

Геохимия, 2020, T. 65, № 10, стр. 998-1008
Биогеохимия импактных регионов: роль эдафических и фитоценотических факторов среды
В. С. Безель a, *, Т. В. Жуйкова a, b, **, В. А. Гордеева b, Е. В. Голоушкина b
a Институт экологии растений и животных УрО РАН
620144 Екатеринбург, ул. 8-е Марта, 202, Россия
b Нижнетагильский государственный социально-педагогический институт (филиал)
ФГАОУ ВО “Российский государственный профессионально-педагогический университет”
622031 Нижний Тагил, ул. Красногвардейская, 57, Россия
* E-mail: bezel@ipae.uran.ru
** E-mail: hbfnt@rambler.ru
Поступила в редакцию 10.03.2020
После доработки 04.04.2020
Принята к публикации 10.04.2020
Аннотация
Исследован вынос химических элементов надземной и подземной фитомассами травяных фитоценозов и последующий их возврат в процессе разложения растительных остатков. Результаты работы позволили оценить биогеохимические циклы ряда эссенциальных (Zn, Cu) и токсичных (Pb, Cd) элементов в природных биогеоценозах Среднего Урала. Показано, что интенсивность подобного обмена на территориях, подверженных различному уровню антропогенной трансформации, определяется не только прямым влиянием подвижных форм химических элементов, которые содержатся в почвах и выступают в роли загрязнителей природной среды, но и совокупностью эдафических (физико-химические параметры почв), ценотических (обилие и соотношение в фитоценозе агроботанических групп) и микробиологических (уровень развития почвенного микробоценоза) условий.
ВВЕДЕНИЕ
Стабильное функционирование природных биогеоценозов (бгц) определяется тем, в какой мере такие системы способны поддерживать необходимый уровень обмена вещества, энергии и информации внутри себя и по отношению к смежным бгц (Ермаков, 2017). Подобный обмен основан на двух противоположно направленных процессах: создание живым веществом первичной продукции и последующее ее разложение. В результате происходит пополнение почвы запасами гумуса и элементами минерального питания.
Синтез органического вещества идет преимущественно на первом трофическом уровне бгц в пределах растительных сообществ, последующие процессы деструкции и минерализации осуществляются другими структурными компонентами. Таким образом, в результате продукционно-деструкционной деятельности “живого вещества” формируются циклы химических элементов, интенсивность которых зависит как от факторов автогенной сукцессии, так и от степени антропогенного воздействия.
В многочисленных работах, связанных с изучением продуктивности фитоценозов, чаще всего учитывается только первая сторона этого процесса. Имеются также исследования, посвященные изучению разложения фитомассы (Андреяшкина, Пешкова, 2001; Курачев, Батурина, 2005; Жуйкова В.А., Жуйкова Т.В., 2010; Воробейчик, Пищулин, 2011 и др.). Комплексные исследования, охватывающие обе стороны продукционного процесса, малочисленны (Анреяшкина, Пешкова, 2003; Жуйкова В.А. и др., 2011; Безель и др., 2016).
Последние годы особая острота проблемы биогеохимического обмена связана с вмешательством в указанные процессы антропогенного фактора, включая химическое загрязнение природных бгц (Ермаков, 2015, 2017; Петрунина и др., 2003; Калабин, Моисеенко, 2011; Безель и др., 2015; Моисеенко, 2017; Ермаков и др., 2018 и др.).
В рамках поставленной задачи рассмотрен вынос ряда химических элементов надземной и подземной фитомассами травяных фитоценозов, подверженных различному уровню антропогенной трансформации. Последующие процессы разложения растительных остатков позволили оценить интенсивность их возврата в биогенный обмен.
Целью работы была оценка интенсивности биогеохимических циклов ряда элементов (Cu2+, Zn2+, Cd2+, Pb2+) в травяных бгц и зависимость этих процессов от эдафических и ценотических факторов среды. Предполагалось, что почвенные условия, включая уровень их загрязнения тяжелыми металлами (ТМ), могут в значительной мере влиять на интенсивность биогенного обмена исследуемых химических элементов, отражая тем самым меру деградации природных бгц, подверженных антропогенному влиянию.
МЕТОДИКА
Характеристика участков исследования. Район исследования – таежная географическая зона, подзона южной тайги (Притагильская часть Среднего Урала, 58° с.ш., 60° в.д.). Интенсивность биогеохимических циклов изучена в четырех фитоценозах, произрастающих на залежах и отвалах промышленных предприятий, относящихся к фоновой и техногенно трансформированным территориям.
Учитывая комплекс физико-химических параметров почв, выделены две их группы: агроземы, техноземы. Участки агроземов расположены в агроландшафтах с агродерново-подзолистыми почвами, со средним плодородием, слабой и средней насыщенностью основаниями (V = 50–95%), низкой и средней обеспеченностью подвижными соединениями фосфора и калия. Содержание легко гидролизуемого азота в агроземах среднее и низкое.
Участки техноземов расположены в техногенных ландшафтах (на промышленных отвалах, возраст которых более 45 лет). Это молодые почвы, формируемые по буроземному и литоземному типам, обладающие более высоким плодородием, сильно насыщенные основаниями (V > 95%), с высокими и очень высокими показателями обеспеченности обменными формами фосфора и калия. Подробная диагностика антропогенно-нарушенных почв выполнена ранее (Кайгородова и др., 2013; Жуйкова и др., 2015; 2019). В соответствие с группой почв участки, на которых произрастают различные травяные фитоценозы обозначены А-1, А-2 (агроземы) и Т-1, Т-2 (техноземы). Данный ряд участков представляет градиент загрязнения почв ТМ, интегральным показателем которого выступает суммарная токсическая нагрузка – Z = = Σ(Ci/Cф) (отн. ед.), где Ci/Cф отношение концентраций элемента на исследуемом участке к фоновому значению. Соответствующие значения Z приведены в табл. 1.
Таблица 1.
Запасы органического вещества в блоках травяных сообществ в градиенте антропогенно преобразованных почв, т/га сух. вес
| Фитомасса | Участки | |||
|---|---|---|---|---|
| агроземы | техноземы | |||
| А-1 (1.0) | А-2 (3.33) | Т-1 (6.19) | Т-2 (22.78) | |
| Максимальная зеленая | 2.67 ± 0.16 | 2.49 ± 0.29 | 2.05 ± 0.27 | 2.30 ± 0.45 |
| Общая надземная* | 4.65 ± 0.28 | 4.33± 0.50 | 3.57 ± 0.47 | 4.00 ± 0.78 |
| Общая подземная** | 3.22 ± 0.23 | 3.15 ± 0.33 | 1.98 ± 0.46 | 1.63 ± 0.33 |
Примечания. * – расчетные значения годовой продукции надземной фитомассы с учетом данных А.А. Титляновой (1977) ** – подземная фитомасса с учетом не идентифицированных остатков. В таблице усредненные данные за 2006, 2009–2012 гг. В качестве меры изменчивости представлена ошибка средней арифметической. В скобках значение Z.
Синтаксономический статус сообществ: А-1 – безранговое сообщество Deschampsia caespitosa-Festuca pratensis [Arrhenatheretalia], А-2 – безранговое сообщество Alchemilla vulgaris–Festuca pratensis [Arrhenatheretalia/Carici macrourae–Crepidetalia sibiricae], Т-1 – безранговое сообщество Carum carvi-Festuca pratensis [Arrhenatheretalia], Т-2 – безранговое сообщество Tussilago farfara–Calamagrostis arundinacea [Dauco–Melilotion/Agropyrion repentis].
Рассматриваемые травяные сообщества являются серийными, формирующимися на залежах и отвалах. На этих территориях их развитие идет в направлении увеличения видового богатства и повышения cуммарного проективного покрытия растений (Жуйкова и др., 2019).
Определение надземной и подземной фитомассы. В качестве меры первичной продукции травяных сообществ была использована величина фитомассы, собранная в период ее максимального развития (Букварева, Алещенко, 2013). В вегетационные сезоны 2006–2012 гг. методом случайной выборки в исследуемых фитоценозах закладывали по 10 учетных площадок размером 25 × 25 см, расположенных на расстоянии друг от друга более чем на 3 метра. Отбор проб проводили методом монолитов с 25 см глубины (Шалыт, 1960). Растения в пределах учетной площадки разбирали по видам. После предварительной подготовки определяли атмосферно-сухую надземную и подземную фитомассы каждого вида (г/м2). Всего выполнено 1370 взвешиваний для надземной фитомассы и 1285 для подземной.
Изучение актуальной скорости разложения растительных остатков проведено в этих же фитоценозах. В качестве экспонируемого материала в полевом эксперименте использовали воздушно-сухую надземную фитомассу трех агро-ботанических групп (бобовые, злаки, разнотравье). Образцы помещали в верхний пяти сантиметровый слой почвы по трансекте через каждые 30 см. На каждой трансекте последовательно закладывали по 10 образцов бобовых, злаков и разнотравья. Период экспонирования образцов – двенадцать месяцев.
После этого срока, извлеченные пакеты, очищали от частиц почвы и тонких корней и высушивали в термостате до абсолютно-сухой массы при температуре 105°С. Скорость разложения экспонируемого материала оценивали по убыли массы навески выраженной в процентах (Воробейчик, 2007).
Скорость разложения подземных частей растений определяли расчетным путем, основываясь на данных Н. В. Переверзева (1987), с учетом того, что годовая скорость разложения надземной фитомассы бобовых в 1.26 раз больше, чем подземной, злаков – в 1.69 раз, разнотравья – в 1.48. Для группы “прочие” рассчитывали среднее между скоростью разложения корней трех агроботанических групп.
Определение содержания тяжелых металлов в почве и растениях. Отбор почвенных образцов и их анализ выполнен по РД 52.18.191-89 в соответствии с аттестованными методами в лаборатории ИЭРиЖ УрО РАН (Аттестат № РОСС RU. 0001.515630). Отбор растительных проб проводили в соответствии с требованиями методик (Методические…, 1992). Экстракцию металлов из почвы проводили 5%-ной НNO3, из растительных образцов – 70%-ной HNO3 или смесью HNO3 и HCl. В кислотных вытяжках измеряли содержание Cu2+, Zn2+, Cd2+, Pb2+ методом пламенной атомно-абсорбционной спектрометрии на спектрометре AAS Vario 6 фирмы Analytik Jena AG. Выполнено элемент-анализов почвы – 200, надземных органов растений – 360, подземных – 235.
Статистический анализ результатов включал вычисление среднего арифметического (М) и его ошибки (m). Нормальность выборочных распределений проверяли с помощью W-теста Шапиро–Уилка, а также через отношение асимметрии (As) к ошибке асимметрии (SAs) и через отношение эксцесса (Ex) к ошибке эксцесса (SEx). Нулевая гипотеза отвергалась, если ${{t}_{{As}}} = \frac{{As}}{{{{S}_{{As}}}}} > {\text{3}}$ и ${{t}_{{Ex}}} = \frac{{Ex}}{{{{S}_{{Ex}}}}} > {\text{3}}{\text{.}}$ Принятие нулевой гипотезы служило допущением к использованию параметрических критериев. Различия между выборками и процент объясненной дисперсии оценивали одно-, двухфакторным дисперсионным анализом с вычислением F-критерия. Групповые сравнения по признакам проведены S-методом Шеффе (методом линейных контрастов).
Сопряженность между признаками оценивали через коэффициент ранговой корреляции Спирмена (Rs), а также с помощью регрессионного анализа (R2). Статистический анализ выполнен в ПСП Statistica v. 10.0 (Stat Soft, Inc., 2012).
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Фитомасса и содержание химических элементов в растениях в условиях антропогенной трансформации почвы. Растительная фитомасса природных бгц – важнейший источник органического вещества в экосистемах и важнейший фактор в формировании биогеохимических процессов. Величина надземной фитомассы (peak standin gcrop), собранной в период ее максимального развития в травяных сообществах, как правило, используется в качестве меры первичной продукции (Титлянова, 1979; Букварева, Алещенко, 2013). Подземная фитомасса отражает ее запасы на момент исследования.
В наших исследованиях травяных фитоценозов была оценена общая надземная и подземная фитомасса, а также содержание ряда химических элементов в органах растений отдельных видов, входящих в состав этих сообществ (Жуйкова, 2009; Безель и др., 2015; 2016). Это позволило оценить запасы химических элементов в блоках исследованных травяных бгц (табл. 1, табл. 2).
Таблица 2.
Вынос химических элементов надземной и подземной фитомассами агроботанических групп травяных фитоценозов
| Участок | Агроботаническая группа | Вынос ионов металлов, мкг/м2 год | |||
|---|---|---|---|---|---|
| Zn2+ | Cu2+ | Pb2+ | Cd2+ | ||
| Надземая фитомасса | |||||
| А-1 | Бобовые | 2553.3 ± 145.6 | 411.5 ± 145.6 | 357.3 ± 126.5 | 11.1 ± 3.9 |
| Злаки | 1711.6 ± 15.1 | 165.5 ± 15.1 | 304.0 ± 27.8 | 13.1 ± 1.2 | |
| Разнотравье | 4499.1 ± 198.9 | 1335.7 ± 198.9 | 868.3 ± 129.3 | 44.4 ± 6.6 | |
| А-2 | Бобовые | 2101.9 ± 49.1 | 372.5 ± 49.1 | 368.8 ± 48.6 | 15.9 ± 2.1 |
| Злаки | 1273.8 ± 38.7 | 224.3 ± 38.7 | 263.1 ± 45.4 | 21.7 ± 3.7 | |
| Разнотравье | 6347.9 ± 218.4 | 1451.6 ± 218.4 | 1891.0 ± 284.5 | 89.5 ±1 3.5 | |
| Т-1 | Бобовые | 5202.7 ± 188.4 | 542.8 ± 188.4 | 480.5 ± 166.8 | 16.4 ± 5.7 |
| Злаки | 2664.8 ± 54.7 | 246.5 ± 54.7 | 387.5 ± 85.9 | 10.2 ± 2.3 | |
| Разнотравье | 9345.2 ± 166.7 | 980.4 ± 166.7 | 1657.5 ± 281.9 | 80.9 ± 13.8 | |
| Т-2 | Бобовые | 5075.3 ± 413.5 | 1642.4 ± 413.5 | 558.3 ± 140.6 | 40.5 ± 10.2 |
| Злаки | 4866.9 ± 460.5 | 1857.0 ± 460.5 | 472.3 ± 117.1 | 33.1 ± 8.2 | |
| Разнотравье | 3916.6 ± 352.2 | 2001.2 ± 352.2 | 1088.8 ± 191.6 | 107.7± 19.0 | |
| Подземная фитомасса | |||||
| А-1 | Бобовые | 2426.3 ± 933.1 | 3274.0 ±1259.0 | 2616.9 ± 1006.3 | 56.9 ± 21.9 |
| Злаки | 5059.3 ± 576.1 | 1743.9 ± 198.6 | 1211.8 ± 138.0 | 379.0 ± 43.2 | |
| Разнотравье | 4936.5 ± 549.9 | 1509.5 ± 168.1 | 631.8 ± 70.4 | 32.9 ± 3.7 | |
| Прочие | 926.9 ± 312.4 | 610.2 ± 205.6 | 197.9 ± 66.7 | 12.4 ± 4.2 | |
| А-2 | Бобовые | 1097.2 ± 286.1 | 231.7 ± 60.4 | 484.4 ± 126.3 | 12.6 ± 3.3 |
| Злаки | 2003.3 ± 657.4 | 4497.2 ± 1475.8 | 640.6 ± 210.2 | 808.4 ± 265.3 | |
| Разнотравье | 5812.9 ± 716.5 | 5513.8 ± 679.6 | 1815.6 ± 223.8 | 1321.2 ± 162.9 | |
| Прочие | 359.1 ± 74.7 | 2488.2 ± 517.3 | 1308.2 ± 272.0 | 199.2 ± 41.4 | |
| Т-1 | Бобовые | 2531.4 ± 731.0 | 1071.5 ± 309.4 | 551.3 ± 159.2 | 42.7 ± 12.3 |
| Злаки | 3581.1 ± 835.7 | 1551.6 ± 362.1 | 314.9 ± 73.5 | 43.3 ± 10.1 | |
| Разнотравье | 7232.0 ± 1973.8 | 2414.0 ± 658.8 | 1296.9 ± 354.0 | 818.0 ± 223.3 | |
| Прочие | 119.5 ± 40.4 | 290.7 ± 98.2 | 935.7 ± 316.2 | 1584.8 ± 535.5 | |
| Т-2 | Бобовые | 4765.3 ± 1646.3 | 12 109.6 ± 4183.5 | 1089.8 ± 376.5 | 24.4 ± 8.4 |
| Злаки | 7614.3 ± 1698.1 | 20 047.3 ± 4470.8 | 994.1 ± 221.7 | 30.0 ± 6.7 | |
| Разнотравье | 5959.1 ± 682.1 | 6454.5 ± 738. 8 | 821.7 ± 94.1 | 50.9 ± 5.8 | |
| Прочие | 3318.4 ± 1412.1 | 5722.6 ± 2435.2 | 272.2 ± 115.8 | 10.5 ± 4.5 | |
Деструкция растительных остатков. В ходе исследования определена актуальная скорость разложения бобовых, злаков и разнотравья на участках с различной степенью антропогенной трансформации (табл. 3).
Таблица 3.
Количество разложившихся растительных остатков в градиенте антропогенной трансформации почвы (% за год)
| Агроботанические группы |
Агроземы | Техноземы | ||
|---|---|---|---|---|
| А-1 | А-2 | Т-1 | Т-2 | |
| Б | $\frac{{32.59 \pm 2.13}}{{10}}$ | $\frac{{23.53 \pm 3.77}}{{10}}$ | $\frac{{41.87 \pm 0.54}}{3}$ | $\frac{{48.79 \pm 1.23}}{{10}}$ |
| З | $\frac{{27.81 \pm 1.90}}{{10}}$ | $\frac{{30.59 \pm 2.59}}{{10}}$ | $\frac{{41.93 \pm 2.67}}{5}$ | $\frac{{45.29 \pm 3.81}}{8}$ |
| Р | $\frac{{36.86 \pm 2.08}}{8}$ | $\frac{{33.62 \pm 2.44}}{{10}}$ | $\frac{{55.34 \pm 5.29}}{3}$ | $\frac{{54.49 \pm 1.54}}{9}$ |
| Среднее | 32.10 ± 1.33 | 29.24 ± 1.87 | 48.47 ± 1.29 | 35.81 ± 2.14 |
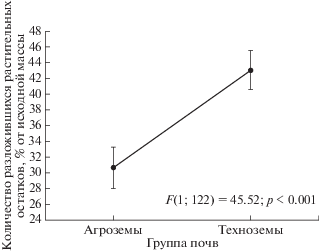
Проверка выборочных распределений на соответствие закону нормального распределения позволила принять нулевую гипотезу и использовать методы параметрической статистики. Зависимость среднего количества разложившихся растительных остатков (у) от уровня токсической нагрузки на почвах различного типа (х) аппроксимируется уравнением линейной регрессии: у = = (29.41 ± 2.05) + (2.68 ± 0.63)х (R2 = 0.12; p < 0.001). В целом, количество разложившихся растительных остатков на участках техноземов за период экспонирования, выше, чем на участках агроземов, что подтверждено однофакторным дисперсионным анализом (рис. 1) и S-методом Шеффе (F = 5.08; df = 3; 116; p = 0.001).
Рис. 1.
Результаты однофакторного дисперсионного анализа различий в количестве разложившихся растительных остатков на агроземах и техноземах за год.
Таким образом, интенсивность вовлечения ТМ в биогенный цикл определяется двумя параметрами: накоплением их в фитомассе и возвратом в биогенный обмен в результате деструкции и минерализации растительных остатков. Описание нами подобных процессов является лишь некоторым приближением к реальному кругообороту химических элементов, поскольку не учитывает разложение растительного материала предшествующих лет. Полученные оценки следует рассматривать лишь в качестве некоторых интегральных характеристик общего состояния бгц, в том числе меры его антропогенной трансформации.
Согласно данным многих авторов (Медведева и др., 2006; Узбек, 2006; Санданова, 2007; Нетрусов, Котова, 2009; Лисецкий, 2012; Головацкая, Никонова, 2013; Käärik, 1974; Global-scale similarities… , 2007) интенсивность деструкции растительных остатков определяется главным образом химическим составом разлагающихся образцов, временным фактором и почвенными условиями (составом почвенных микробиоценозов, содержанием органических компонентов в почвах, уровнем загрязнения их ТМ и пр.).
Полученные результаты проанализированы в ходе двухфакторного (“группа почв” и “агроботаническая группа”) ковариационного анализа с “токсической нагрузкой (Z)" в качестве ковариаты объема разложившихся растительных остатков. Для участков с различными эдафическими условиями, включая загрязнение их ТМ, показано, что с увеличением срока экспонирования существенно возрастает вклад фактора "группа почв”: от 3.55% после первых двух месяцев до 25.20% к концу первого года. При этом вклад фактора “агроботаническая группа” уменьшается за этот период в 3 раза (рис. 2).
Рис. 2.
Вклад факторов “группа почв” (1) и “агроботаническая группа” (2) в разложение растительных остатков.
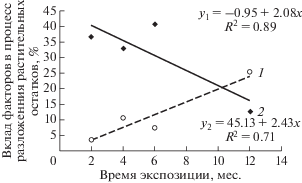
Следовательно, на начальных этапах разложения (первые 6 мес.) значительно влияние структуры и химического состава разлагаемого растительного материала. К окончанию первого года определяющими становятся эдафические условия. В нашем случае, вероятно, физико-химические особенности почв, включая их загрязнение ТМ.
Зависимость возврата ТМ от их концентрации в почвах. Содержание подвижных форм ТМ в почвах в существенной мере определяет как уровень их накопления фитомассой, так и качественный и количественный состав почвенных деструкторов (Курачев, Батурина, 2005; Узбек, 2006; Семенова и др., 2011; Ившина и др., 2014).
Данные таблиц 1, 2, позволяют оценить количество химических элементов ежегодно возвращающихся в биогенные циклы за счет разложения надземной и подземной фитомасс. На рисунке 3 приведены результаты анализа вовлечения в биогенный обмен Zn, Cu, Pb и Cd надземной и подземной фитомассами травяных сообществ, ежегодный их возврат в ходе разложения растительных остатков, а также ежегодная доля такого возврата.
Рис. 3.
Ежегодный вынос ТМ надземной и подземной фитомассами, возврат и доля возврата их в биогенный цикл при разложении растительных остатков: 1 – агроземы, 2 – техноземы.
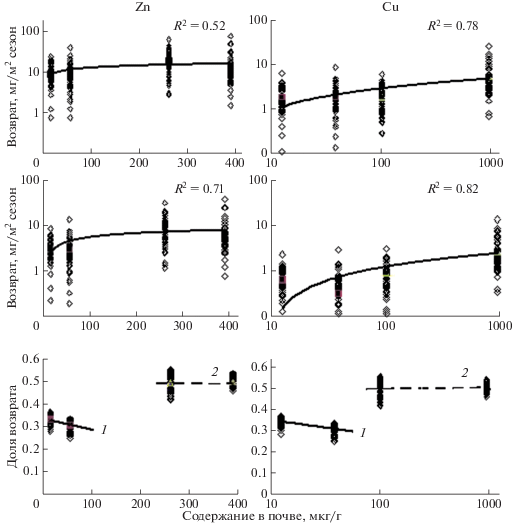
Рис. 3.
Окончание
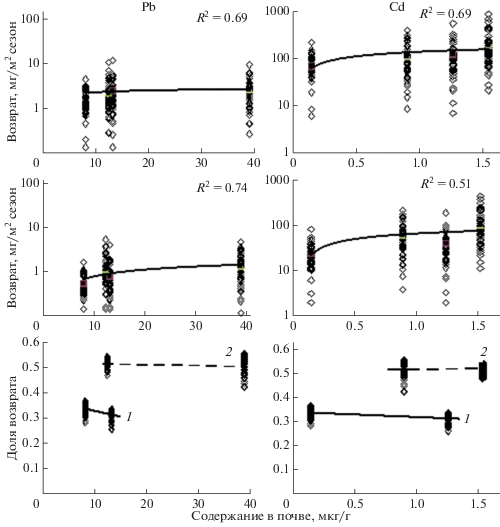
Полученные данные показывают четкую зависимость выноса ТМ надземной и подземной фитомассой от концентрации соответствующих химических элементов в почвах, а также уровень их возврата в биогеохимические циклы за счет деструкции и последующей минерализации растительных остатков. При увеличении концентрации цинка в почвах в 20 раз его вынос фитомассой и последующий возврат в биогеохимический цикл возрастает только в 1.5–2.0 раза. При загрязнении почвы медью более чем в 75 раз вынос и возврат этого металла возрастает соответственно в 3–8 раз. При нашем незначительном градиенте загрязнения почвы свинцом в 1.5 раза его вынос практически не меняется, но возврат в биогенный цикл возрастает в 2.5 раза. Из наиболее токсичных элементов концентрация в почвах кадмия возрастает в 10 раз, а вынос и его возврат в биогенный обмен возрастает лишь в 3–4 раза.
Эти данные противоречат некоторым опубликованным, согласно которым скорость деструкции опада в почвах, подверженных загрязнению ТМ и соединениями серы снижается (Воробейчик, 2002; Reduction of decomposition rate …, 1991 и др.). С другой стороны, на загрязненных почвах процессы разложения растительного материала могут протекать быстрее, чем на фоновых (Помазкина, 2011). Возможно, при высоких концентрациях ТМ в почвах возрастает роль устойчивых к загрязнению деструкторов (бактерий и грибов), что ведет к ускорению процессов разложения (Ившина и др., 2014).
В большей степени интенсивность биогеохимических циклов характеризуется отношением выноса ТМ фитомассой растений к уровню их возврата в геохимический обмен. Полученные данные показали четкое различие этого показателя на участках агроземов и техноземов (рис. 3). Для всех исследуемых элементов доля их возврата в биогенный обмен на агроземах ниже, чем на техноземах, даже при относительно низких концентрациях свинца в почвах на участке Б-2. Эти данные особо интересны, поскольку показывают, что минеральный обмен в травяных фитоценозах определяется в нашем случае не только уровнем загрязнения почвы ТМ. Важную роль играют фитоценотические факторы, а также эдафическая специфика условий существования растительного сообщества.
Роль агроботанических групп в обмене ТМ в травяных бгц. В многочисленных исследованиях показана видовая специфичность накопления ТМ надземными и подземными органами растений. Наши данные также свидетельствуют, что в течение первых 6 месяцев экспозиции растительных остатков в почвах определяющим является состав разлагающихся агроботанических групп (рис. 2). Соотношение фитомассы агроботанических групп “разнотравье–бобовые–злаки” изменяется в градиенте техногенной трансформации среды (%): надземная от 46.8 : 21.3 : 31.9 на участке А-1 до 17.7 : 19.1 : 63.2 на Т-2; подземная – от 45 : 23 : 32 до 21 : 17 : 62, соответственно.
Поскольку доля разложившейся в течение года фитомассы агроботанических групп различна на исследуемых участках (табл. 3), то их участие в биогенных циклах определяется не только объемом синтезированной фитомассы, но и скоростью ее разложения. На рисунке 4 представлена доля ежегодного возврата этих элементов в биогенный обмен надземными и подземными органами растений различных агроботанических групп. При низких концентрациях ТМ в почвах максимальный вклад в биогеохимический обмен почти всех исследуемых элементов вносят виды разнотравья, на долю которых в наших условиях приходится от 30 до 80% общего обменного пула Cu2+, Zn2+, Pb2+. Вклад злаков и бобовых меньше (от 15 до 30%). Исключение составляет Cd2+, возврат которого в биогенный обмен в условиях низкого содержания этого элемента в почвах осуществляется преимущественно злаками.
Рис. 4.
Доля участия бобовых (1), злаков (2), разнотравья (3) в возврате ТМ в процессе разложения их растительных остатков.
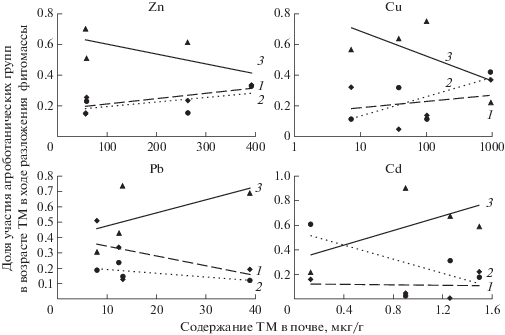
С увеличением содержания Zn2+ и Cu2+ в почвах участие разнотравья в биогенных циклах снижается, а злаков и бобовых возрастает. С другой стороны, в градиенте наших условий участие разнотравья в биогенном обмене Pb2+ и Cd2+ возрастает за счет снижение доли бобовых и злаков.
Таким образом, различные агроботанические группы, входящие в состав изученных травяных фитоценозов, в силу их неодинаковой представленности в общей надземной и подземной фитомассах, а также в силу различного химического состава их тканей, с различной интенсивностью участвуют в обмене химических элементов.
Роль микробиологического комплекса почв. Важнейшим компонентом эдафического статуса почвы, в конечном счете, определяющим и интенсивность циклов биогеохимического обмена, является состав и обилие почвенной микрофлоры. Выполненные ранее микробиологические исследования позволили установить качественное и количественное различие микробиологического комплекса на рассмотренных нами участках (Ившина и др., 2014).
Важную роль в процессе разложения растительных остатков играют анаэробные и аэробные целлюлозолитические бактерии, численность которых положительно коррелирует с уровнем загрязнения почвы ТМ. Коэффициенты ранговой корреляции Rs равны соответственно для аэробов 0.65–0.74; анаэробов и мезофиллов 0.79; термофилов 0.60 (N = 15; р < 0.05–0.001). При этом общая численность почвенных микроорганизмов на загрязненных территориях выше, чем на фоновой.
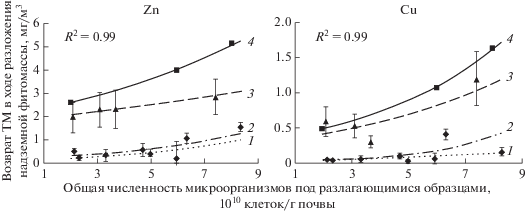
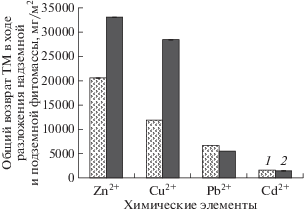
Таким образом, установлено, что возврат цинка и меди в биогенный обмен возрастает при увеличении общей численности микроорганизмов. Максимальное влияние обилия микроорганизмов в почве на возврат ТМ проявляется при разложении разнотравья (рис. 5). Влияние этого фактора на разложение двух других агроботанических групп менее выражено. Поскольку высокая интенсивность этих процессов отмечена при наиболее высоком общем обилии микроорганизмов на техноземах (участки Т-1 и Т-2) по сравнению с агроземами (участки А-1 и А-2), то, возможно, в этих условиях формируется упрощенная структура микробного сообщества с преобладанием r-стратегов. С этим связана и высокая метаболическая активность микроорганизмов (Ившина и др., 2014). Высокая общая численность в составе микробиоценоза целлюлозоразрушающих бактерий определяет высокие темпы разложения растительных остатков на техноземах и больший по сравнению с агроземами возврат химических элементов в биогенные циклы. Максимально это проявляется в случае цинка и меди (рис. 6).
ЗАКЛЮЧЕНИЕ
Участие химических элементов в биогеохимических циклах определяется сложным комплексом взаимосвязанных биоценотических и эдафических характеристик природных бгц, включая их возможную антропогенную трансформацию.
Данные о выносе ТМ надземной и подземной фитомассами травяных фитоценозов в ходе синтеза первичного органического вещества и последующем их возврате в процессе деструкции растительных остатков и минерализации органики позволили сравнить интенсивность биогеохимического обмена ряда эссенциальных (цинк, медь) и токсических (свинец, кадмий) элементов в природных биогеоценозах, подверженных различному уровню антропогенной трансформации.
Показано, что в условиях антропогенно преобразованных ландшафтов Среднего Урала интенсивность биогеохимического обмена ТМ зависит не только от содержания в почвах подвижных форм химических элементов, выступающих в роли загрязнителей природной среды, но в существенной мере определяется совокупностью ценотических и эдафических условий. Наличие в травяных фитоценозах видов разнотравья способствует более интенсивному накоплению ТМ в растительной фитомассе, а последующее их интенсивное разложение, активному возврату ионов в биогенный цикл. На интенсивность обменных процессов в природных бгц активно влияет также состав и обилие почвенной микрофлоры, в том числе численность целлюлозоразрушающих микроорганизмов.
Полученные данные свидетельствуют о том, что в рассмотренных нами уровнях загрязнения почвы ТМ интенсивность биогенных циклов в значительной мере зависит от степени антропогенной трансформации почвы, связанной не только с прямым влиянием ТМ, но в значительной мере от обилия и соотношения в растительном сообществе агроботанических групп, а также от степени развития почвенного микробоценоза.
Список литературы
Андреяшкина Н.И., Пешкова Н.В. (2001) К оценке темпов разложения растительного опада и стандартных образцов целлюлозы в тундровых сообществах. Экология (1), 57-60.
Андреяшкина Н.И., Пешкова Н.В. (2003) К характеристике продукционного и деструкционного процессов в равнинных и горных тундрах Крайнего Севера. Экология (2), 108-114.
Безель В.С., Жуйкова Т.В., Гордеева В.А. (2015) Геохимия травянистых биогеоценозов: биогенные циклы химических элементов при загрязнении природной среды тяжелыми металлами. Геохимия (3), 252-263.
Bezel’ V.S., Zhuikova T.V., Gordeeva V.A. (2015) Geochemistry of grass biocenoses: Biogenic cycles of chemical elements at contamination of the environment with heavy metals. Geochem. Int. 53(3), 241-252.
Безель В.С., Жуйкова Т.В., Гордеева В.А., Мелинг Э.В., Трубянов А.Б., Глотов Н.В. (2016) Надземная фитомасса и скорость деструкции растительных остатков в травянистых сообществах при загрязнении почвы тяжелыми металлами. Экология (4), 264-269.
Букварева Е.Н., Алещенко Г.М. (2013) Принцип оптимального разнообразия биосистем. Товарищество научных изданий КМК, 522 с.
Воробейчик, Е.Л. (2002) Изменение пространственной структуры деструкционного процесса в высотном и токсичных градиентах: природно-техногенные аналоги. Экологические проблемы горных территорий (Под ред. Васильева А.Г.). Екатеринбург: Академкнига, 224-232.
Воробейчик Е.Л. (2007) Сезонная динамика пространственного распределения целлюлозолитической активности почвенной микрофлоры в условиях атмосферного загрязнения. Экология (6), 427-437.
Воробейчик Е.Л., Пищулин П.Г. (2011) Влияние деревьев на скорость деструкции целлюлозы в почвах в условиях промышленного загрязнения. Почвоведение (5), 597-610.
Головацкая Е.А., Никонова (Абзалимова) Л.Г., Головацкая Е.А. (2013) Разложение растительных остатков в торфяных почвах олиготрофных болот. Вестник Томского гос. ун-та. Биология 3(23), 137-151.
Ермаков В.В. (2015) Геохимическая экология и биогеохимические критерии оценки экологического состояния таксонов биосферы. Геохимия (3), 203-221.
Ермаков В.В. (2017) Концепция биогеохимических провинций А.П. Виноградова и ее развитие. Геохимия (10), 875-890.
Ermakov V.V. (2017) A.P. Vinogradov’s Concept of Biogeochemical Provinces and Its Development. Geochem. Int. 55(10), 872-886.
Ермаков В.В., Тютиков С.Ф., Дегтерев А.П., Сафонов В.А., Данилова В.Н., Хушвахтова С.Д., Гуляева Ю.А., Кречетова Е.В. (2018) Биогеохимическая дифференциация живого вещества и биоразнообразия в условиях Адонского полиметаллического субрегиона биосферы. Геохимия (4), 336-350.
Ermakov V.V., Tyutikov S.F., Degtyarev A.P., Safonov V.A., Danilova V.N., Khushvakhtova S.D., Gulyaeva U.A., Krechetova E.V. (2018) Biogeochemical Differentiation of Living Matter and Biodiversity in the Ardon Polymetallic Subregion of the Biosphere. Geochem. Int. 56(4), 318-331.
Жуйкова Т.В. (2009) Реакция ценопопуляций и травянистых сообществ на химическое загрязнение среды. Автореф. дис. … д-ра биол. наук. Екатеринбург: Институт экологии растений и животных УрО РАН, 40 с.
Жуйкова В.А., Жуйкова Т.В. (2010) О скорости разложения растительного опада в луговых сообществах техногенно нарушенных территорий. Современные проблемы биомониторинга и биоиндикации (Под ред. Ашихмина Т.Я., Алалыкина Н.М. и др.). Киров: ООО “Лобань”, Ч. 2, 162-166.
Жуйкова В.А., Волохина К.С., Волохин И.С., Жуйкова Т.В. (2011) Две стороны продукционного процесса в луговых сообществах техногенно нарушенных территорий. Экология: сквозь время и расстояние. Екатекринбург: Гощицкий, 73-82.
Жуйкова Т.В., Мелинг Э.В., Кайгородова С.Ю., Безель В.С., Гордеева В.А. (2015) Особенности почв и травянистых растительных сообществ в условиях техногенеза на Среднем Урале. Экология (3), 163-172.
Жуйкова Т.В., Безель В.С., Бергман И.Е., Мелинг Э.В., Кривошеева А.В. (2019) Фертильность и жизнеспособность пыльцевых зерен Taraxacum officinale Wigg. s.l. (Asteraceae, Magnoliopsida) в градиенте антропогенно трансформированной среды. Поволжский экологический журн. (3), 275-290.
Ившина И.Б., Костина Л.В., Каменских Т.Н., Жуйкова В.А., Жуйкова Т.В., Безель В.С. (2014) Почвенный микробиоценоз как показатель стабильности луговых сообществ при химическом загрязнении среды тяжелыми металлами. Экология (2), 83-90.
Кайгородова С.Ю., Жуйкова Т.В., Жуйкова В.А., Безель В.С., Ившина И.Б. (2013) Характеристика антропогенно-деградированных почв и микробного комплекса территорий Нижнее-Тагильского промузла. Современные проблемы загрязнения почв (Под ред. Ладонина Д.В., Мотузовой Г.В.). М.: МГУ, 226-230.
Калабин Г. В., Моисеенко Т. И. (2011) Экодинамика техногенных провинций горнопромышленных производств: от деградации к восстановлению. ДАН 437(3), 398-403.
Курачев В.М., Батурина В.Б. (2005) Темпы разложения растительных остатков в почвах техногенных ландшафтов. Сибирский экологический журн. (5), 789-793.
Лисецкий Ф.Н. (2012) Особенности трансформации растительного вещества степных экосистем. Фундаментальные исследования. Биологические науки (3), 245-249.
Медведева М.В., Бахмет О.Н., Яковлев А.С. (2006) Процессы деструкции опада ели в условиях аэрогенного загрязнения. Лесоведение (4), 75-77.
Методические указания по определению тяжелых металлов в почвах сельхозугодий и продукции растениеводства (изд. 2-е, перераб. и доп.) (1992) (Подгот.: Кузнецов Н.А. и др.). М.: ЦИНАО, 126.
Моисеенко Т.И. (2017) Эволюция биогеохимических циклов в современных условиях антропогенных нагрузок: пределы воздействия. Геохимия (10), 841-862. Moiseenko T.I. (2017) Evolution of biogeochemical cycles under anthropogenic loads: Limits impacts. Geochem. Int. 55(10), 841-860.
Нетрусов А.И., Котова И.Б. (2009) Общая микробиология: учебник для студентов биолог. специальностей ун-тов (3-е изд.). М.: Академия [Academia], 352 с.
Переверзев Н.В. (1987) Биохимия гумуса и азота почв Кольского полуострова. М.: Наука, 306 с.
Петрунина Н.С., Ермаков В.В. Дегтярева О.В. (2003) Геохимическая экология растений в условиях полиметаллических биогеохимических провинций. Тр. Биогеохим. лаб. (24). М.: Наука, 226-253.
Помазкина Л.В. (2011) Интегральная оценка влияния техногенного загрязнения и климатических факторов на агросистемы Байкальской природной территории. Успехи современной биологии 131(3), 194-203.
Санданова И.Б. (2007) Микробиологическая деструкция растительного опада степных экосистем Юго-Восточного Забайкалья. Автореф. дис. … канд. биол. наук. Улан-Уде, Бурятской гос. ун-т, 20 с.
Семенова И.Н., Ильбулова Г.Р., Суюндуков Я.Т. (2011) Изучение эколого-трофических групп почвенных микроорганизмов в зоне влияния горнорудного производства. Фундаментальные исследования. Биологические науки (11), 410-414.
Титлянова А.А. (1977) Биологический круговорот углерода в травяных биогеоценозах. Новосибирск: Наука, Сибирское отделение, 219 с.
Титлянова А.А. (1979) Биологический круговорот азота и зольных элементов в травянистых биогеоценозах. Новосибирск: Наука, Сиб. отд-ние, 149 с.
Узбек И.Х. (2006) Целлюлозоразрушающие микроорганизмы как компонент биологического фактора почвообразования. Екологія та ноосферологія 17(1-2), 11-16.
Шалыт М.С. (1960). Вегетативное размножение и возобновление высших растений и методы его изучения. Полевая геоботаника (Под ред. Корчагина А.А. и др.). М.; Л. (2), 163-208.
Parton W., Silver W.L., Burke I.C., Grassens L., Harmon M.E., Currie W.S. (2007) Global-scale similarities in nitrogen release patterns during long-term decomposition [Electronic resource] Science 315(5810), 361-364.
Käärik A.A. (1974) Decomposition of wood. Biology of plant litter decomposition (Dickinson H., Pugh J. F. (eds). N.Y.: Academic Press (1), 129-174.
Berg M.P., Ekbohm G., Soderstrom B., Staaf H. (1991) Reduction of decomposition rate of Scots pine needle litter due to heavy-metal pollution [Electronic resource] Water, Air and Soil Pollution (59), 165-177.
Дополнительные материалы отсутствуют.