

Геохимия, 2021, T. 66, № 11, стр. 1035-1041
Механизм эволюционного упорядочения биологических систем (в свете теории академика Э.М. Галимова)
Т. И. Моисеенко *
Институт геохимии и аналитической химии им. В.И. Вернадского РАН
119991 Москва, ул. Косыгина, 19, Россия
* E-mail: moiseenko.ti@gmail.com
Поступила в редакцию 17.03.2020
После доработки 20.03.2021
Принята к публикации 22.03.2021
Аннотация
Рассматривается универсальность концепции Э.М. Галимова в объяснении механизма эволюции биологических систем. Показано, что концепция позволяет дать объяснение не только зарождению жизни в первично абиогенном мире, но и биологическому развитию в системах надорганизменного уровня организации живого – в экосистемах и биосфере в целом. Дается понимание общих закономерности формирования экосистем и приводятся ключевые характеристики их стабильности, при которых система минимизирует рассеяние энергии. Приводятся воззрения Э.М. Галимова на эволюцию современного мира, как фазу вовлечения в упорядочение огромных масс вещества, не принадлежащих собственно к органическому миру.
ВВЕДЕНИЕ
Э.М. Галимов в 2001 г. публикует первое издание монографии “Феномен жизни: между равновесием и нелинейностью. Происхождение и принципы эволюции”, которая посвящена одной из наиболее фундаментальных проблем естествознания – проблеме происхождения жизни и законам эволюции. В книге формируется представление о возможных путях зарождения жизни на планете и ее развития, управляющими механизмами которых являются законы неравновесной термодинамики. Его работа еще два раза была переиздана и вызывала большой интерес у научного мира. После публикации концепции возникновения жизни А.И. Опарина (1959), работа Э.М. Галимова была наиболее яркая и научно доказательная работа, дающая понимание возможных путей зарождения жизни и возникновения генетического кода.
Генетический код рассматривается как опосредованный автокатализ полипептидов через нуклеотидные последовательности и показывает, что действенным соединением, которое обладает уникальными свойством осуществлять функцию соответствия между структурными формами нуклеиновых оснований и структурными формами аминокислот является t-РНК (Галимов, 2008). Он отмечает, что концепция эволюционного упорядочения позволяет рассматривать в одинаковом ключе не только механизм зарождения жизни, но и собственно биологическое развитие – эволюцию.
Наиболее очевидный и яркий феномен жизни состоит в высокой плотности упорядочения живого вещества и последовательном возрастании упорядочения в ходе эволюции. Эволюционное упорядочение обеспечивается сочетанием производства низкоэнтропийного продукта и процессов итерации (Галимов, 2005, 2008).
Э.М. Галимов (2008) показывает, что дарвинская теория не объясняет полностью эволюционный процесс, он пишет: “дарвинизм правильная теория, но в силу ряда причин, является недостаточной для объяснения происхождения жизни и ее эволюционного усложнения” (с. 28). Формулирует 6 ключевых принципов эволюции, с позиций которых объясняет движущую силу эволюции. “В основе биологической эволюции лежит механизм естественного упорядочения. Источником упорядочения являются диспропорционирование энтропии в сопряженных процессах”. “Эволюция упорядочения обеспечивается сочетанием производства низкоэнтропийного продукта с итеративным его воспроизводством Упорядочение может осуществляться на микроскопическом уровне. Процесс репликации выводит микроскопическое упорядочение на макроскопический уровень” (с. 78). Мерой упорядочения он считает функциональное соответствие, а не фенотипическую полезность. В работе подчеркивается значимая роль в эволюции комбинаторного видообразования, горизонтального переноса генов в “генном резервуаре биосферы”. Он подчеркивает, что “биологическая эволюция представляет сочетание восходящей ветви, связанной с усложнением и упорядочением и нисходящей, которая характеризует процессы, ведущие к адаптации и видовому разнообразию, равновесие является аттрактором” (с. 197).
Следует отметить, что использование энергетического критерия для объяснения основных тенденций в эволюции биосферы было предпринято и в ряде других предшествующих работ (Хильми, 1966; Завадский, 1959; Шварц, 1980; Колчинский, 1990; Ervin, 1991). Важнейшая термодинамическая характеристика организмов, экосистем и биосферы в целом – состояние низкой энтропии, т.е. способность создавать и поддерживать высокую степень упорядоченности вещества (Пригожин, Стенгерс, 2008). В этой работе мы приведем примеры, которые подтверждают универсальность концепции упорядочения вещества Э.М. Галимова (2005, 2008) и рассмотрим на примере экосистем развитие к их устойчивому стабильному состоянию в открытых неравновесных системах, каковыми являются экосистемы и биосфера в целом.
ЕСТЕСТВЕННОЕ УПРОРЯДОЧИВАНИЕ ЭКОСИСТЕМ
Известно, что в ходе эволюции биосферы была выработана оптимальная организация, связанная с особенностями использования ассимилированной энергии на различных уровнях жизни (Шварц, 1980; Stearns, 1992). В работе Ю. Одума (1986) обоснована общая закономерность сукцессий экосистем, которые в своем развитии проходят ряд стадий, достигающих устойчивую стадию своего состояния в данных условиях. При развитии экосистем характерен целый ряд общих закономерностей развития: постепенное увеличение видового разнообразия, смена доминирующих видов, усложнение цепей питания, увеличение в сообществах доли видов с длительными циклами развития, усиление взаимовыгодных связей в биоценозах и т.д. Структура экосистемы формируется под влиянием адаптации организмов к условиям конкретной среды и максимальному использованию энергетических ресурсов в экологических нишах. Эволюция реализуется через экологические взаимоотношения, в которые вступают все организмы, а ее механизмы – те же самые, которые постоянно поддерживают упорядоченность структуры и функционирования экологических систем” (Гиляров, 2003).
Стабильность (стационарность) есть функция устойчивости экосистемы к действию дестабилизирующего фактора, будь то природные катаклизмы или стрессовые условия, каковым является загрязнение окружающей среды. Стабильность может характеризоваться как структурная (поддержание основной структуры при изменении функции), так и функциональная (сохранение основных функций в измененной структуре) (Алимов, 1994). На примере загрязняемых заливов крупных озер были рассмотрены основные модификации водных экосистем при прохождении их через критическое состояние в период сильного загрязнения и восстановительной сукцессии при сокращении загрязнения. Было показано, что система приобрела новые свойства стабильности и возврата к прежним характеристикам не произойдет (Moiseenko, Sharov, 2019). В последующей работе были рассмотрены модификации экосистем в условиях загрязнения, их эволюция объясняется в свете теории Э.М. Галимова (Моисеенко, 2020).
Э.М. Галимов (2008), анализирую феномен жизни, вводит следующие определения: живые системы – это итеративные стационарные системы необратимых процессов, протекающих в области соотношений сил и потоков энергии. Стационарность живых систем обеспечивается минимумом производства энтропии. Система стремиться организовываться таким образом, чтобы в ней возникли необратимые процессы линейной передачи энергии, которые препятствовали нарастанию энтропии и свели это нарастание к минимуму. Устойчивость биологических систем обусловлена их существенной линейностью, способностью к воспроизводству и тем, что они являются не просто сложными, а представлены подсистемными множествами. Рассмотрим, в какой степени эти положения (необходимые условия стабильности) могут дать понимание механизмов устойчивого функционирования экосистем.
Соотношения сил и потоков энергии. В природных условиях, если приход энергии в экосистему равен расходу с учетом рассеяния, то состояние экосистемы остается стабильным во времени. Приток вещества и энергии (в виде биогенных и органических веществ, солнечной радиации, характерной для региона) постоянен в сезонных циклах. Образование продукции (Р) в пределах годовых флуктуаций, уравновешено тратами на дыхание (R), т.е. P/R ≈ 1. Биодоступные формы биогенных элементов и веществ утилизированы в экосистеме, поступающие извне – потребляются, избыток – выводится путем захоронения или стока. Если приход превышает расход, система приспосабливается за счет возрастания численности организмов, обеспечивающих экосистеме более полное использование энергии. Это может выглядеть как усложнение. Наконец, если возникает дефицит поступающей энергии (расход превышает приход), то система упрощается (Одум, 1986; Алимов, 2000).
Необратимость и сопряжение процессов осуществляется путем восходящих (от продуцентов к консументам 1, 2, 3…. i-го порядков) и нисходящих (редуцентами) потоков передачи энергии и вещества, т.е. выполняется важное условие стационарности – осуществляется путем последовательности передачи энергии внутри открытых систем, каковыми являются экосистемы (от одних подсистем к другим), при котором диссипация энергии минимальна. Процессы продукции и деструкции вещества в подсистемах (организмы, сообщества) протекают сопряжено таким образом, что продукция одной подсистемы является ресурсом для другой, формируя упорядоченную структуру. Вместе с тем Э.М. Галимов (2008) отмечал, в биологических системах всегда присутствуют элементы нелинейности и обратимости в подсистемах или на отдельных отрезках времени, не изменяя общий ход необратимости в циклах “жизни”.
Функциональное соответствие, как одно из условий стационарности, достигается таким количеством видов и особенностями их функционирования в трофической структуре экосистемы, которые минимизируют рассеивание энергии (как в восходящих, так и нисходящих линиях передачи энергии в пищевых цепях). Разнообразие каждой из подсистем (сообществ) будет определяться функциональным соответствием видов как внутри, так и между видами, так и внутри сообществ: продуценты–консументы, хищник–жертва, хозяин–паразит и др.
Итеративность процессов. Воспроизводство членов (индивидуумов разных видов) в сообществах определяет итеративность всего сообщества. Итеративность (воспроизводство) энергозатратный механизм. Э. Пьянка (1998) сформулировал теорию о стратегиях жизненного цикла видов (особей) в стабильных и изменчивых условиях. В постоянно действующих стабильных условиях в подсистемах преимущество для выживания получают крупные долгоживущие виды с редким циклом воспроизводства (K-стратеги), которым требуется больше энергии на поддержание метаболизма и обеспечение роста, накопления резервов для относительно редкого цикла воспроизводства. В изменчивых условиях с успехом выживают более мелкие формы (r-стратеги) с частой сменой поколений (Pianka, 1970). Воспроизводство – энергозатратный механизм, поэтому долгоживущие виды с более редким циклом воспроизводства (итеративности) минимизируют в той или иной степени рассеяние энергии. При условиях действия деструктивного (стрессового) фактора, например, токсичного загрязнения, преимущество для выживания получают более мелкие формы. Данное явление в деталях рассмотрено на примере изменчивости экосистем озер северо-запада в период загрязнения и восстановительной сукцессии (Moiseenko, Sharov, 2019).
Низкая энтропийность. Структурная сложность экосистем определяется таким образом, чтобы минимизировать потери (рассеяние) энергии при передаче из одной подсистемы в другую. Она зависит от ряда природных абиотических факторов, таких как климат, ландшафтные характеристики и др., которые определяют приток энергии в экосистему и ее отвод. Более сложно организованные экосистемы формируются в условиях высокого потока энергии и вещества, поэтому биоразнообразие закономерно возрастает от северных к более южным широтам, где потоки энергии возрастают. Биоразнообразие и увеличение количества сопряженных процессов обеспечивает снижение диссипации энергии и приводит к формированию наиболее низкоэнтропийной системы, как основного признака стабильности.
А.Ф. Алимов (2000) измеряемыми параметрами стабильности сообщества или экосистемы в конкретных регионах предлагает принимать пределы варьирования их характеристик, не выходящих за рамки среднего уровня годовых флуктуаций, сложившихся в ходе эволюции и свойственных данной системе. По мнению Э.М. Галимова (2008) – аттрактором стабильного состояния является состояние системы с минимальным рассеянием энергии и минимумом производства энтропии (рис. 1).
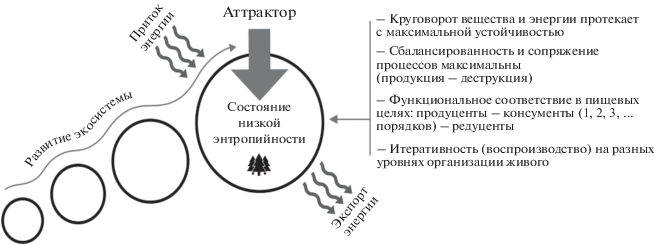
ЧЕРЕЗ КРИТИЧЕСКОЕ СОСТОЯНИЕ К САМООРГАНИЗАЦИИ
“Стационарные системы обладают высокой устойчивостью и гибкостью, пока выполнены два обязательных условия стационарности: постоянство притока энергии и обмена веществом. Нарушение этих условий закрывает путь формирования низкоэнтропийных структур и система гибнет” (Галимов, 2008, с. 221). Термин “гибель” может быть применен к процессам, происходящим на молекулярном, клеточном и организменном уровнях, на экосистемном – протекают процессы дезорганизации.
Когда деструктивный фактор, каковым является любой стрессовый фактор, включая природные и антропогенные катаклизмы, воздействует на экосистему, нарушается ее целостность и стабильность, увеличивается диссипация энергии. Преобразования в природе развиваются сложными путями в постоянном взаимодействии процессов различной направленности – изменчивости и поддержания стабильности.
И. Одум (1986) сформулировал концепцию “энергетических субсидий”, под которой понимает всякий источник энергии, увеличивающий ту долю энергии, которая может пойти на продукцию. К ним он относит умеренный поток органических и биогенных веществ в экосистему, однако, высокий и нерегулярный их приток он рассматривает как фактор стресса. Токсичное загрязнение – только как фактор стресса, приводящий к усиленной диссипации энергии и деградации экосистемы. Феномен дополнительного неупорядоченного рассеяния энергии экосистемы в критическом состоянии (при ее реорганизации или деградации) в современной экологии получил статус “закона” (Шварц, 1980; Одум, 1986; Колчинский, 1990; Алимов, 2000). С позиций энергетического критерия под воздействием деструктивного фактора происходит реорганизация экосистемы, нарушаются пути передачи энергии в трофической структуре и увеличивается энтропия.
Приведем основные признаки критического состояния экосистем (или на стадии развития), как высоко энтропийной (хаотичной) системы, которая эволюционирует в новое состояние. В условиях действия деструктивного фактора (будь то поток загрязнения, неумеренное изъятие продукции, вырубка или пожары в лесах) снижается биоразнообразие за счет уничтожения отдельных видов, возрастных когорт или наиболее чувствительных к действию загрязнения видов. Относительно высокая численность некоторых устойчивых видов поддерживается большим количеством энергии, которая поступает в экосистемы извне и не используется в разбалансированной экосистеме и/или высвобождается вследствие разрушенных путей ее передачи в исходной исторически сложившейся экосистеме (Moiseenko, Sharov, 2019). Таким образом происходит нарушение сбалансированности потоков вещества и энергии.
Среди видов близкой толерантности преимущества в конкуренции получают мелкие формы (r-стратеги) с более быстрой частотой воспроизводства и сменой поколений (Pianka, 1970). Долгоживущие виды К-стратегии более уязвимы к действию стрессовых условий (например, токсичных агентов), поскольку вероятность физиологических нарушений на протяжении более продолжительного периода их жизни высокая (Большаков, Моисеенко, 2009). Вследствие появления мелких форм с коротким жизненным циклом в каждой из подсистем происходит ускорение итеративности всей системы.
На фоне снижения количества чувствительных видов или снижения их обилия в экосистеме возрастает численность нескольких эврибионтных видов, т.е. происходит резкое увеличение их доминантности (Моисеенко, 2011). С одной стороны, вследствие элиминации наиболее чувствительных видов снижается конкуренция, с другой – способные выживать эврибионтные виды получают больше энергетических субсидий и имеют преимущества для роста и размножения. Это в свою очередь ведет к упрощению разнообразия системы и “разрывам” путей передачи энергии и ее диссипация.
Структурные перестройки, связанные с действием деструктивного фактора приводят к возникновению новой сети взаимодействий между компонентами. При вторжении новых единиц, если они успешно приживаются и размножаются в достаточном количестве, то вся система перестраивается на новый режим функционирования (Пригожин, Стенгерс, 2008), что мы наблюдаем на примере экосистем в критическом состоянии, например, в экосистемах озер северных регионов (Moiseenko, Sharov, 2019).
Снижение неупорядоченного рассеяния энергии может достигаться не только за счет усложнения структуры, но и ускорения оборота биомассы, которое обеспечивается короткоцикловыми и более мелкими видами (или особями одного вида). Признаки критического состояния экосистем (в стадии усиления диссипации энергии) носят одновременно черты противодействия хаосу и нарастания энтропии, поэтому могут рассматриваться в качестве адаптивной стадии реорганизации экосистемы, стремящейся к упорядочению вещества. Уменьшение размеров членов сообщества (доминирование мелких форм устойчивых видов) и частая их итерация, которые наблюдаются в загрязняемых условиях, обеспечивают более эффективную утилизацию энергии, поступающей в виде энергетических субсидий или высвобождающуюся вследствие нарушения ее передачи в трофической структуре экосистемы. Переход через критическое состояние (точку или последовательность точек бифуркации) приводит к необратимой изменчивости экосистем, как показано в работе Т.И. Моисеенко (2020).
Критическое состояние под влиянием дезорганизующего фактора запускает механизм упорядочения вещества, аттрактором которого является минимизация диссипации энергии. Во многих случаях довольно сложно провести черту между понятиями “хаос” и “порядок”. Система может эволюционировать, то приближаясь, то удаляясь от равновесного состояния. Неравновесность – движущая сила эволюции (или необратимых преобразований) которая приводит к изменению потоков энергии при стремлении к равновесию эволюционирующих открытых систем. Необратимость существует на всех уровнях, однонаправлена во времени и играет конструктивную роль в формировании новой структуры (Галимов, 2008).
ЭВОЛЮЦИЯ СОВРЕМЕННОГО МИРА
Загрязнение окружающей среды предоставило уникальную модель наблюдать эволюцию в действии (Бигон и др., 1998). Галимов (2008) дает свое понимание о вступлении биосферы в фазу антропогенного мира. Он избегает термина “ноосфера” и характеризует современный период как фазу вовлечения в упорядочивание огромных масс вещества, не принадлежащих собственно к органическому миру – загрязняющие отходы производства, влияние выбросов на климат, техногенные катастрофы и т.д. Антропогенно-обусловленные процессы способны оказывать влияние и на устойчивость биосферы в целом, невероятно ускоряя эволюционный процесс, создавая “эволюционные вихри” “К биосфере приложимы те же принципы эволюции, которые сформулированы в отношении эволюции жизни: это должна быть стационарная система, обеспеченная притоком энергии, возвращение которой в стационарное состояние после возмущающих событий должно порождать низкоэнтропийный продукт” (Галимов, 2008, с. 125). “В биосфере возникают внутренние процессы, масштаб которых соизмерим с масштабами – пространственными, энергетическими, информационными, характеризующими биосферу в целом” (Галимов, 2008, с. 205).
В основе концепции биологического упорядочения – процесс, происходящий в стационарной системе в линейной области зависимости сил и потоков, при этом Э.М. Галимов (2008, с. 162) отмечает, что “линейность, как и равновесие является некой идеализацией. Высокая упорядоченность биологических систем есть результат выработанной эволюцией специфической структуры взаимодействий. Наряду с процессами упорядочения всегда протекают в той или иной степени процессы дезинтеграции”. Если рассматривать биосферу как континуум экосистем на суше или на морских пространствах, то изменчивость будет представлять волны возмущения на том или ином отрезке, система в целом будет развиваться, балансируя между равновесием и нелинейностью при необратимом векторе эволюции.
В работе (Моисеенко, 2017) показаны масштабы нашего влияния на биогеохимические циклы. Активированные человеческой деятельностью потоки элементов и веществ вызывают развитие негативных процессов, изменяют биогеохимические циклы и приводят к эволюционным изменениям органического мира, как необратимого процесса. Изменения средообразующих факторов под воздействием загрязнения приводят к изменчивости индивидуумов, популяций и экосистем, которые развиваются по восходящей линии передачи энергии и информации, протекают в единстве и взаимосвязанности сквозь уровни организации живого. Регуляция может идти в обратном порядке.
Большая группа ученых (Rockstrom et al., 2009) проанализировала исторические эпохи кризисных состояний и предприняли попытку определить допустимые (граничные) нагрузки по ключевым антропогенным факторам, т.е. определить запас способности экосистем и биосферы “усваивать” антропогенно-индуцированные потоки элементов и веществ. По мнению ученых, в настоящее время превышен порог допустимости концентраций СО2 в целом на планете, который, растворяясь в воде, вызывает снижение насыщения вод арагонитом. Был обоснован критерий снижения насыщения океанических вод арагонитом, как значение 350 ррmv и ниже. Основываясь на обобщении обширной литературы Rockstrom et al. (2009) предсказали, что увеличение концентрации СО2 выше этого критического значения в морской воде: из-за высокие значения в атмосфере могут привести к появлению следующих эффектов в целом на планете: произойдет перераспределение растительности на поверхности Земли и сдвиг фенологических фаз, соответственно, изменится облик окружающей среды, который сохранял относительную стабильность в течение голоцена, изменится населенность территорий; будет происходить оттаивание ледяного покрова и вечной мерзлоты. Обоснование критического порога допустимого повышения содержания СО2 основывается на палеоклиматических данных и необходимости сохранности ледяного покрова Арктического океана, Антарктики, Гренландии и вечной мерзлоты Сибири. Доказано, что последние 50 млн лет относительно низкая концентрация СО2 была основным охлаждающим фактором на планете, при концентрации более 450 ррmv в прошлом (>100 млн лет назад) поверхность Земли была свободна от льда. С повышением парниковых газов связано изменение радиационного баланса, который по оценкам ученых превышен. Критически нарушен цикл азота, хотя резерв по фосфору имеет еще быть место.
Приведенные примеры свидетельствуют, что в “биосфере возникают внутренние процессы, масштаб которых соизмерим с масштабами – пространственными, энергетическими и информационными, характеризующие биосферу в целом”, которые были обозначены в работе Э.М. Галимовым (2008, с. 205). Возможно ли восстановить нарушенные или человечеством преобразованные экосистемы до природных показателей и есть ли необходимость в этом? Мы показали в работах (Большаков, Моисеенко, 2009; Моисеенко, 2017, 2020; Moiseenko, Sharov, 2019), что после разрушающего действия загрязнения экосистемы не восстанавливаются, эволюционируют в новое состояние.
Дискуссионный вопрос поднимается рядом ученых – должны ли мы стремиться воссоздать природное состояние (Сairns, 2005; Palmer et al., 2005; Folk et al., 2006) и насколько это оправдано. Знание траектории сукцессий сообществ и экосистем в условиях увеличения и снижения антропогенного давления, позволяет предсказать будущие изменения и правильно направить практические усилия на ускорение процессов восстановления. Palmer et al. (2005) обосновывает основную цель восстановления – дизайн экологического восстановления должен учитывать имиджу природного объекта и быть направлен на поддержание основных показателей структуры и функций экосистемы, сохранения жизнеобеспечивающих условий на планете.
ЗАКЛЮЧЕНИЕ
Теория естественного эволюционного упорядочения вещества Э.М. Галимова объясняет механизм зарождения жизни и ее эволюционное развитие. Главное свойство живых организмов – выживать и воспроизводиться в пределах, которые обеспечиваются энергетическими ресурсами экологических ниш, определяют экологические законы поддержания стабильности экосистем и их преобразования в условиях дезорганизующего фактора (природные катаклизмы или антропогенное загрязнение) и находятся в соответствии с термодинамическими законами упорядочения вещества в живых системах.
Современный мир Э.М. Галимов (2008) характеризует как фазу вовлечения в упорядочения огромных масс вещества, не принадлежащих собственно к органическому миру – загрязняющие отходы производства, влияние выбросов на климат, техногенные катастрофы и т.д. В биосфере возникают локальные изменения на уровне отдельных экосистем или их континуума. Признаками критического состояния экосистем являются: увеличение энергетических затрат на поддержания биомассы; повышение роли поступающей извне энергии в экосистемы; ускорение оборота биогенных элементов; разрывы путей передачи энергии и вещества в трофической структуре; увеличение пропорции мелких форм (виды r-стратегии); снижение видового разнообразия и увеличение видовой доминантности, снижение биоразнообразия. При этом нарушается одно из главных условий стабильности – функциональное соответствие внутри подсистем, которое приводит к неупорядоченному рассеянию энергии.
В то же время “хаос” явление детерминистическое и в этом состоянии система стремится к минимизации рассеяния энергии, например, мелкие особи с коротким жизненным циклом обеспечивают более активное использованию поступающей извне энергии, частая итеративность которых (воспроизводство) поддерживает биомассу. Ускорение процессов итерации (воспроизводства) за счет мелких форм и видов является механизмом снижения энтропии. Исходя из экологической теории и термодинамических законов упорядочения энергии в биологических системах, ведущим механизмом стабильности вновь эволюционирующей экосистемы и биосферы в целом будут являться ее новые свойства, направленные на уменьшение диссипации энергии.
Бесспорно, огромные массы техногенных выбросов, вовлекаемые в биогеохимический круговорот, вырубка лесов и вовлечение земель в сельскохозяйственное использование меняют облик нашей Земли. Эволюционное развитие идет согласно законам упорядочения вещества и минимизации энтропии по траектории дальнейшей эволюции, поэтому возврата к прежним условиям невозможно, как предсказано теорией Э.М. Галимова. Адаптация к изменяющимся условиям должна быть направлена на поддержание основных показателей структуры и функций экосистем, удовлетворяющих потребности человека в продуктах питания, рекреации и эстетического потенциала.
Работа выполнена в рамках Государственного задания № 0137-2019-0008.
Список литературы
Алимов А.Ф. (2020) Элементы теории функционирования водных экосистем. СПб.: Наука. 148 с.
Бигон М., Харпер Дж., Таунсенд К. (1989) Экология. Особи, популяции и сообщества. М.: Мир, Т. 1, 667 с.; Т. 2, 447 с.
Большаков В.Н., Моисеенко Т.И. (2009) Антропогенная эволюция животных: факты и их интерпретация. Экология 40 (5), 323-332.
Галимов Э.М. (2005) Предпосылки и условия возникновения жизни. Задачи исследования. Геохимия (5), 467-484.
Galimov E.M. (2005) Prerequisites and conditions of the origin of life: bjectives for research. Geochem. Int. 43(5), 421-437.
Галимов Э.М. (2009) Феномен жизни:Между равновесием и нелинейностью. Происхождение и принципы эволюции. М.: Книжный дом “ЛИБРОКОМ”, 269 с.
Гиляров А.М. (2003) Становление эволюционного подходя как объяснение началa в экологии. Журнал общей биологии 64 (1). С. 3-22.
Завадский К.М. (1959) К пониманию прогресса в органической природе. Проблема развития в природе и обществе. М.–Л., С. 78-120.
Колчинский Э.И. (1990). Эволюция биосферы. Л.: Наука. 236 с.
Моисеенко Т.И. (2011) Устойчивость водных экосистем и их изменчивость в условиях токсичного загрязнения. Экология 42 (6), 441-448.
Моисеенко Т.И. (2017) Эволюция биогеохимических циклов в современных условиях антропогенных нагрузок: пределы воздействий. Геохимия (10), 841–862.
Moiseenko T.I. (2017) Evolution of Biogeochemical Cycles under Anthropogenic Loads: Limits Impacts. Geochem. Int. 55 (10), 841-860.
Моисеенко Т.И. (2020). Эволюция экосистем в условиях антропогенных нагрузок: через дезорганизацию к самоорганизации. Геохимия 65 (10), 939-948.
Moiseenko T. I. (2020) Evolution of Ecosystems under an Anthropogenic Load: From Disorganization to Self-Organization. Geochem. Int. 58 (10), 1083-1091.
Одум Ю. (1986) Экология (Пер. с англ. / Под ред. Гилярова А.М.). М.: Мир, 328 с.
Пианка Э.P. (1981) Эволюционная экология. (Пер. с англ. / Под ред. Гилярова А.М.): М.: Мир, 399 с.
Одум Ю. (1986) Экология. (Пер.с англ. / Под ред. Гилярова А.М. М.): Мир, 1986. Т. I. 328 с.
Опарин А. И. (1959) Происхождение жизни. М.: Военное издательство Министерства обороны СССР, 1959. 268 с.
Пригожин И., Стенгерс И. (2008) Порядок из хаоса. Новый диалог человека с природой (Пер. с англ. / Под ред. Аршинова В.И., Климонтовича Ю.Л., Сачкова Ю.В.) М., 269 с. Хильми Г.Ф. (1966) Основы физики биосферы. Л. Гидрометеоиздат. 1966г. 300с.
Шварц С.С.(1980) Экологические закономерности эволюции. М.: Наука, 1980. 278 с.
Cairns J. Jr. (2005) Restoration Ecology and Ecotoxicology. Handbook of Ecotoxicology (Eds Hoffman). N.Y.: Levis publishers Ltd., 2005. P. 1015-1029.
Erwin T. A. (1991) An Evolutionary basis for conservation strategies. Science, 253, 750-752.
Falk D.A., Palmer M., Zedler J. (2006) Foundation of restoration ecology. The Science and Practice of Ecological restoration. Washington: Island Press, 2006. 518 p.
Moiseenko T, Sharov A. (2019) Large Russian Lakes Ladoga, Onega, and Imandra under Strong Pollution and in the Period of Revitalization: A Review. Geosciences 9, 492.
Palmer M.A., Ambrose R.F., Poff N. (2007) Ecology theory and community restoration. Restoration Ecology. 5(4), 291-300.
Pianka E.R. (1970) On r- and K- selection. American Naturalist, 104, 592-597.
Rockström J., Steffen W., Noone K., Persson Å., Chapin F.S., Lambin E.F., Lenton T.M., Scheffer M. (2009) A safe operating space for humanity. Nature (461), 472-475.
Stearns S. C. (1992) The Evolution of life History. Oxsford: Oxford University 249 p.
Дополнительные материалы отсутствуют.