девяти (Иордания) семейств гордеина А. Аналогичная картина наблюдалась и для семейств гордеина В (табл. 3). В израильских образцах дикого ячменя были обнаружены представители 17 семейств, за исключением BXIV. Варианты гордеина В, вошедшие в это семейство, присутствовали только в популяциях из Турции и Ирана с суммарными частотами 0.0359 и 0.0932 соответственно (табл. 3). Относительно широко семейства гордеина В были представлены в H. spontaneum из Турции, Сирии и Ирана (по 14 семейств), а также из Ирака (13 семейств), Палестины (11 семейств) и Иордании (10 семейств). В образцах дикого ячменя из других стран были найдены варианты, относящиеся к от одного (Египет, Армения) до семи (Туркменистан) семействам гордеина В (табл. 3).
Таблица 3.
Суммарные частоты вариантов блоков компонентов семейств HRD A и HRD B в H. spontaneum из стран Северной Африки, Ближнего Востока и Азии
| Семейство | Страна | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ливия | Египет | Кипр | Армения | Азербайджан | Турция | Ливан | Израиль | Палестина | Сирия | Иордания | Ирак | Иран | Туркменистан | Узбекистан | Таджикистан | Афганистан | Пакистан | |
| AI | 0.3429 | 1 | 0 | 0 | 0 | 0.1486 | 0.24 | 0.1138 | 0.2236 | 0.2646 | 0.2316 | 0 | 0.0407 | 0.1889 | 0 | 0 | 0 | 0 |
| AIII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0179 | 0 | 0 | 0 | 0 | 0.0233 | 0 | 0 | 0 | 0 | 0.1 |
| AIV | 0.0286 | 0 | 0 | 0 | 0 | 0.0495 | 0 | 0.034 | 0 | 0.0696 | 0.0526 | 0 | 0.0851 | 0 | 0 | 0 | 0 | 0.6 |
| AV | 0.2286 | 0 | 0.3333 | 0 | 0 | 0.1443 | 0.1 | 0.2864 | 0.0588 | 0.1787 | 0.1579 | 0.16 | 0.2981 | 0.0777 | 0 | 0.3 | 0 | 0 |
| AVI | 0 | 0 | 0 | 0 | 0 | 0 | 0.12 | 0.008 | 0 | 0.0261 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| AVII | 0.0857 | 0 | 0 | 0 | 0 | 0 | 0 | 0.016 | 0 | 0.1913 | 0.1789 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| AVIII | 0 | 0 | 0 | 0.2 | 0 | 0.1394 | 0.08 | 0.0895 | 0.0706 | 0.0261 | 0.0527 | 0.24 | 0.1222 | 0.1112 | 0 | 0 | 0 | 0 |
| AIX | 0.2571 | 0 | 0 | 0 | 0 | 0.027 | 0 | 0.1194 | 0 | 0.0174 | 0.0631 | 0.05 | 0.0931 | 0.1889 | 0.4 | 0 | 0.1334 | 0.1 |
| AX | 0 | 0 | 0 | 0 | 0 | 0.0135 | 0 | 0.01 | 0 | 0.0087 | 0.0421 | 0.05 | 0 | 0.1333 | 0 | 0 | 0 | 0 |
| AXI | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.006 | 0.1176 | 0 | 0 | 0 | 0.1164 | 0.0556 | 0 | 0 | 0.3333 | 0 |
| AXII | 0 | 0 | 0 | 0.6 | 1 | 0.3066 | 0 | 0.0779 | 0.1883 | 0.0957 | 0.1685 | 0.2 | 0.0931 | 0.1444 | 0.05 | 0 | 0.2 | 0.2 |
| AXIII | 0 | 0 | 0 | 0.2 | 0 | 0.09 | 0.1 | 0.0339 | 0.1411 | 0.0522 | 0.0526 | 0.08 | 0.0232 | 0 | 0 | 0 | 0 | 0 |
| AXIV | 0.0571 | 0 | 0 | 0 | 0 | 0 | 0.04 | 0.0775 | 0.1647 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| AXV | 0 | 0 | 0 | 0 | 0 | 0.027 | 0 | 0.022 | 0 | 0 | 0 | 0 | 0.0815 | 0 | 0 | 0 | 0 | 0 |
| AXVI | 0 | 0 | 0 | 0 | 0 | 0 | 0.32 | 0.0459 | 0.0353 | 0.0435 | 0 | 0.03 | 0 | 0 | 0 | 0 | 0 | 0 |
| AXVII | 0 | 0 | 0.6667 | 0 | 0 | 0.0541 | 0 | 0.0418 | 0 | 0.0261 | 0 | 0.19 | 0.0233 | 0.1 | 0.55 | 0.7 | 0.3333 | 0 |
| BI | 0.2857 | 0 | 0.3333 | 0 | 0 | 0.036 | 0.06 | 0.0419 | 0.0588 | 0.2684 | 0.2843 | 0 | 0.031 | 0.0235 | 0 | 0 | 0 | 0 |
| BII | 0 | 0 | 0.2667 | 0 | 0 | 0.036 | 0 | 0.0418 | 0.0588 | 0.0174 | 0 | 0.04 | 0.0154 | 0 | 0.25 | 0 | 0.3333 | 0 |
| BIII | 0 | 0 | 0.4 | 0 | 0 | 0.0313 | 0 | 0.0598 | 0 | 0 | 0.0316 | 0 | 0.0349 | 0.2235 | 0 | 0 | 0 | 0 |
| BIV | 0.2571 | 0 | 0 | 0 | 0 | 0.1376 | 0.1 | 0.2054 | 0 | 0.0609 | 0.0315 | 0.11 | 0.0581 | 0 | 0 | 0 | 0 | 0 |
| BV | 0 | 0 | 0 | 0 | 0 | 0.0676 | 0.08 | 0.1153 | 0.0353 | 0.1391 | 0 | 0.04 | 0.1125 | 0.1529 | 0 | 0 | 0 | 0 |
| BVI | 0 | 0 | 0 | 0 | 0 | 0.0089 | 0 | 0.018 | 0.0706 | 0.0174 | 0 | 0.13 | 0 | 0 | 0 | 0 | 0 | 0 |
| BVII | 0 | 0 | 0 | 0 | 0.25 | 0.0676 | 0.1 | 0.0339 | 0 | 0.0435 | 0.0105 | 0.06 | 0.0582 | 0.2471 | 0 | 0 | 0 | 0 |
| BVIII | 0.0572 | 0 | 0 | 0 | 0 | 0.0834 | 0 | 0.1096 | 0.0706 | 0.0187 | 0 | 0.03 | 0.0233 | 0 | 0 | 0 | 0.1334 | 0 |
| BIX | 0 | 0 | 0 | 0 | 0.45 | 0.0767 | 0 | 0.0358 | 0.0353 | 0.0521 | 0.0631 | 0.05 | 0.0929 | 0.1412 | 0 | 0.2 | 0 | 0 |
| BX | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.004 | 0 | 0.0087 | 0.179 | 0.01 | 0 | 0 | 0 | 0 | 0 | 0.3 |
| BXI | 0 | 0 | 0 | 0 | 0 | 0.0135 | 0 | 0.002 | 0.1177 | 0 | 0.0421 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| BXII | 0 | 0 | 0 | 0 | 0 | 0.0359 | 0 | 0.0299 | 0.2353 | 0.0261 | 0.0842 | 0.07 | 0 | 0 | 0 | 0 | 0 | 0 |
| BXIII | 0.4 | 1 | 0 | 1 | 0.3 | 0.2206 | 0.28 | 0.211 | 0.2353 | 0.1738 | 0.1263 | 0.04 | 0.0772 | 0 | 0 | 0 | 0.3333 | 0 |
| BXIV | 0 | 0 | 0 | 0 | 0 | 0.0359 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0932 | 0 | 0 | 0 | 0 | 0 |
| BXV | 0 | 0 | 0 | 0 | 0 | 0.149 | 0 | 0.014 | 0.0235 | 0.0174 | 0 | 0.17 | 0.0233 | 0 | 0 | 0 | 0 | 0 |
| BXVII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0338 | 0 | 0.0087 | 0 | 0.02 | 0.0772 | 0.0118 | 0 | 0 | 0 | 0 |
| BXVIII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0 | 0 | 0 | 0 | 0.1281 | 0 | 0 | 0 | 0 | 0 |
| BXIX | 0 | 0 | 0 | 0 | 0 | 0 | 0.38 | 0.0418 | 0.0588 | 0.1478 | 0.1474 | 0.23 | 0.1747 | 0.2 | 0.75 | 0.8 | 0.2 | 0.7 |
Распространение вариантов блоков компонентов, составляющих семейства гордеина А в различных странах также было неравномерным (табл. 3). В частности, варианты семейства AXII были обнаружены в 13 странах с суммарными частотами от 0.05 (Узбекистан) до 1.00 (Азербайджан). С относительно высокими суммарными частотами представители этого семейства встречались в H. spontaneum из Турции (0.3066), Ирака (0.2), Палестины (0.1883), Иордании (0.1685) и Туркменистана (0.1444). Варианты, составляющие семейство AV, обнаруженные в диком ячмене из 12 стран, с наибольшими частотами присутствовали в образцах из Кипра (0.3333), Ирана (0.2981), Израиля (0.2864), Ливии (0.2286). В популяциях дикого ячменя из 11 стран идентифицированы варианты гордеина А, объединенные в семейство AIX. Наибольшие суммарные частоты этого семейства наблюдались в образцах из Узбекистана (0.4), Ливии (0.2571), Туркменистана (0.1889), Афганистана (0.1334) и Израиля (0.1194). В образцах из десяти стран Ближнего Востока и Азии присутствовали варианты блоков компонентов, входящие в семейства: AI с частотами от 0.0407 (Иран) до 1.0 (Египет), семейство AVIII с частотами от 0.0261 (Иордания) до 0.24 (Ирак) и семейство AXVII с частотами от 0.0261 (Сирия) до 0.6667 (Кипр). Варианты блоков компонентов гордеина А, составляющие остальные десять семейств, были обнаружены с различными суммарными частотами в образцах H. spontaneum от трех до девяти стран.
Неравномерное распространение в образцах дикого ячменя из различных стран наблюдалось и для вариантов семейств гордеина В (табл. 3). Так, представители семейства BXIII присутствовали в H. spontaneum из 13 стран с суммарными частотами от 0.04 (Ирак) до 1.0 (Египет, Армения). Относительно широкое распространение имели также семейства: BXIX – в 12 странах с частотами от 0.0418 (Израиль) до 0.8 (Таджикистан), BI и BIX – в 10 странах с частотами от 0.0235 (Туркменистан) до 0.3333 (Кипр) и от 0.0353 (Палестина) до 0.45 (Азербайджан) соответственно. Представители остальных 14 семейств были обнаружены в образцах от двух (BXIV и BXVIII) до девяти (BII и BVII) стран с различными частотами (табл. 3).
Выше было отмечено, что в исследованных образцах H. spontaneum присутствовали варианты гордеинов А и В, идентичные обнаруженным ранее в H. vulgare (по 33 для HRD A и HRD B). В целом “культурные” варианты гордеина А были идентифицированы в образцах дикого ячменя из 14 стран и относились к 11 семействам (табл. 4). В H. spontaneum из 11 стран присутствовали блоки компонентов семейства AXII, контролируемые аллелями А22 (в семи странах), А97 (в пяти странах) и А98 (в четырех странах) с различными частотами (табл. 4, приложение). Заметим, что в культурном ячмене это семейство включает пять вариантов и обнаружено в образцах местного ячменя из девяти стран [1]. В образцах дикого ячменя из семи стран (Турции, Израиля, Палестины, Иордании, Ирака, Ирана, Туркменистана) с различными частотами присутствовали “культурные” варианты, контролируемые аллелями Hrd А1, А12, А18, А40, А93, относящиеся к семейству AV (табл. 4, приложение). В образцах местного культурного ячменя это семейство включает 16 вариантов и обнаружено в популяциях из 19 стран [1]. В H. spontaneum из Ливии, Турции, Израиля, Сирии, Иордании и Ирана с различными частотами присутствовали варианты блоков компонентов А16, А23, А28, А106 и А139, вошедшие в семейство AIV (табл. 4, приложение). В H. vulgare это семейство объединяет восемь вариантов, и его представители встречались в местных сортах из 13 стран [1]. Варианты блоков компонентов, контролируемые аллелями Hrd А2, А24, А63, А67, А110, А123, A144 и А115, представляющие семейство AI, с различными частотами выявлены в диком ячмене из Турции, Израиля, Сирии, Иордании и Ирана. При этом все указанные варианты, исключая А144, присутствовали в H. spontaneum из Израиля. Заметим, что вариант HRD A2 имеет самое широкое распространение в культурном ячмене. Он присутствовал с разными частотами в местных образцах H. vulgare из 21 страны от Марокко до Китая и Непала, за исключением Горного Бадахшана (Таджикистан). В популяциях из других стран обнаружены отдельные варианты, входящие в это семейство (табл. 4, приложение). Сравнительно большое число вариантов HRD A, выявленных ранее в культурном ячмене, присутствовало в образцах из Ирана (12), Иордании и Турции (по 7), Сирии и Ирака (по 6) (табл. 4). В целом “культурные” варианты гордеина А обнаружены главным образом в популяциях дикого ячменя от Юго-Восточной Турции до Западного Ирана, исключая Ливан. При этом самое большое число таких вариантов было выявлено в H. spon-taneum из Израиля – 18 (табл. 4, приложение).
Таблица 4.
Число вариантов блоков компонентов в семействах HRD А H. spontaneum из стран Северной Африки, Ближнего Востока и Азии
| Семейства HRD А | Страна | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ливия | Египет | Кипр | Армения | Азербайджан | Турция | Ливан | Израиль | Палестина | Сирия | Иордания | Ирак | Иран | Туркменистан | Узбекистан | Таджикистан | Афганистан | Пакистан | |
| I | 5/0 | 2/0 | 0/0 | 0/0 | 0/0 | 6/2 | 4/0 | 15/7 | 6/0 | 5/2 | 6/1 | 0/0 | 1/1 | 1/0 | 0/0 | 0/0 | 0/0 | 0/0 |
| III | 0/0 | 0/0 | 0/0 | 0/0 | 0/0 | 0/0 | 0/0 | 1/0 | 0/0 | 0/0 | 0/0 | 0/0 | 0/1 | 0/0 | 0/0 | 0/0 | 0/0 | 0/1 |
| IV | 0/1 | 0/0 | 0/0 | 0/0 | 0/0 | 1/1 | 0/0 | 9/2 | 0/0 | 1/1 | 2/1 | 0/0 | 2/3 | 0/0 | 0/0 | 0/0 | 0/0 | 1/0 |
| V | 2/0 | 0/0 | 1/0 | 0/0 | 0/0 | 2/1 | 1/0 | 17/4 | 0/1 | 2/0 | 3/1 | 2/2 | 2/2 | 2/1 | 0/0 | 1/0 | 0/0 | 0/0 |
| VI | 0/0 | 0/0 | 0/0 | 0/0 | 0/0 | 0/0 | 3/0 | 2/1 | 0/0 | 1/0 | 0/0 | 0/0 | 0/0 | 0/0 | 0/0 | 0/0 | 0/0 | 0/0 |
| VII | 1/0 | 0/0 | 0/0 | 0/0 | 0/0 | 0/0 | 0/0 | 2/1 | 0/0 | 3/0 | 2/0 | 0/0 | 0/0 | 0/0 | 0/0 | 0/0 | 0/0 | 0/0 |
| VIII | 0/0 | 0/0 | 0/0 | 1/0 | 0/0 | 6/0 | 2/0 | 6/0 | 2/0 | 2/0 | 2/0 | 2/0 | 4/0 | 2/0 | 0/0 | 0/0 | 0/0 | 0/0 |
| IX | 2/0 | 0/0 | 0/0 | 0/0 | 0/0 | 2/0 | 0/0 | 6/3 | 0/0 | 0/1 | 1/1 | 0/1 | 3/0 | 1/0 | 1/0 | 0/0 | 1/0 | 1/0 |
| X | 0/0 | 0/0 | 0/0 | 0/0 | 0/0 | 1/0 | 0/0 | 4/0 | 0/0 | 1/0 | 1/0 | 1/0 | 0/0 | 1/0 | 0/0 | 0/0 | 0/0 | 0/0 |
| XI | 0/0 | 0/0 | 0/0 | 0/0 | 0/0 | 0/0 | 0/0 | 3/0 | 0/1 | 0/0 | 0/0 | 0/0 | 1/2 | 0/1 | 0/0 | 0/0 | 0/1 | 0/0 |
| XII | 0/0 | 0/0 | 0/0 | 1/1 | 1/1 | 0/2 | 0/0 | 7/0 | 2/1 | 1/2 | 1/2 | 1/2 | 1/2 | 0/1 | 0/1 | 0/0 | 0/1 | 1/0 |
| XIII | 0/0 | 0/0 | 0/0 | 1/0 | 0/0 | 4/1 | 1/0 | 5/0 | 2/0 | 1/0 | 2/0 | 4/1 | 1/1 | 0/0 | 0/0 | 0/0 | 0/0 | 0/0 |
| XIV | 0/0 | 0/0 | 0/0 | 0/0 | 0/0 | 0/0 | 1/0 | 9/0 | 2/0 | 0/0 | 0/1 | 0/0 | 0/0 | 0/0 | 0/0 | 0/0 | 0/0 | 0/0 |
| XV | 0/0 | 0/0 | 0/0 | 0/0 | 0/0 | 1/0 | 0/0 | 3/0 | 0/0 | 0/0 | 0/0 | 0/0 | 4/0 | 0/0 | 0/0 | 0/0 | 0/0 | 0/0 |
| XVI | 1/0 | 0/0 | 0/0 | 0/0 | 0/0 | 0/0 | 4/0 | 4/0 | 1/0 | 3/0 | 0/0 | 2/0 | 0/0 | 0/0 | 0/0 | 0/0 | 0/0 | 0/0 |
| XVII | 0/0 | 0/0 | 2/0 | 0/0 | 0/0 | 2/0 | 0/0 | 4/0 | 0/0 | 2/0 | 0/0 | 3/0 | 1/0 | 1/0 | 1/0 | 1/0 | 1/0 | 0/0 |
| Всего | 11/1 | 2/0 | 3/0 | 3/1 | 1/1 | 25/7 | 16/0 | 97/18 | 15/3 | 22/6 | 20/7 | 15/6 | 20/12 | 8/3 | 2/1 | 2/0 | 2/2 | 3/1 |
Примечание. В табл. 4 и 5 в числителе – число вариантов блоков компонентов гордеинов, обнаруженных исключительно в H. spontaneum, в знаменателе – число вариантов блоков компонентов гордеинов, обнаруженных как в H. spontaneum, так и в H. vulgare.
Аналогичная картина наблюдается и в распространении вариантов блоков компонентов, контролируемых аллелями локуса Hrd B, идентифицированных ранее в H. vulgare. Так, “культурные” варианты гордеина В, относящиеся к 12 семействам, обнаружены в образцах H. spontaneum из десяти стран (табл. 5). В популяциях из восьми стран с различными частотами присутствовали блоки компонентов семейства BXIII, контролируемые аллелями: B12 (в Турции), В17 и В182 (в Иране), В29 и В162 (в Израиле), В82 (в Ливии), В141 (в Израиле и Ливии), В142 (в Израиле и Ираке), В145 (в Армении), В163 (в Израиле, Палестине и Ираке) (приложение). В местных образцах H. vulgare к этому семейству относится 41 вариант, эти варианты присутствуют в популяциях из 21 страны [1]. “Культурные” варианты, относящиеся к семейству BIV, идентифицированы в диком ячмене из пяти стран: в Израиле (В15, B16 и В65), в Иордании (В65), в Сирии и Иране (В118), а также в Ираке (В166) (приложение). В культурном ячмене к этому семейству относятся восемь вариантов, и его представители были найдены в образцах из 21-ой страны [1]. Единичные представители семейств BI, BII, BIII, BV, BVI, BVIII, BIX, BXII, BXV и BXVII с различными частотами были обнаружены в образцах H. spontaneum от одной до трех стран (приложение). В целом “культурные” варианты гордеина В, как и гордеина А, выявлены главным образом в популяциях дикого ячменя от Юго-Восточной Турции до Западного Ирана, исключая Ливан. При этом самое большое число таких вариантов было идентифицировано в H. spontaneum из Израиля – 15 (табл. 5, приложение).
Таблица 5.
Число вариантов блоков компонентов в семействах HRD B H. spontaneum в странах Северной Африки, Закавказья, Ближнего Востока и Азии
| Семейства HRD B | Страна | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ливия | Египет | Кипр | Армения | Азербайджан | Турция | Ливан | Израиль | Палестина | Сирия | Иордания | Ирак | Иран | Туркменистан | Узбекистан | Таджикистан | Афганистан | Пакистан | |
| I | 1/0 | 0/0 | 1/0 | 0/0 | 0/0 | 2/0 | 3/0 | 8/1 | 2/0 | 7/0 | 8/0 | 0/0 | 1/1 | 1/0 | 0/0 | 0/0 | 0/0 | 0/0 |
| II | 0/0 | 0/0 | 1/0 | 0/0 | 0/0 | 2/0 | 0/0 | 6/1 | 1/0 | 1/0 | 0/0 | 1/2 | 1/1 | 0/0 | 1/0 | 0/0 | 1/0 | 0/0 |
| III | 0/0 | 0/0 | 1/0 | 0/0 | 0/0 | 3/0 | 0/0 | 5/1 | 0/0 | 0/0 | 1/0 | 0/0 | 0/1 | 1/1 | 0/0 | 0/0 | 0/0 | 0/0 |
| IV | 1/0 | 0/0 | 0/0 | 0/0 | 0/0 | 5/0 | 1/0 | 8/3 | 0/0 | 1/1 | 2/1 | 1/1 | 3/1 | 0/0 | 0/0 | 0/0 | 0/0 | 0/0 |
| V | 0/0 | 0/0 | 0/0 | 0/0 | 0/0 | 4/0 | 1/0 | 13/1 | 1/0 | 5/1 | 0/0 | 1/0 | 2/1 | 2/0 | 0/0 | 0/0 | 0/0 | 0/0 |
| VI | 0/0 | 0/0 | 0/0 | 0/0 | 0/0 | 1/0 | 0/0 | 2/2 | 1/0 | 1/1 | 0/0 | 1/1 | 0/0 | 0/0 | 0/0 | 0/0 | 0/0 | 0/0 |
| VII | 0/0 | 0/0 | 0/0 | 0/0 | 2/0 | 4/0 | 1/0 | 6/0 | 0/0 | 1/0 | 1/0 | 1/0 | 3/0 | 1/0 | 0/0 | 0/0 | 0/0 | 0/0 |
| VIII | 2/0 | 0/0 | 0/0 | 0/0 | 0/0 | 4/1 | 0/0 | 5/0 | 0/1 | 2/0 | 0/0 | 2/0 | 1/0 | 0/0 | 0/0 | 0/0 | 1/0 | 0/0 |
| IX | 0/0 | 0/0 | 0/0 | 0/0 | 1/0 | 2/1 | 0/0 | 7/0 | 1/0 | 1/0 | 1/1 | 1/0 | 6/1 | 2/0 | 0/0 | 1/0 | 0/0 | 0/0 |
| X | 0/0 | 0/0 | 0/0 | 0/0 | 0/0 | 0/0 | 0/0 | 2/0 | 0/0 | 1/0 | 5/0 | 1/0 | 0/0 | 0/0 | 0/0 | 0/0 | 0/0 | 1/0 |
| XI | 0/0 | 0/0 | 0/0 | 0/0 | 0/0 | 1/0 | 0/0 | 1/0 | 2/0 | 0/0 | 1/0 | 0/0 | 0/0 | 0/0 | 0/0 | 0/0 | 0/0 | 0/0 |
| XII | 0/0 | 0/0 | 0/0 | 0/0 | 0/0 | 3/0 | 0/0 | 9/0 | 2/1 | 1/0 | 2/1 | 1/1 | 0/0 | 0/0 | 0/0 | 0/0 | 0/0 | 0/0 |
| XIII | 1/1 | 1/0 | 0/0 | 0/1 | 1/0 | 6/1 | 2/0 | 17/5 | 3/1 | 7/0 | 5/1 | 0/2 | 3/2 | 0/0 | 0/0 | 0/0 | 1/0 | 0/0 |
| XIV | 0/0 | 0/0 | 0/0 | 0/0 | 0/0 | 3/0 | 0/0 | 0/0 | 0/0 | 0/0 | 0/0 | 0/0 | 3/0 | 0/0 | 0/0 | 0/0 | 0/0 | 0/0 |
| XV | 0/0 | 0/0 | 0/0 | 0/0 | 0/0 | 6/0 | 0/0 | 4/0 | 0/1 | 1/0 | 0/0 | 4/0 | 1/0 | 0/0 | 0/0 | 0/0 | 0/0 | 0/0 |
| XVII | 0/0 | 0/0 | 0/0 | 0/0 | 0/0 | 0/0 | 0/0 | 1/1 | 0/0 | 1/0 | 0/0 | 0/1 | 6/1 | 1/0 | 0/0 | 0/0 | 0/0 | 0/0 |
| XVIII | 0/0 | 0/0 | 0/0 | 0/0 | 0/0 | 0/0 | 0/0 | 1/0 | 0/0 | 0/0 | 0/0 | 0/0 | 5/0 | 0/0 | 0/0 | 0/0 | 0/0 | 0/0 |
| XIX | 0/0 | 0/0 | 0/0 | 0/0 | 0/0 | 0/0 | 2/0 | 4/0 | 1/0 | 6/0 | 2/0 | 3/0 | 4/0 | 2/0 | 3/0 | 1/0 | 1/0 | 1/0 |
| Всего | 5/1 | 1/0 | 3/0 | 0/1 | 4/0 | 46/3 | 10/0 | 99/15 | 14/4 | 36/3 | 28/4 | 17/8 | 39/9 | 8/1 | 4/0 | 2/0 | 4/0 | 2/0 |
Таким образом, наблюдается мозаичность и неравномерность частот в распространении семейств вариантов блоков компонентов, контролируемых аллелями локусов Hrd A и Hrd B в исследованных образцах H. spontaneum из различных стран мира. Матрицу суммарных частот семейств одновременно гордеинов А и В в популяциях дикого ячменя из 16 стран, за исключением Армении и Египта (каждая из которых была представлена одним образцом), подвергли факторному анализу методом главных компонент. Первые две компоненты описывают 31.2% изменчивости частот семейств HRD A и HRD B (рис. 5). Как видно из рисунка, по первой главной компоненте популяции дикого ячменя разделились следующим образом: в отрицательную область значений вошли популяции из стран Азии восточнее Ирана, а также из Азербайджана и Кипра. В положительной области значений первой компоненты оказались популяции из Северной Африки (Ливия), Ближнего Востока и Ирана. Отметим, что аналогичное распределение популяций на основе суммарных частот семейств HRD A и HRD B было получено и при факторном анализе местных образцов культурного ячменя [1].

ОБСУЖДЕНИЕ
Анализ структуры полиморфизма гордеинов, контролируемых аллелями локусов Hrd A и Hrd B в образцах H. spontaneum и H. vulgare показал значительное сходство между ними. Среди 12 семейств блоков компонентов гордеина А в культурном ячмене и 16 семейств в диком ячмене общими оказались 13 семейств (с учетом семейства AXIII, в которое вошел вариант HRD A21, не отнесенный ни к одному из семейств в культурном ячмене). При этом в 11 семействах присутствовали варианты, обнаруженные как исключительно в H. spontaneum, так и идентичные ранее выявленным в H. vulgare. Аналогичное сходство между диким и культурным ячменем наблюдалось и в структуре полиморфизма вариантов блоков компонентов, контролируемых аллелями локуса Hrd B. Из 18 семейств гордеина B H. spontaneum в 12 семейств вошли варианты блоков компонентов, обнаруженные как в диком, так и в культурном ячмене. Разное число представителей семейств А и В гордеинов, выделенных в диком ячмене, были обнаружены в популяциях из всех стран от Ливии до Пакистана. При этом представители всех семейств HRD A и HRD B (за исключением BXIV) были найдены только в образцах из Израиля. Здесь же обнаружено наибольшее число “культурных” вариантов гордеинов А (18) и В (15), относящихся к шести и восьми семействам соответственно. Однако “культурные” варианты гордеина А, входящие в семейства AIII, AXI, AXII, AXIII и AXIV, ненайденные в израильских популяциях, присутствовали в H. spontaneum из Армении, Азербайджана, Турции, Палестины, Сирии, Иордании, Ирака, Ирана, Туркменистана, Узбекистана, Афганистана и Пакистана (табл. 4, приложение).
Подобным же образом, “культурные” варианты HRD B семейств BVIII, BIX, BXII и BXV, ненайденные в израильских популяциях H. spontaneum, обнаружены в образцах дикого ячменя из Турции, Палестины, Иордании, Ирака и Ирана (табл. 5, приложение). Такое “мозаичное” распространение как семейств гордеинов А и В, так и “культурных” вариантов, не исключает, что различные популяции H. spontaneum из стран от Ливии до Пакистана могли быть донорами тех или иных аллелей локусов Hrd A и Hrd B в результате спонтанной гибридизации с H. vulgare при его распространении. При этом, учитывая присутствие в популяциях дикого ячменя из Израиля представителей практически всех семейств HRD A и HRD B, а также самое большое число вариантов блоков компонентов, идентичных выявленным в культурном ячмене, можно предположить, что именно на территории Израиля был первоначально введен в культуру ячмень.
Наши данные согласуются с результатами геномного анализа с использованием 6152 SNP в 803 местных популяциях культурного ячменя из Европы, Северной Африки, Ближнего Востока и Азии, а также в 277 образцах H. spontaneum [12]. Эти авторы пришли к заключению, что генетический вклад отдельных диких популяций в культурный ячмень различается по всему геному. Отдельные геномы местных популяций H. vulgare из различных стран указывают на общую их родословную с географически близкими популяциями H. spontaneum. При этом самая большая доля вклада в родословную культурного ячменя (57%) происходит от популяции дикого ячменя из Южного Леванта (современный Израиль). В генофонд местных популяций культурного ячменя вклад вносят также популяции H. spontaneum из Северного Леванта (12%), Центральной Азии (11%), Северной Месопотамии (10%) и Сирийской пустыни (9%).
К аналогичным выводам привело и исследование экзома в 310 образцах дикого и культурного ячменя, которое позволило структурировать H. vulgare на шесть генетических групп [13]. Тем не менее, как показали авторы, все они произошли от единой популяции, которая появилась на западе Плодородного Полумесяца. Распространение культурного ячменя за пределы этого района сопровождалось потоком генов от местных популяций дикого ячменя и последующим отбором, что привело к появлению современной генетической характеристики H. vulgare, которая была интерпретирована как свидетельство множественного одомашнивания [13]. Этому выводу соответствуют и результаты кластерного анализа на основе суммарных частот семейств HRD A и HRD B в культурном и диком ячмене.
Важнейшим признаком, по которому отличаются культурный и дикий ячмень, является неломкость и ломкость колосового стержня. Вероятно, что первым шагом в одомашнивании дикого ячменя было выделение форм с прочным колосовым стержнем, не распадающимся на членики при созревании [14]. Ломкость колосового стержня обусловлена наличием двух комплементарных генов Btr1 и Btr2, локализованных в коротком плече хромосомы 3H ячменя. У культурных ячменей существуют три варианта генотипов. 1) Btr1Btr1btr2btr2 – восточный тип, обозначаемый R. Takahashi как “Е”; 2) btr1btr1Btr2Btr2 – западный тип “W” и 3) btr1btr1btr2btr2, или тип “we” [14]. Подробный анализ сортов и линий ячменя из разных районов мира показал, что большинство сортов (95–100%) из “восточных” районов – Китая, Южной Кореи и Южной Японии, состоят из сортов типа Е. В оставшейся части мира, которую R. Takahashi называет “западным” районом, включая Юго-Западную Азию, Европу, Россию, Манчжурию, Северную Корею, Северную Японию и большую часть Индии, 60–80% культурных ячменей имеют тип W, 20–40% – тип Е и очень редко – тип we, означая, таким образом, существование двух линий потомства у культурных ячменей, которые ведут свое происхождение из различных мест.
Современные исследования молекулярной структуры генов Btr1/btr1 и Btr2/btr2 с помощью секвенирования показало, что btr1, в сравнении с Btr1, несет делецию одной пары нуклеотидов в кодирующей последовательности, а btr2, в сравнении с Btr2, – делецию 11 пн. Анализ молекулярного датирования показал, что аллель btr1 возник 50 000 ± 10 000 лет назад, а аллель btr2 – 40 000 ± ± 10 000 лет назад. На основе анализа серии тестовых гибридов было установлено, что сорта типа btr1 широко распространены, но в основном встречаются у сортов ячменя Европы и Ближнего Востока. Сорта типа btr2 распространены в основном в Восточной Азии и в Северной Африке. Исследование разнообразия последовательностей в обоих локусах в выборке образцов культурного и дикого ячменя, показало, что предок культурных сортов типа btr2 произрастал в северной части Сирии и на юго-востоке Турции. H. spontaneum, наиболее близкий к btr1-типу, можно разделить на две группы, которые расположены в южной части Леванта, другая генетически однородная группа – в Центральной Азии. Наиболее генетически разнообразные сохранившиеся популяции дикого ячменя относятся к южной части Леванта [15]. Эти авторы считают вероятным, что азиатские образцы H. spontaneum могли происходить из материала, распространенного людьми при миграции из Леванта. Таким образом, прослеживая эволюционную историю аллельной изменчивости генов Btr1 и Btr2, можно заключить, что как в пространстве, так и во времени, независимые отборы генотипов с неломким колосовым стержнем были сделаны во время одомашнивания ячменя человеком в южных и северных регионах Леванта. Предполагается, что неравномерность распространения генотипов Btr1Btr1btr2btr2 и btr1btr1Btr2Btr2 на востоке и западе может быть связана, с одной стороны с лучшей адаптацией их к различным эколого-климатическим условиям, с другой стороны является прямым результатом миграции людей [15].
Еще одним аргументом, что Израиль может быть местом первоначальной доместикации ячменя, являются данные археологических исследований. На Ближнем Востоке самое раннее использование человеком зерна ячменя зафиксировано в Охало II (около Галилейского моря) и датируется XVII тысячелетием до н.э. [16]. Кроме этого, по данным археоботаников, климатические условия на территории современного Израиля и Палестины существенно не менялись за последние 10–12 тыс. лет [17]. Это позволяет надеяться на отсутствие существенного влияния внешних условий на генетическую структуру популяций как H. spontaneum, так и H. vulgare. Вместе с тем, при археологических раскопках в южном Египте около Асуана, в районе Каббания, были обнаружены зерна культурного ячменя, возраст которых оценивался в 18 000 лет [18]. Однако позднее эта датировка была уточнена – 4850 ± 200 лет до н.э. [19]. Таким образом, на сегодняшний день самые древние свидетельства использования ячменя человеком находятся в Охало II.
В заключении следует отметить мнение Н.И. Вавилова о центре происхождения ячменя. Так, по свидетельству Ф.Х. Бахтеева [20], в своей неопубликованной работе “Опыт эволюционной генетики культурных ячменей”, относящейся еще к 1939 г., Н.И. Вавилов “с предельной ясностью указывает, что происхождение культурных ячменей территориально связано с Передней Азией. Вследствие этого другие, прежде установленные им центры формообразования, он сам признает вторичными”. О значении Передней Азии как центра происхождения важнейших зерновых культур, Н.И. Вавилов пиcал в своем личном письме доктору Г. Филду (H. Field) 6 декабря 1938 г.: “Сейчас у меня нет сомнений в том, что первоначальной родиной пшеницы была Передняя Азия, включая Сирию, Палестину, Закавказье, Иран и Малую Азию. Здесь родина пшеницы, ячменя, ржи и некоторых зернобобовых” [21].
Настоящая статья не содержит каких-либо исследований с использованием в качестве объекта животных.
Настоящая статья не содержит каких-либо исследований с участием в качестве объекта людей.
Авторы заявляют, что у них нет конфликта интересов.
ПРИЛОЖЕНИЕ Частоты аллелей локусов Hrd A и Hrd B в исследованных образцах H. spontaneum из различных стран
| Alleles of locus Hrd A | Family | Libya | Egypt | Cyprus | Armenia | Azerbaijan | Turkey | Lebanon | Israel | Palestine | Syria | Jordan | Iraq | Iran | Turkmenistan | Uzbekistan | Tajikistan | Afghanistan | Pakistan |
| A1s | I | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0 | 0 | 0.0105 | 0 | 0.0233 | 0 | 0 | 0 | 0 | 0 |
| A2s | IV | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0077 | 0 | 0 | 0 | 0 | 0 |
| A3s | IV | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0116 | 0 | 0 | 0 | 0 | 0 |
| A4s | XV | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0116 | 0 | 0 | 0 | 0 | 0 |
| A5s | XIII | 0 | 0 | 0 | 0 | 0 | 0.0406 | 0 | 0 | 0.1176 | 0 | 0 | 0.01 | 0.0116 | 0 | 0 | 0 | 0 | 0 |
| A6s | XV | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0233 | 0 | 0 | 0 | 0 | 0 |
| A7s | XV | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0233 | 0 | 0 | 0 | 0 | 0 |
| A8s | XV | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0233 | 0 | 0 | 0 | 0 | 0 |
| A9s | XVII | 0 | 0 | 0 | 0 | 0 | 0.0135 | 0 | 0 | 0 | 0.0087 | 0 | 0.04 | 0.0233 | 0 | 0 | 0 | 0 | 0 |
| A10s | V | 0 | 0 | 0 | 0 | 0 | 0 | 0.1 | 0.008 | 0 | 0 | 0 | 0 | 0.0229 | 0 | 0 | 0 | 0 | 0 |
| A11s | V | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0 | 0 | 0 | 0 | 0.0233 | 0 | 0 | 0 | 0 | 0 |
| A12s | IX | 0 | 0 | 0 | 0 | 0 | 0.0135 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0465 | 0 | 0 | 0 | 0 | 0 |
| A13s | IX | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0233 | 0 | 0 | 0 | 0 | 0 |
| A14s | IX | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0233 | 0 | 0 | 0 | 0 | 0 |
| A15s | VIII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.006 | 0 | 0 | 0 | 0 | 0.0291 | 0 | 0 | 0 | 0 | 0 |
| A16s | VIII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0 | 0.0261 | 0 | 0 | 0.0291 | 0 | 0 | 0 | 0 | 0 |
| A17s | VIII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.21 | 0.0291 | 0 | 0 | 0 | 0 | 0 |
| A18s | VIII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0349 | 0 | 0 | 0 | 0 | 0 |
| A19s | XII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0824 | 0 | 0 | 0 | 0.0233 | 0 | 0 | 0 | 0 | 0 |
| A20s | XI | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0 | 0 | 0 | 0 | 0.0233 | 0 | 0 | 0 | 0 | 0 |
| A21s | I | 0 | 0 | 0 | 0 | 0 | 0.027 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A22s | I | 0 | 0 | 0 | 0 | 0 | 0.0135 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A23s | I | 0 | 0 | 0 | 0 | 0 | 0.0406 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A24s | I | 0 | 0 | 0 | 0 | 0 | 0.0135 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A25s | VIII | 0 | 0 | 0 | 0 | 0 | 0.0089 | 0.04 | 0.0159 | 0 | 0 | 0.0316 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A26s | VIII | 0 | 0 | 0 | 0 | 0 | 0.0135 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A27s | VIII | 0 | 0 | 0 | 0 | 0 | 0.0089 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0556 | 0 | 0 | 0 | 0 |
| A28s | VIII | 0 | 0 | 0 | 0 | 0 | 0.027 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A29s | IV | 0 | 0 | 0 | 0 | 0 | 0.0406 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A30s | VIII | 0 | 0 | 0 | 0.2 | 0 | 0.0541 | 0.04 | 0.0577 | 0.0588 | 0 | 0.0211 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A31s | VIII | 0 | 0 | 0 | 0 | 0 | 0.027 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A32s | V | 0 | 0 | 0 | 0 | 0 | 0.063 | 0 | 0 | 0 | 0 | 0.0632 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A33s | V | 0 | 0 | 0 | 0 | 0 | 0.0135 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A34s | XIII | 0 | 0 | 0 | 0 | 0 | 0.0135 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A35s | XV | 0 | 0 | 0 | 0 | 0 | 0.027 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A36s | XVII | 0 | 0 | 0 | 0 | 0 | 0.0406 | 0 | 0 | 0 | 0 | 0 | 0.01 | 0 | 0 | 0 | 0 | 0 | 0 |
| A37s | XIII | 0 | 0 | 0 | 0 | 0 | 0.0135 | 0.1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A38s | XIII | 0 | 0 | 0 | 0 | 0 | 0.0089 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A39s | X | 0 | 0 | 0 | 0 | 0 | 0.0135 | 0 | 0.002 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A40s | I | 0 | 0 | 0 | 0 | 0 | 0.0135 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A41s | IX | 0 | 0 | 0 | 0 | 0 | 0.0135 | 0 | 0.002 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A42s | I | 0 | 0 | 0 | 0 | 0 | 0.0135 | 0.06 | 0 | 0 | 0.0435 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A43s | I | 0 | 0 | 0 | 0 | 0 | 0 | 0.18 | 0 | 0 | 0.0435 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A44s | I | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0783 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A45s | I | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0087 | 0.0105 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A46s | I | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0384 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A47s | V | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0435 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A48s | IV | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.6 |
| A49s | I | 0 | 0.4 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A50s | I | 0 | 0.6 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A51s | XVII | 0 | 0 | 0.4 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0174 | 0 | 0 | 0 | 0.1 | 0.55 | 0.7 | 0 | 0 |
| A52s | VII | 0.086 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0 | 0.0174 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A53s | VII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.006 | 0 | 0.1304 | 0.0526 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A54s | XVI | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0261 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A55s | XVI | 0 | 0 | 0 | 0 | 0 | 0 | 0.02 | 0.0298 | 0 | 0.0087 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A56s | XVI | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0087 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A57s | IX | 0.114 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A58s | VI | 0 | 0 | 0 | 0 | 0 | 0 | 0.04 | 0 | 0 | 0.0261 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A59s | XII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0087 | 0.0421 | 0.05 | 0 | 0 | 0 | 0 | 0 | 0 |
| A60s | X | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0421 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A61s | X | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0087 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A62s | XIII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0087 | 0.0105 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A63s | IV | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0435 | 0.0316 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A64s | V | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0358 | 0 | 0 | 0 | 0.02 | 0 | 0 | 0 | 0 | 0 | 0 |
| A65s | V | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.01 | 0 | 0 | 0 | 0 | 0 | 0 |
| A66s | XIII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.01 | 0 | 0 | 0 | 0 | 0 | 0 |
| A67s | I | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A68s | XIII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.01 | 0 | 0 | 0 | 0 | 0 | 0 |
| A69s | XVI | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.02 | 0 | 0 | 0 | 0 | 0 | 0 |
| A70s | XVI | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.01 | 0 | 0 | 0 | 0 | 0 | 0 |
| A71s | VIII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.03 | 0 | 0 | 0 | 0 | 0 | 0 |
| A72s | XVII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.14 | 0 | 0 | 0 | 0 | 0.333 | 0 |
| A73s | X | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.05 | 0 | 0 | 0 | 0 | 0 | 0 |
| A74s | I | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0105 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A75s | V | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0105 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A76s | XIII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.01 | 0 | 0 | 0 | 0 | 0 | 0 |
| A77s | I | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0526 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A78s | I | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0421 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A79s | IX | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.133 | 0 |
| A80s | IV | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0105 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A81s | XVII | 0 | 0 | 0.267 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A82s | VII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0435 | 0.1263 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A83s | XIII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0435 | 0.0421 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A84s | XIII | 0 | 0 | 0 | 0.2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A85s | IX | 0.143 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A86s | V | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.1352 | 0.0421 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A87s | XII | 0 | 0 | 0 | 0 | 0.8 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.2 |
| A88s | XVI | 0 | 0 | 0 | 0 | 0 | 0 | 0.1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A89s | XVI | 0 | 0 | 0 | 0 | 0 | 0 | 0.1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A90s | XVI | 0 | 0 | 0 | 0 | 0 | 0 | 0.1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A91s | XIV | 0 | 0 | 0 | 0 | 0 | 0 | 0.04 | 0.002 | 0.0588 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A92s | VI | 0 | 0 | 0 | 0 | 0 | 0 | 0.02 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A93s | VI | 0 | 0 | 0 | 0 | 0 | 0 | 0.06 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A94s | IX | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0 | 0 | 0 | 0 | 0 | 0.1889 | 0.4 | 0 | 0 | 0.1 |
| A95s | V | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0222 | 0 | 0.3 | 0 | 0 |
| A96s | V | 0.2 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0 | 0 | 0 | 0 | 0 | 0.0111 | 0 | 0 | 0 | 0 |
| A97s | I | 0.029 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A98s | I | 0.029 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A99s | I | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.1889 | 0 | 0 | 0 | 0 |
| A100s | X | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.1333 | 0 | 0 | 0 | 0 |
| A101s | I | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0118 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A102s | VIII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0118 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A103s | XIV | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.1059 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A104s | I | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0706 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A105s | I | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0.0118 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A106s | I | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0.0353 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A107s | I | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0.0588 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A108s | I | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0353 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A109s | XII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.006 | 0.0471 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A110s | XVI | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0353 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A111s | XIII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0235 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A112s | IX | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0457 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A113s | XIII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.006 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A114s | IV | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.008 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A115s | IV | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.004 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A116s | XII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0338 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A117s | XVII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0159 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A118s | VIII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.006 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A119s | XIII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.004 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A120s | XIII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A121s | XIII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A122s | XV | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A123s | V | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A124s | I | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A125s | I | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A126s | I | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.004 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A127s | V | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A128s | I | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A129s | IV | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.006 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A130s | I | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A131s | I | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.004 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A132s | IV | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.004 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A133s | V | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0179 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A134s | I | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0098 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A135s | V | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A136s | XIV | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A137s | I | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A138s | V | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.006 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A139s | V | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A140s | V | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A141s | XVI | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A142s | V | 0 | 0 | 0.333 | 0 | 0 | 0 | 0 | 0.002 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A143s | IV | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A144s | X | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A145s | V | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A146s | VI | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.004 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A147s | XI | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A148s | XII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0118 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A149s | V | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0098 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A150s | XIV | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0298 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A151s | X | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.004 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A152s | XII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0098 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A153s | V | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0118 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A154s | XVI | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.006 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A155s | IV | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.004 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A156s | IV | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A157s | XI | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A158s | I | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A159s | V | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0219 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A160s | VIII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A161s | I | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0138 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A162s | III | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0138 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A163s | XII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.004 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A164s | IX | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.004 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A165s | IX | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.004 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A166s | XVII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A167s | X | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A168s | V | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A169s | XVII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0138 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A170s | XII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0098 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A171s | XIV | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.004 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A172s | XII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A173s | IX | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A174s | XIII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0199 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A175s | XVII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0098 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A176s | XV | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.004 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A177s | XV | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.006 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A178s | IV | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A179s | IV | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A180s | XIV | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.006 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A181s | XIV | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0358 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A182s | XIV | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.004 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A183s | XVI | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.008 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A184s | XIV | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A185s | XIV | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A186s | VIII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0556 | 0 | 0 | 0 | 0 |
| A187s | IX | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0526 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A188s | I | 0.086 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A189s | I | 0.143 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A190s | I | 0.057 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A191s | V | 0.029 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A192s | XVI | 0.057 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| А1 | V | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0933 | 0 | 0 | 0.0421 | 0.03 | 0.0349 | 0 | 0 | 0 | 0 | 0 |
| А2 | I | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0 | 0 | 0 | 0 | 0.0174 | 0 | 0 | 0 | 0 | 0 |
| А3 | VII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.008 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| А12 | V | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0199 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| А13 | IX | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A14 | III | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.1 |
| А16 | IV | 0 | 0 | 0 | 0 | 0 | 0.0089 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0309 | 0 | 0 | 0 | 0 | 0 |
| А18 | V | 0 | 0 | 0 | 0 | 0 | 0.0678 | 0 | 0.0278 | 0 | 0 | 0 | 0.1 | 0.217 | 0.0444 | 0 | 0 | 0 | 0 |
| А21 | XIII | 0 | 0 | 0 | 0 | 0 | 0.0135 | 0 | 0 | 0 | 0 | 0 | 0.04 | 0.0116 | 0 | 0 | 0 | 0 | 0 |
| A22 | XII | 0 | 0 | 0 | 0.6 | 0.2 | 0.063 | 0 | 0 | 0 | 0.0261 | 0.1053 | 0.02 | 0.0465 | 0 | 0 | 0 | 0 | 0 |
| А23 | IV | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.004 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| А24 | I | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.004 | 0 | 0 | 0.0105 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| А28 | IV | 0.029 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0 | 0 | 0 | 0 | 0.0233 | 0 | 0 | 0 | 0 | 0 |
| А33 | IX | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0557 | 0 | 0 | 0.0105 | 0.05 | 0 | 0 | 0 | 0 | 0 | 0 |
| А35 | VI | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.004 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| А40 | V | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0138 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| А61 | IX | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0 | 0.0174 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| A62 | XIV | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0949 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| А63 | I | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.006 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| А67 | I | 0 | 0 | 0 | 0 | 0 | 0.0135 | 0 | 0.002 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| А68 | XI | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0698 | 0.0556 | 0 | 0 | 0.333 | 0 |
| А78 | XI | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.1176 | 0 | 0 | 0 | 0.0233 | 0 | 0 | 0 | 0 | 0 |
| A93 | V | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0588 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| А97 | XII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0588 | 0.0609 | 0.0211 | 0.13 | 0 | 0 | 0.05 | 0 | 0 | 0 |
| А98 | XII | 0 | 0 | 0 | 0 | 0 | 0.2436 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0233 | 0.1444 | 0 | 0 | 0.2 | 0 |
| А103 | III | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0233 | 0 | 0 | 0 | 0 | 0 |
| А106 | IV | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0105 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| А107 | I | 0 | 0 | 0 | 0 | 0 | 0.0135 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| А110 | I | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| А123 | I | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0098 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| А139 | IV | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0261 | 0 | 0 | 0.0116 | 0 | 0 | 0 | 0 | 0 |
| А144 | I | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0261 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| А152 | I | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0338 | 0 | 0.0261 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Alleles of locus Hrd B | Family | Libya | Egypt | Cyprus | Armenia | Azerbaijan | Turkey | Lebanon | Israel | Palestine | Syria | Jordan | Iraq | Iran | Turkmenistan | Uzbekistan | Tajikistan | Afghanistan | Pakistan |
| B1s | I | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0233 | 0 | 0 | 0 | 0 | 0 |
| B2s | IV | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0233 | 0 | 0 | 0 | 0 | 0 |
| B3s | XIX | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0815 | 0.0235 | 0 | 0 | 0 | 0 |
| B4s | XIX | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.05 | 0.0233 | 0 | 0 | 0 | 0 | 0 |
| B5s | XIX | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0466 | 0 | 0 | 0 | 0 | 0 |
| B6s | XIX | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0233 | 0 | 0 | 0 | 0 | 0 |
| B7s | II | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0077 | 0 | 0 | 0 | 0 | 0 |
| B8s | XV | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0233 | 0 | 0 | 0 | 0 | 0 |
| B9s | IX | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0526 | 0 | 0.0233 | 0 | 0 | 0 | 0 | 0 |
| B10s | IV | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0116 | 0 | 0 | 0 | 0 | 0 |
| B11s | VII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0233 | 0 | 0 | 0 | 0 | 0 |
| B12s | V | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0139 | 0.0353 | 0 | 0 | 0 | 0.0543 | 0 | 0 | 0 | 0 | 0 |
| B13s | V | 0 | 0 | 0 | 0 | 0 | 0.0135 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0466 | 0 | 0 | 0 | 0 | 0 |
| B14s | VII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0116 | 0 | 0 | 0 | 0 | 0 |
| B15s | IX | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.004 | 0 | 0 | 0 | 0.05 | 0.0116 | 0 | 0 | 0 | 0 | 0 |
| B16s | IX | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0116 | 0 | 0 | 0 | 0 | 0 |
| B17s | IX | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0077 | 0 | 0 | 0 | 0 | 0 | |
| B18s | VII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0233 | 0 | 0 | 0 | 0 | 0 |
| B19s | VIII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0 | 0 | 0 | 0 | 0.0233 | 0 | 0 | 0 | 0 | 0 |
| B20s | XVIII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0466 | 0 | 0 | 0 | 0 | 0 |
| B21s | XVIII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0233 | 0 | 0 | 0 | 0 | 0 |
| B22s | XVIII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0233 | 0 | 0 | 0 | 0 | 0 |
| B23s | XVIII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0116 | 0 | 0 | 0 | 0 | 0 |
| B24s | XVIII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0233 | 0 | 0 | 0 | 0 | 0 |
| B25s | IX | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0233 | 0 | 0 | 0 | 0 | 0 |
| B26s | XIV | 0 | 0 | 0 | 0 | 0 | 0.0089 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0233 | 0 | 0 | 0 | 0 | 0 |
| B27s | XVII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0077 | 0 | 0 | 0 | 0 | 0 |
| B28s | XVII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0077 | 0 | 0 | 0 | 0 | 0 |
| B29s | XVII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0087 | 0 | 0 | 0.0077 | 0 | 0 | 0 | 0 | 0 |
| B30s | XVII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0077 | 0 | 0 | 0 | 0 | 0 |
| B31s | XVII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0154 | 0 | 0 | 0 | 0 | 0 |
| B32s | XVII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0233 | 0 | 0 | 0 | 0 | 0 |
| B33s | XIII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0 | 0 | 0 | 0 | 0.0114 | 0 | 0 | 0 | 0 | 0 |
| B34s | XIII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0115 | 0 | 0 | 0 | 0 | 0 |
| B35s | XIII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0233 | 0 | 0 | 0 | 0 | 0 |
| B36s | IV | 0 | 0 | 0 | 0 | 0 | 0 | 0.1 | 0 | 0 | 0 | 0 | 0 | 0.0116 | 0 | 0 | 0 | 0 | 0 |
| B37s | XIV | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0466 | 0 | 0 | 0 | 0 | 0 |
| B38s | XIV | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0233 | 0 | 0 | 0 | 0 | 0 |
| B39s | IX | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0077 | 0 | 0 | 0 | 0 | 0 |
| B40s | II | 0 | 0 | 0 | 0 | 0 | 0.0225 | 0 | 0 | 0 | 0 | 0 | 0.01 | 0 | 0 | 0 | 0 | 0 | 0 |
| B41s | II | 0 | 0 | 0 | 0 | 0 | 0.0135 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B42s | IV | 0 | 0 | 0 | 0 | 0 | 0.0157 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B43s | IV | 0 | 0 | 0 | 0 | 0 | 0.0271 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B44s | IV | 0 | 0 | 0 | 0 | 0 | 0.0135 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B45s | IV | 0 | 0 | 0 | 0 | 0 | 0.0542 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B46s | IV | 0 | 0 | 0 | 0 | 0 | 0.0271 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B47s | V | 0 | 0 | 0 | 0 | 0 | 0.0135 | 0 | 0.004 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B48s | VI | 0 | 0 | 0 | 0 | 0 | 0.0089 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B49s | XIII | 0 | 0 | 0 | 0 | 0 | 0.0135 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B50s | V | 0 | 0 | 0 | 0 | 0 | 0.0271 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B51s | XIII | 0 | 0 | 0 | 0 | 0 | 0.0721 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B52s | XIII | 0 | 0 | 0 | 0 | 0 | 0.0766 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B53s | XIII | 0 | 0 | 0 | 0 | 0 | 0.0271 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B54s | IX | 0 | 0 | 0 | 0 | 0 | 0.0271 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B55s | VIII | 0 | 0 | 0 | 0 | 0 | 0.0271 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B56s | IX | 0 | 0 | 0 | 0 | 0 | 0.0407 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B57s | V | 0 | 0 | 0 | 0 | 0 | 0.0135 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B58s | VIII | 0 | 0 | 0 | 0 | 0 | 0.0135 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B59s | VII | 0 | 0 | 0 | 0 | 0 | 0.0135 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B60s | VII | 0 | 0 | 0 | 0 | 0 | 0.0135 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B61s | XII | 0 | 0 | 0 | 0 | 0 | 0.0089 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B62s | XIV | 0 | 0 | 0 | 0 | 0 | 0.0135 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B63s | XII | 0 | 0 | 0 | 0 | 0 | 0.0135 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B64s | VIII | 0 | 0 | 0 | 0 | 0 | 0.0271 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B65s | XIV | 0 | 0 | 0 | 0 | 0 | 0.0135 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B66s | XIII | 0 | 0 | 0 | 0 | 0 | 0.0089 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B67s | I | 0 | 0 | 0 | 0 | 0 | 0.0089 | 0 | 0 | 0.0353 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B68s | I | 0 | 0 | 0 | 0 | 0 | 0.0271 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B69s | III | 0 | 0 | 0 | 0 | 0 | 0.0089 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B70s | III | 0 | 0 | 0 | 0 | 0 | 0.0089 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B71s | III | 0 | 0 | 0 | 0 | 0 | 0.0135 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B72s | XI | 0 | 0 | 0 | 0 | 0 | 0.0135 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B73s | XV | 0 | 0 | 0 | 0 | 0 | 0.0135 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B74s | XV | 0 | 0 | 0 | 0 | 0 | 0.0271 | 0 | 0 | 0 | 0 | 0 | 0.07 | 0 | 0 | 0 | 0 | 0 | 0 |
| B75s | XV | 0 | 0 | 0 | 0 | 0 | 0.0407 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B76s | XV | 0 | 0 | 0 | 0 | 0 | 0.0271 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B77s | XV | 0 | 0 | 0 | 0 | 0 | 0.0135 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B78s | XV | 0 | 0 | 0 | 0 | 0 | 0.0271 | 0 | 0 | 0 | 0 | 0 | 0.01 | 0 | 0 | 0 | 0 | 0 | 0 |
| B79s | XIII | 0 | 0 | 0 | 0 | 0 | 0.0135 | 0 | 0 | 0 | 0 | 0.0105 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B80s | XII | 0 | 0 | 0 | 0 | 0 | 0.0135 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B81s | VIII | 0 | 0 | 0 | 0 | 0 | 0.0089 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B82s | VII | 0 | 0 | 0 | 0 | 0 | 0.0068 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B83s | I | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0261 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B84s | I | 0 | 0 | 0 | 0 | 0 | 0 | 0.02 | 0 | 0 | 0.0261 | 0 | 0 | 0 | 0.0235 | 0 | 0 | 0 | 0 |
| B85s | I | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0 | 0.0087 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B86s | II | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0174 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B87s | XIII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0 | 0.0347 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B88s | I | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0087 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B89s | I | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0857 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B90s | I | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0696 | 0.0105 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B91s | IV | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0174 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B92s | XIII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0087 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B93s | XIII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0174 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B94s | XIII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0087 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B95s | V | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0174 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B96s | XV | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0174 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B97s | V | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0 | 0.0087 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B98s | V | 0 | 0 | 0 | 0 | 0 | 0 | 0.08 | 0.002 | 0 | 0.0261 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B99s | XIX | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0087 | 0 | 0.05 | 0 | 0 | 0 | 0 | 0 | 0 |
| B100s | XIX | 0 | 0 | 0 | 0 | 0 | 0 | 0.28 | 0 | 0 | 0.0435 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B101s | XIX | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0347 | 0.0948 | 0.13 | 0 | 0 | 0.3 | 0 | 0 | 0.7 |
| B102s | XIX | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0261 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B103s | XIX | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0087 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B104s | XIX | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0261 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B105s | VII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0435 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B106s | VIII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0087 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B107s | V | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0261 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B108s | V | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0087 | 0 | 0.04 | 0 | 0 | 0 | 0 | 0 | 0 |
| B109s | VI | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0087 | 0 | 0.02 | 0 | 0 | 0 | 0 | 0 | 0 |
| B110s | IX | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0521 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B111s | I | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0435 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B112s | X | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0087 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B113s | XII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0261 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B114s | XIII | 0 | 0 | 0 | 0 | 0 | 0 | 0.2 | 0 | 0 | 0.0087 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B115s | VIII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.01 | 0 | 0.01 | 0 | 0 | 0 | 0 | 0 | 0 |
| B116s | XIX | 0 | 0 | 0 | 0 | 0 | 0 | 0.1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B117s | I | 0 | 0 | 0 | 0 | 0 | 0 | 0.02 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B118s | I | 0 | 0 | 0 | 0 | 0 | 0 | 0.02 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B119s | XIII | 0 | 0 | 0 | 0 | 0 | 0 | 0.08 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B120s | II | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.25 | 0 | 0 | 0 |
| B121s | XIX | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.05 | 0 | 0 | 0 |
| B122s | XIX | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.1765 | 0.4 | 0.8 | 0 | 0 |
| B123s | V | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0 | 0 | 0 | 0 | 0 | 0.1529 | 0 | 0 | 0 | 0 |
| B124s | III | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.1059 | 0 | 0 | 0 | 0 |
| B125s | IX | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0706 | 0 | 0 | 0 | 0 |
| B126s | IX | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0706 | 0 | 0 | 0 | 0 |
| B127s | XVII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0118 | 0 | 0 | 0 | 0 |
| B128s | XII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.153 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B129s | XII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0588 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B130s | XI | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0118 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B131s | XI | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.1059 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B132s | VI | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0706 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B126s | IX | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0706 | 0 | 0 | 0 | 0 |
| B127s | XVII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0118 | 0 | 0 | 0 | 0 |
| B128s | XII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.153 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B129s | XII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0588 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B130s | XI | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0118 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B131s | XI | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.1059 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B132s | VI | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0706 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B133s | XIII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0824 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B134s | XIII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.004 | 0.0588 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B135s | XIX | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0588 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B136s | II | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0588 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B137s | I | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0235 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B138s | IX | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0353 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B139s | XIII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0588 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B140s | II | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.004 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B141s | II | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.006 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B142s | II | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B143s | IV | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B144s | IX | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B145s | XV | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B146s | XIII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.006 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B147s | VII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.004 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B148s | XV | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B149s | I | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.008 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B150s | XIII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.004 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B151s | VII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B152s | II | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0119 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B153s | XII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B154s | VIII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B155s | IV | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.004 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B156s | IX | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0099 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B157s | XIII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B158s | XIII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.006 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B159s | IX | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B160s | XVII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0298 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B161s | XIII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0596 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B162s | VI | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.008 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B163s | XIII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0159 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B164s | XIX | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.004 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B165s | VIII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0658 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B166s | IX | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.006 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B167s | XIII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B168s | V | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B169s | IX | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B170s | I | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.004 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B171s | V | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0119 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B172s | IV | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.006 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B173s | XII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B174s | V | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0099 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B175s | XIII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B176s | XII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B177s | X | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B178s | XI | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B179s | III | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B180s | IV | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B181s | XII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B182s | XII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0119 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B183s | VIII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B184s | IX | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0099 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B185s | IV | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0716 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B186s | IV | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.08 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B187s | XII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B188s | III | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0398 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B189s | XIX | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0099 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B190s | V | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B191s | V | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0298 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B192s | II | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0099 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B193s | I | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.004 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B194s | V | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0119 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B195s | III | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.006 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B196s | VII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.006 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B197s | IV | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0159 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B198s | III | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.004 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B199s | VII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B200s | IV | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0159 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B201s | I | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0119 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B202s | VI | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.004 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B203s | XIII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.006 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B204s | XVIII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B205s | XIII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0119 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B206s | V | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0159 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B207s | I | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.004 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B208s | XV | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.008 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B209s | XV | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B210s | VII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.004 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B211s | II | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.006 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B212s | XIII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B213s | XIII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0298 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B214s | XII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B215s | I | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B216s | IV | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.05 | 0 | 0 | 0 | 0 | 0 | 0 |
| B217s | VII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0159 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B218s | XII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B219s | VII | 0 | 0 | 0 | 0 | 0 | 0 | 0.1 | 0 | 0 | 0 | 0 | 0.06 | 0 | 0.2471 | 0 | 0 | 0 | 0 |
| B220s | V | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.006 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B221s | XIX | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0199 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B222s | XIX | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.008 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B223s | X | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.01 | 0 | 0 | 0 | 0 | 0 | 0 |
| B224s | VIII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0378 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B225s | XIX | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.2 | 0 |
| B226s | VIII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.02 | 0 | 0 | 0 | 0 | 0 | 0 |
| B227s | VIII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.133 | 0 |
| B228s | XIII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.333 | 0 |
| B229s | XV | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.04 | 0 | 0 | 0 | 0 | 0 | 0 |
| B230s | XV | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.05 | 0 | 0 | 0 | 0 | 0 | 0 |
| B231s | II | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.333 | 0 |
| B232s | XII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.05 | 0 | 0 | 0 | 0 | 0 | 0 |
| B233s | XIII | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B234s | I | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0526 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B235s | I | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0737 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B236s | XIX | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0526 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B237s | II | 0 | 0 | 0.267 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B238s | I | 0 | 0 | 0.333 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0421 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B239s | X | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0526 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B240s | VII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0105 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B241s | XIII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0211 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B242s | IV | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0105 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B243s | X | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0211 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B244s | IV | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0105 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B245s | X | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0 | 0 | 0.0421 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B246s | I | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0316 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B247s | XII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.004 | 0 | 0 | 0.0211 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B248s | XIII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.006 | 0 | 0 | 0.0105 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B249s | XIII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0421 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B250s | X | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0421 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B251s | XII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0105 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B252s | III | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.006 | 0 | 0 | 0.0316 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B253s | XII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0421 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B254s | I | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0 | 0 | 0.0211 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B255s | III | 0 | 0 | 0.4 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B256s | XI | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0421 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B257s | I | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0316 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B258s | X | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0211 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B259s | I | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0211 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B260s | XIII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0099 | 0 | 0.0521 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B261s | XIII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0435 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B262s | IV | 0.257 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B263s | VIII | 0.029 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B264s | XIII | 0.114 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B265s | IX | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.2 | 0 | 0 |
| B266s | X | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.3 |
| B267s | I | 0.286 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B268s | VIII | 0.029 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B269s | XIII | 0 | 0 | 0 | 0 | 0.3 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B270s | IX | 0 | 0 | 0 | 0 | 0.45 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B271s | VII | 0 | 0 | 0 | 0 | 0.1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B272s | VII | 0 | 0 | 0 | 0 | 0.15 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B273s | VII | 0 | 0 | 0 | 0 | 0 | 0.0338 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B1 | I | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.004 | 0 | 0 | 0 | 0 | 0.0077 | 0 | 0 | 0 | 0 | 0 |
| B8 | XII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0235 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B12 | XIII | 0 | 0 | 0 | 0 | 0 | 0.0089 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B16 | IV | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| В15 | IV | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B17 | XIII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0077 | 0 | 0 | 0 | 0 | 0 |
| B19 | IX | 0 | 0 | 0 | 0 | 0 | 0.0089 | 0 | 0 | 0 | 0 | 0.0105 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B21 | XVII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.02 | 0.0077 | 0 | 0 | 0 | 0 | 0 |
| B25 | III | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0349 | 0.1176 | 0 | 0 | 0 | 0 |
| В29 | XIII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.004 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B35 | IX | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0077 | 0 | 0 | 0 | 0 | 0 |
| В41 | II | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0 | 0 | 0 | 0.01 | 0 | 0 | 0 | 0 | 0 | 0 |
| B61 | VI | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0087 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B65 | IV | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.004 | 0 | 0 | 0.0105 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B82 | XIII | 0.286 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B86 | VIII | 0 | 0 | 0 | 0 | 0 | 0.0068 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B88 | VIII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0706 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B118 | IV | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0435 | 0 | 0 | 0.0116 | 0 | 0 | 0 | 0 | 0 |
| B141 | XIII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0 | 0 | 0.0421 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| В142 | XIII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.008 | 0 | 0 | 0 | 0.01 | 0 | 0 | 0 | 0 | 0 | 0 |
| B145 | XIII | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B149 | II | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.02 | 0 | 0 | 0 | 0 | 0 | 0 |
| В150 | III | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| В162 | XIII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0239 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B163 | XIII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0.0353 | 0 | 0 | 0.03 | 0 | 0 | 0 | 0 | 0 | 0 |
| B166 | IV | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.06 | 0 | 0 | 0 | 0 | 0 | 0 |
| B168 | XII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.02 | 0 | 0 | 0 | 0 | 0 | 0 |
| В169 | VI | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.004 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B182 | XIII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0233 | 0 | 0 | 0 | 0 | 0 |
| В189 | VI | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0 | 0 | 0 | 0.11 | 0 | 0 | 0 | 0 | 0 | 0 |
| B213 | XV | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0235 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B217 | XVII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.004 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| B228 | V | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.002 | 0 | 0.0521 | 0 | 0 | 0.0116 | 0 | 0 | 0 | 0 | 0 |
| B251 | II | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0077 | 0 | 0 | 0 | 0 | 0 |
| B265 | XII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0105 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
Список литературы
Поморцев А.А., Рубанович А.В., Лялина Е.В. Структура и пути формирования полиморфизма гордеинов, контролируемых аллелями гордеин-кодирующих локусов в культурном ячмене (Hordeum vulgare L.) // Генетика. 2021. Т. 57. № 5. С. 544–556. https://doi.org/10.31857/S0016675821050088
Поморцев А.А., Нецветаев В.П., Созинов А.А. Полиморфизм культурного ячменя (Hordeum vulgare L.) по гордеинам // Генетика. 1985. Т. 21. № 4. С. 629–639.
Shewry P.R., Bunce N.A.C., Kreis M. et al. Polymorphism at the Hor 1 locus of barley (Hordeum vulgare L.) // Biochemical Genetics. 1985. V. 23. № 5/6. P. 391–403.
Bunce N.A.C., Forde B.G., Kreis M. et al. DNA restriction fragment length polymorphism of hordein loci: Application to identifying and fingerprinting barley cultivars // Seed Sci. Techn. 1986. V. 14. P. 419–429.
Саянова О.В., Мехедов С.Л., Буркитбаев Е.М. и др. Структурная организация генов С-гордеинов ячменя // Генетика. 1994. Т. 30. № 6. С. 749–755.
Sayanova O., Pomortsev A., Ladogina M. et al. Analysis of C-hordein deficient mutant of barley // Barley Genet. Newslet. 1992. V. 22. P. 53–56.
Kanazin V., Ananiev E., Blake T. Variability among members of Hor-2 multigene family // Genome. 1993. V. 26. P. 397–403.
von Bothmer R., Jacobsen N., Baden C. et al. An Ecogegraphical Study of the Genus Hordeum. Rome: International Plant Genetic Resources Institute, 1991. P. 25.
Поморцев А.А., Лялина Е.В. Аллельное разнообразие гордеин-кодирующих локусов Hrd A и Hrd B у культурного (Hordeum vulgare L.) и дикого (Hordeum spontaneum C. Koch) ячменя в Иране (как части Дуги Плодородия) // Генетика. 2016. Т. 52. № 10. С. 1146–1158. https://doi.org/10.7868/S001667581610009X
Созинов А.А., Попереля Ф.А. Методика вертикального дискового электрофореза в крахмальном геле и генетический принцип классификации глиадинов. Одесса, 1978. 16 с.
Поморцев А.А., Лялина Е.В. Оценка сортовой принадлежности и сортовой чистоты семян ячменя методом электрофоретического анализа запасных белков зерна // Методич. пособие к практикуму “Белковые маркеры для генетической паспортизации и улучшения геномов растений хозяйственно ценных видов”. М.: Цифровичок, 2011. 86 с.
Poets A.M., Fang Z. Clegg M.T. et al. Barley landraces are characterized by geographically heterogeneous genomic origin // Genome Biology. 2015. V. 16. № 173. https://doi.org/10.1186/s13059-015-0712-3
Civáň P., Drosou K., Armisen-Gimenez D. Episodes of gene flow and selection during the evolutionary history of domesticated barley // BMC Genomic. 2021. V. 22. № 227. https://doi.org/0.1186/s12864-021-07511-7
Takahashi R. The origin and evolution of cultivated barley // Advances in Genetics. N.Y.: Academic Press Inc. Publishers, 1955. V. II. P. 227–266.
Pourkheirandish M., Hensel G., Kilian B. et al. Evolution of the grain dispersal system in barley // Cell. 2015. V. 162. P. 527–539. https://doi.org/10.1016/j.cell.2015.07.002
Willcox G. Archeobotanical evidence for the beginnings of agriculture in Southwest Asia // Proc. of Harlan Symp. The Origins of Agriculture and Crop Domestication. Aleppo, 10–14 May, 1997. P. 25–38.
Лисицына Г.Н., Прищепенко Л.В. Палеоботанические находки Кавказа и Ближнего Востока. М.: Наука, 1977. 115 с.
Wendorf F., Schild R., El Hadidi N. et al. Use of barley in the Egyptian late paleolithic // Science. 1979. V. 205. P. 1341–1347. https://doi.org/10.1126/science.205.4413.1341
Wendorf F., Endore F., Schild R. et al. New radiocarbon dates on the cereals from Wadi Kubbaniya // Science. 1984. V. 225. № 4662. P. 645–646.
Бахтеев Ф.Х. Эколого-географические основы филогении и селекции ячменей Hordeum sativum Jess. М.-Л.: Изд-во академии наук СССР, 1948. 202 с.
Вавилов Н.И. Научное наследие в письмах. Международная переписка. М.: Наука, 2003. Т. VI (1938–1940). С. 101.
Дополнительные материалы отсутствуют.