

Вопросы ихтиологии, 2019, T. 59, № 2, стр. 175-185
Воспроизводство тихоокеанских лососей (Oncorhynchus) в реках, впадающих в залив Терпения (Сахалин)
А. А. Живоглядов 1, 2, *, Л. А. Живоглядова 1
1 Азовский научно-исследовательский институт рыбного хозяйства − АзНИИРХ
Ростов-на-Дону, Россия
2 Сахалинский научно-исследовательский институт рыбного хозяйства – СахНИРО
Южно-Сахалинск, Россия
* E-mail: tauroskiff@mail.ru
Поступила в редакцию 24.01.2017
После доработки 02.06.2018
Принята к публикации 29.10.2018
Аннотация
Оценены условия воспроизводства тихоокеанских лососей рода Oncorhynchus в реках, впадающих в зал. Терпения. Описаны различия между реками северной и западной частей исследуемого района по морфологии русла, фаунистическому составу водной биоты и преобладающему виду лососей. В реках северного участка (бассейн р. Поронай и оз. Невское), имеющих как горно-предгорные, так и равнинные типы русел, воспроизводство горбуши O. gorbusha находится на низком уровне. В реках западного участка, имеющих выраженный горный характер и сравнительно бедную ихтиофауну, сосредоточено воспроизводство горбуши зал. Терпения. Приведены данные по динамике численности покатников и производителей. Величина пропуска производителей горбуши в реки зал. Терпения, необходимая для устойчивого воспроизводства, оценена в пределах 3.63−10.42 млн экз.
В конце XIX−начале XX вв. зал. Терпения являлся одним из основных промысловых районов Сахалина (Шмидт, 1905). В реках и на побережье залива добывали исключительно лососей рода Oncorhynchus (преимущественно кету O. keta), тогда как другие промысловые районы Сахалина были местами смешанного, сельдево-лососёвого, промысла.
История изучения пресноводной ихтиофауны и тихоокеанских лососей бассейна зал. Терпения довольно продолжительная (Таранец 1937а, 1937б; Линдберг, 1953; Воловик и др., 1972; Гриценко и др., 1987; Никифоров, Гришин, 1989; Сафронов, Никифоров, 1995; Никифоров и др., 1997; Сафронов и др., 2000; Гриценко, 2002). В силу большой рыбохозяйственной значимости вопрос регулирования промысла лососей и оптимального заполнения нерестовых рек Сахалина производителями затрагивали в разное время многие исследователи (Таранец, 1939; Леванидов, 1964; Воловик, 1967; Рухлов, 1968; Риккер, 1971; Воловик и др., 1972; Голованов, 1982; Гриценко и др., 1987, 1989; Ким, Антонов, 2002; Ефанов, 2003; Антонов, 2005). Ретроспективный анализ работ по данной проблеме приведён в работе Макеева (2010), который отмечает, что величина заполнения, рекомендуемая исследователями в качестве оптимальной для рек Сахалина, варьирует в довольно широких пределах (0.3–2.0 экз/м2 нерестовой площади), различаются и подходы к определению величины заполнения. Так, одни исследователи (Wickett, 1958; Иванков, Андреев, 1972) считают, что при оптимальной величине заполнения должен обеспечиваться наибольший выход молоди с единицы площади нерестилищ, другие (Семко, 1939; Таранец, 1939; Рухлов, 1968) отталкиваются от средней площади нерестового гнезда лососей и площади нерестилища, необходимой для одной самки. Нам представляется наиболее чётко сформулированным определение Гриценко с соавторами (1987. С. 116), которые за оптимальную принимают величину, “ниже которой выживание (икры и личинок) уменьшается из-за слабой их (нерестилищ) мелиорированности, а превышение оптимума снижает выживаемость вследствие перекопок ранее устроенных бугров и попадания мёртвой икры в гнёзда к живой”. При этом авторы указывают, что для рек, имеющих различные характеристики и протекающих по разным географическим районам, значение оптимума будет отличаться.
К настоящему моменту в СахНИРО накоплен большой объём материалов о сроках подходов, величине и динамике уловов, биологических показателях тихоокеанских лососей Сахалинской области, включая зал. Терпения (Каев, 2008, 2009; Каев и др., 2010). Несмотря на значительный массив данных, приведённых в указанных публикациях, они имеют преимущественно описательный характер и относятся к отдельно взятым видам. Сравнительно подробно выполнен анализ приуроченности крупных стад кеты к основным артезианским бассейнам и водоносным комплексам Сахалина (Иванков, 1993; Каев, 2001). Для горбуши O. gorbusha попытки анализа связей между продуктивностью нерестилищ и абиотическими условиями предприняты на ограниченном материале (Каев, 2007, 2011а; Живоглядов и др., 2013). Между тем для эффективного управления промыслом очевидна необходимость анализа экосистемных связей тихоокеанских лососей в многовидовом аспекте с привлечением данных как по биотической, так и по абиотической составляющей среды (Шунтов, 2010, 2016; Фельдман и др., 2016). Без этого трудно понять причину различий в потенциале воспроизводства разных промысловых районов и рек разных типов, выделить закономерности и перейти к обоснованному определению ёмкости нерестовых площадей и величин заполнения нерестовых рек Сахалина. Такая ситуация осложняет управление промыслом и напрямую влияет на точность прогнозирования величины рекомендуемого вылова лососей (Золотухин, 2003; Заварина, 2008; Шунтов, 2010).
Цель работы − на основании многолетних данных мониторинга тихоокеанских лососей побережья зал. Терпения, материалов по морфологии, гидрологии, донным сообществам и составу ихтиофауны текучих вод выявить места наиболее эффективного воспроизводства массовых видов лососей, описать основные фоновые характеристики, определить численность производителей, необходимую для устойчивого воспроизводства в реках данного района.
МАТЕРИАЛ И МЕТОДИКА
В основу статьи положены данные, собранные при непосредственном участии авторов, а также фондовые материалы СахНИРО и Сахалинрыбвода за 1978−2014 гг. Все работы проведены в соответствии с принятыми в Сахалинской области методами (Инструкция …, 1987; Каев, 2011б; Методические рекомендации …, 2013). Данные по статистике промысла лососей ставными неводами собраны в районе г. Поронайск и в западной части побережья зал. Терпения (прибрежье Макаровского городского округа). Учёт мигрирующих производителей в реках Поронай, Лазовая, Лесная, Макарова и определение эффективности нереста лососей (выживаемость особей эмбрионально-личиночных стадий, численность пополнения) выполнены на базе Смирныховской, Поронайской и Макаровской контрольно-наблюдательных станций и биостанции СахНИРО. В некоторые годы помимо пеших обходов для учёта производителей проводили аэровизуальное обследование.
Статистический анализ данных выполнен стандартными методами (Плохинский, 1961; Правдин, 1966; Лакин, 1980).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Фоновая характеристика района исследований. Побережье зал. Терпения простирается от м. Терпения на востоке до м. Тихий на западе. Акватория восточного побережья Сахалина находится под сильным влиянием холодного Восточно-Сахалинского течения, вследствие чего разрушение ледового покрова у берегов зал. Терпения начинается довольно поздно − в I декаде мая (Ресурсы …, 1973; Гриценко и др., 1987). Исследуемый район в орографическом отношении можно разделить на две части (рис. 1): северный участок, включающий бассейны р. Поронай и оз. Невское и прилегающие реки (от м. Терпения до р. Гастелловка), и западный участок, включающий реки Макаровского городского округа (до р. Тихая).
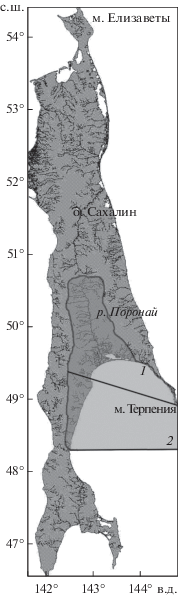
Северный участок расположен в пределах Тымь-Поронайской низменности, заполненной аллювиальными и морскими отложениями и имеющей средневысотный горно расчленённый рельеф, который прорезан рядом широких аккумулятивно-эрозионных террас (Атлас …, 1967). Тымь-Поронайская низменность представляет собой плоскую, интенсивно заболоченную равнину протяжённостью 300 км при ширине от 5−6 км на севере до 90 км на юге. Наиболее значимым водотоком бассейна зал. Терпения является Поронай – самая протяжённая река Сахалина (350 км), во многом определяющая облик местных водных систем. Этот водоток берёт начало на высоте 820 м на западных склонах горы Невельского (Восточно-Сахалинские горы) и протекает по широкой заболоченной долине между Восточно-Сахалинскими и Западно-Сахалинскими горами (Ресурсы …, 1973). Общая площадь нерестилищ в бассейне Пороная составляет 4.65 км2, в том числе 1.76 км2 в основном русле Пороная, а 2.89 км2 распределены по притокам, среди которых наиболее значимы Житница и Орловка; подавляющее большинство нерестилищ являются горбушёвыми (Гриценко и др., 1987). На северном побережье зал. Терпения восточнее устья р. Поронай расположено самое большое на Сахалине оз. Невское (площадь водного зеркала 178 км2). С северной стороны в озеро впадают крупные реки Рукутама и Оленья (Атлас …, 1967; Сафронов и др., 2008), которые имеют большое значение для воспроизводства горбуши, кеты и кижуча O. kisutch (суммарная площадь нерестилищ около 1.22 км2). Общая площадь нерестилищ северного участка (включая 0.33 км2 в малых реках, не относящихся к бассейнам Пороная и оз. Невское) составляет 6.2 км2.
Рельеф западного участка определяется Западно-Сахалинскими резко расчленёнными горами (хребтами Камышовым, Западно-Лисянским, Жданко и другими), является низкогорным и относится к альпинотипному классу (Казаков, Генсиоровский, 2007). Реки западного побережья зал. Терпения (Макаровский городской округ) имеют значительно меньшую протяжённость по сравнению с Поронаем и относятся к горно-предгорному типу. Долины рек узкие, уклоны русел значительные. Грунты галечные, в верхнем течении – крупногалечные, на порожистых участках преобладают валуны и скальные обнажения (Рухлов, 1982; Гриценко и др., 1987). Самая протяжённая река данного участка – Макарова (длина 97 км, площадь бассейна 589 км2) – берёт начало на западном склоне Западно-Лисянского хребта, в северной его части. Ширина наиболее крупных рек составляет 40−60 м, малых рек не превышает 40 м. Речная сеть развита хорошо (коэффициент густоты речной сети 1.81 км/км2) и по территории распределена сравнительно равномерно. Средневзвешенные уклоны рек Макарова, Нитуй, Лазовая и Гастелловка составляют 5.2−9.9‰ (Казаков, Генсиоровский, 2007). Общая площадь нерестилищ западного участка – 1.3 км2. Суммарная нерестовая площадь рек зал. Терпения составляет около 7.5 км2.
Основу донных сообществ ритрали рек Сахалина формируют амфибиотические насекомые (Лабай и др., 2015). Для участков горного типа в верховьях притоков Пороная характерен видовой комплекс с доминированием Hexatoma sp. (15 таксонов). В среднем течении притоков преобладает сообщество с доминированием веснянок Skwala compacta (36 видов и групп беспозвоночных). Характерным для приустьевых участков является летний комплекс бокоплавов Eogammarus kygi, осенью сменяющийся на комплекс E. barbatus, насчитывающий до 58 таксонов донных беспозвоночных (Живоглядова и др., 2012).
Ихтиофауна бассейна р. Поронай (Тымь-Поронайский фаунистический район) представлена 33 видами рыб и рыбообразных из 12 семейств (Никифоров и др., 1997; Сафронов, Никифоров, 2003). Рыбы, населяющие реки и озёра бассейна р. Поронай, имеют разное происхождение. В целом для бассейна Пороная отмечено значительное преобладание бореального комплекса с добавлением древнего верхнетретичного (Сафронов, 2000). Ихтиофауна Пороная имеет ряд черт, сближающих её с ихтиофауной р. Амур, на основании этого Никифоров с соавторами (1997) полагают, что в относительно недавнем геологическом прошлом бассейн Пороная являлся частью бассейна Палеоамура.
В верховье Пороная основу рыбных сообществ составляют ручьевая форма и молодь проходной формы мальмы Salvelinus malma, кунджа S. leucomaenis и молодь симы O. masou (Никифоров и др., 1997). Основу сообществ рыб равнинной части русла р. Поронай составляют представители семейств карповых (Cyprinidae) и лососёвых (Salmonidae). В весенний период в сообществах рыб Пороная доминируют пресноводные (амурский язь Leuciscus waleckii, амурская щука Esox reichertii) и проходные виды (обыкновенная малоротая корюшка Hypomesus olidus, крупночешуйная краснопёрка Tribolodon hakonensis, сахалинская краснопёрка T. ezoe), летом по массе и численности превалируют тихоокеанские лососи (в основном, горбуша и кета), в осенний период практически на всём протяжении равнинной части русла Пороная доминирует типично пресноводный вид − L. waleckii (Живоглядов и др., 2011).
Ихтиофауна западного участка (юго-восточный фаунистический район) включает 25 видов рыб и рыбообразных (Никифоров, 2001). На значительном протяжении рек горно-предгорного типа встречаются всего два−четыре вида рыб. В пределах горной зоны ручьёв (общая протяжённость русла менее 10 км) и малых рек (с длиной русла не более 100 км) доминирует ручьевая форма мальмы; субдоминанты сообщества – молодь симы и кунджа. Предгорную зону малых рек населяет сообщество кунджи и сибирского гольца Barbatula toni с тремя субдоминантами − сахалинским тайменем Parahucho perryi, подкаменщиком Cottus sp. и сахалинской девятииглой колюшкой Pungitius tymensis. Предгорный участок ручьёв населён сообществом сибирского усатого гольца; субдоминанты – сахалинская девятииглая колюшка, китайская колюшка P. sinensis, молодь симы и дальневосточных краснопёрок, пресноводный дальневосточный бычок Gymnogobius urotaenia (Живоглядов, 2014).
Тихоокеанские лососи. В прибрежье зал. Терпения мигрируют четыре вида тихоокеанских лососей − горбуша, кета, кижуч и сима. Наиболее ранняя анадромная миграция отмечена для симы: её производители заходят в реки во второй половине мая, нерестовый ход продолжается до начала августа. Горбуша, воспроизводящаяся в реках зал. Терпения, имеет сложную популяционную структуру и представлена тремя популяционными (темпоральными) группировками − япономорской (наиболее ранней по срокам хода), тихоокеанской (охотоморской) летней и тихоокеанской (охотоморской) осенней (Гриценко и др., 1989). Вследствие этого сроки нерестового хода горбуши зал. Терпения весьма продолжительны – с июня до середины сентября. Первые гонцы япономорской группировки горбуши в бассейне р. Поронай и прилегающих реках появляются рано, начало нерестового хода отмечено в I−II декадах мая. Данную темпоральную группу горбуши нередко отмечают в реке вместе с первыми производителями симы. В нечётные годы ход япономорской группировки начинается раньше, чем в чётные, что справедливо и для других темпоральных группировок горбуши. Япономорская группировка является самой малочисленной. Рунный ход и, соответственно, наибольшие уловы горбуши зал. Терпения приходятся на конец июля − I декаду августа (период летней межени), основой рунного хода и промысла является осенняя группировка. Завершается нерестовый ход горбуши в сентябре, в период осеннего повышенного стока. В реках, впадающих в зал. Терпения, воспроизводятся две формы кеты – летняя (малочисленная в современный период) и осенняя (в последние годы численность находится на сравнительно высоком уровне). Летняя форма кеты мигрирует в Поронай с I декады июня по III декаду августа, осенняя − с конца августа по конец октября. Позднее других видов на нерест идёт кижуч, его производители подходят к рекам в октябре−ноябре.
Скат молоди горбуши с рек бассейна зал. Терпения длится с конца мая по начало июля, при этом большая часть молоди обычно скатывается в две первые декады июня. Сроки массового ската мальков горбуши варьируют, но в основном миграция начинается в начале мая и заканчивается в конце июня. Скат молоди кеты происходит с начала мая по начало августа (Гриценко и др., 1987). Покатники симы отмечаются с первых чисел июля по начало августа, серебрянки кижуча − с конца июня до начала августа (Никифоров и др., 1997).
Промысел тихоокеанских лососей в прибрежье зал. Терпения ведут преимущественно ставными и закидными неводами. С 1876 по 1885 гг. на Сахалине вылавливали 2000−3000 т лососей в год, к 1900-му г. уловы увеличились до 4240 т, в дальнейшем по мере роста населения интенсивность промысла возрастала (Шмидт, 1905). В 1899−1901 гг. зал. Терпения занимал первое место в лососёвом промысле, здесь промышляли в основном кету – до 87% общего вылова данного вида на Сахалине. Уловы горбуши в зал. Терпения были примерно равны уловам на Юго-Восточном Сахалине − соответственно 45 и 43% общего вылова этого вида (Рухлов, 2007).
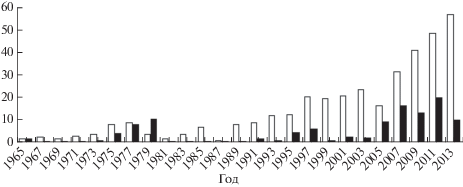
Численность горбуши подвержена периодическим колебаниям, особенно велика разница между поколениями чётных и нечётных лет (Воловик, 1967; Рухлов, 1982; Гриценко и др., 1989). В анализируемый период (1965−2014 гг.) на побережье зал. Терпения в нечётные годы вылавливали от 0.590 до 56.604 тыс. т, в чётные − от 0.003 до 19.583 тыс. т рыбы. Средний объём вылова горбуши в нечётные годы (14.39 тыс. т) превышал таковой в чётные (4.32 тыс. т) в 3.3 раза (рис. 2).
Рис. 2.
Динамика уловов горбуши Oncorhynchus gorbusha в зал. Терпения в чётные (◼) и нечётные (◻) годы (1965−2014), тыс. т.
Горбуше р. Поронай (северный участок) присуще чередование периодов доминирования особей нечётных лет с периодами, когда доминирование исчезает во время подъёмов численности поколений чётных лет (Гриценко и др., 1987). С 1980-х гг. по настоящее время практически постоянно преобладают поколения нечётных лет, рост численности поколений чётных лет существенно меньше. Площадь нерестилищ западного участка по сравнению с северным в 4.7 раза меньше (1326 против 6206 тыс. м2), при этом его доля в общем вылове горбуши на акватории зал. Терпения в 2000−2914 гг. была выше: в нечётные годы на его побережье приходилась половина (до 52.7%), а в чётные – почти 3/4 (до 70.7%) уловов.
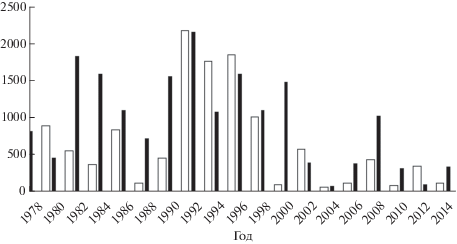
Численность мигрирующей молоди до 2002 г. рассчитывали по наиболее продуктивным притокам Пороная (ручьи Березовый и Холодный, левобережные притоки I порядка), что приводило к существенному завышению оценок численности пополнения. После 2002 г. в дополнение к этому начали оценивать объём ската молоди из менее продуктивного правобережного притока (р. Орловка) и определять общую величину пополнения на основании осреднённых данных (по правобережным и левобережным притокам), а также привлекать данные по рекам западного побережья залива (Каев, 2011а). В результате этого итоговые оценки численности мигрировавшей в море молоди снизились и стали более реалистичными (рис. 3). В период 2003−2014 гг. урожай молоди горбуши линии чётных лет варьировал в пределах 73.50−1481.06 (в среднем 513.80) млн экз., нечётных − 66.1−575.8 (252.14) млн экз., т.е. в нечётные годы численность пополнения была в два раза меньше.
Рис. 3.
Численность покатной молоди горбуши Oncorhynchus gorbusha чётных и нечётных поколений, мигрировавшей из рек бассейна зал. Терпения в 1978−2014 гг., млн экз.; обозначения см. на рис. 2.
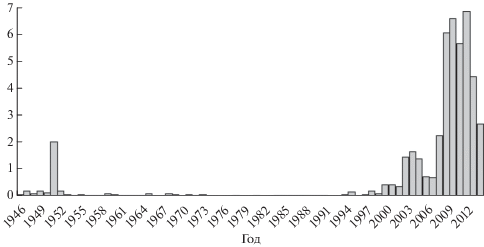
Считается, что амурская и сахалинская летняя кета имеют общее происхождение, это представление основывается на том, что реки о-ва Сахалин в прошлом входили в систему Палеоамура (Лапшина и др., 2012). В настоящее время осенняя форма кеты преобладает по численности над летней и в Амуре, и в Поронае, тогда как ещё в 1950-е гг. летняя форма была гораздо более многочисленной (Двинин, 1952). В современный период летняя кета в сравнительно небольших количествах (в 2014 г. её уловы достигли 2432 т, что свидетельствует о сравнительно высокой численности, в предыдущий период уловы находились на уровне 1000−1600 т) фигурирует в статистике в качестве прилова при специализированном промысле горбуши.
Осенняя кета, воспроизводящаяся в реках зал. Терпения, относится к локальной группировке бассейна р. Поронай (Иванков, 1972, 1993; Каев, 2001). До 1951 г. численность осенней кеты естественного воспроизводства в р. Поронай находилась на сравнительно высоком уровне. В 1950−1960-х гг. её численность существенно снизилась, и осенняя кета практически утратила промысловое значение, что связано с воздействием молевого лесосплава и японского дрифтерного промысла (Гриценко и др., 1987). До начала XXI в. запасы осенней кеты на побережье зал. Терпения были невелики, её специализированный промысел в водах залива не вели. В небольшом количестве (до 50 т) её вылавливали в качестве прилова при промысле горбуши, в основном у рыбоводных заграждений лососёвых рыбоводных заводов (ЛРЗ) в притоках Пороная − Житница и Буюклинка. В 2003 г. отмечен существенный рост возврата рыб в эти реки. В настоящее время промысел кеты в водах зал. Терпения основан на осенней форме заводского воспроизводства (Каев, Игнатьев, 2006, 2015). Уловы за последние 10 лет (2004−2014 гг.) составляют в среднем 3.554 (максимальный 6.896) тыс. т, тогда как в 1946−1999 гг. − 0.780 тыс. т (рис. 4). Рост уловов связан как с усовершенствованием технологий искусственного воспроизводства, так и с изменением условий нагула лососей в Северной Пацифике (Кляшторин, Любушин, 2005; Котенев и др., 2014).
На побережье зал. Терпения (от м. Терпения до м. Тихий включительно) насчитывается 39 нерестовых рек, общая площадь нерестилищ в них составляет около 7.5 км2, что превосходит количество нерестилищ северо-восточного побережья Сахалина (6.02 км2) и является наибольшим по величине нерестовым фондом среди промысловых районов Сахалинской области. Наиболее значимые нерестовые водотоки района сосредоточены в бассейне р. Поронай − более 67% нерестовой площади бассейна зал. Терпения. Площадь нерестилищ в Поронае с учётом притоков (4.65 км2) почти в четыре раза превышает нерестовые площади горбуши в Тыми, второй по протяжённости реке Сахалина.
В бассейне Пороная воспроизводилось крупнейшее на Сахалине стадо горбуши, его численность в разные годы варьировала от сотен тысяч до нескольких миллионов особей. За годы наблюдений (1963−2014 гг.) в реки зал. Терпения заходило 0.21−16.69 (5.9) тыс. рыб. Самые малочисленные заходы отмечены в периоды падения запаса в конце 1960-х (1966, 1968), начале 1970-х (1970, 1972), в 1984 и 2000 гг.
Нерестилища горбуши начинаются в основном русле Пороная на расстоянии примерно 80 км от устья. Основные нерестовые площади расположены по руслу Пороная от устья р. Житница до верховьев (сплошные площади нерестилищ заканчиваются примерно в 200 км от устья), а также в крупнейших притоках (Житница, Орловка, Вальза, Матросовка и др.). До организации лесосплава (до 1950-х гг.) значительное количество горбуши заходило в реки Онорка и Северная Хандаса. В целом горбуша заходит или заходила практически во все реки бассейна Пороная, исключая реки тундрового типа.
Нерестилища рек зал. Терпения эксплуатируются горбушей неравномерно. По данным Гриценко с соавторами (1987), левобережные притоки верхнего течения бассейна Пороная наряду с реками юго-восточного побережья острова в прошлом веке принадлежали к наиболее интенсивно используемым на Сахалине. В таких водотоках, как Вальза, Житница, ключи Холодный и Берёзовый (левобережные притоки Пороная I порядка), в 1960−1970-х гг. при заходах высокочисленных поколений горбуши 1.5−2.0-кратное превышение нормативной плотности заполнения нерестилищ (220 экз/100 м2 (Рухлов, 1968)) было вполне обычным явлением. Сравнимая по интенсивности эксплуатация нерестилищ отмечена на Сахалине только на участке юго-восточного побережья от пос. Фирсово до протоки Красноармейская (оз. Тунайча) − в реках Фирсовка, Очепуха, Симау и Анна. По данным СахНИРО и Сахалинрыбвода, в 2004−2014 гг. средние показатели заполнения нерестилищ в левобережных притоках Пороная составили 68.7 (0.4−220.0) экз/100 м2, тогда как в правобережных притоках и основном русле Пороная соответственно 32.4 (0.2−200.0) и 35.0 (0.3−190.0) экз/100 м2. В период до 2010 г. на нерестилищах ручьёв Березовый и Холодный отмечали случаи переполнения нерестилищ, аналогичные таковым в 1960−1970-х гг. Однако площадь высокопродуктивных нерестилищ притоков левого берега Пороная по сравнению с таковой основного русла и более протяжённых правобережных притоков (Орловка, Онорка и др.) невелика, поэтому, несмотря на более высокие средние показатели заполнения левобережных нерестилищ (рис. 5), они вносят сравнительно небольшой вклад в общую продуктивность данной водной системы.
Рис. 5.
Заполнение производителями горбуши Oncorhynchus gorbusha нерестилищ основного русла (◼), правобережных (◻) и левобережных (◼) притоков верхнего течения р. Поронай в 2003−2014 гг.
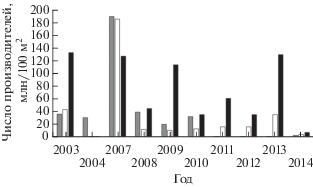
Низкий возврат горбуши в бассейн Пороная обусловлен высокой смертностью молоди при миграции по протяжённому руслу реки и значительным незаконным промыслом, развитым в этом районе. Даже при массовых заходах горбуши возврат производителей дочернего поколения сравнительно невелик. Так, в 2007 г. наблюдался массовый заход производителей горбуши на нерестилища Пороная и его притоков − 10.7 млн экз.; средние показатели заполнения нерестилищ достигли рекордных величин − 193 экз/100 м2. Возврат в цикличном 2009 г., несмотря на феноменально высокие показатели заполнения нерестилищ родительским поколением, находился на низком уровне и составил, по разным оценкам, 1.6−1.9 млн экз. при средних показателях заполнения 34.5 экз/100 м2 (рис. 6). Это заставляет отнести бассейн Пороная к районам неустойчивого воспроизводства горбуши. Кроме того, вызывают серьёзную тревогу экстремально низкие средние показатели заполнения нерестилищ Пороная в последние годы (рис. 7). По личному сообщению А.В. Захарова (СахНИРО), начиная с 2011 г. интенсивность эксплуатации нерестилищ существенно снизилась и отмечено прогрессирующее заиление нерестовых площадей. Эта ситуация свидетельствует о крайне неблагоприятных условиях воспроизводства горбуши в бассейне Пороная в настоящее время. Для восстановления численности стада горбуши р. Поронай и поддержания её на должном уровне необходимо существенно усилить контроль за незаконным промыслом, а также ввести ограничение либо временный запрет промышленного лова данного вида в зал. Терпения (как минимум, в устье Пороная).
Рис. 6.
Динамика численности скатывающейся из рек молоди (◼) и заходящих на нерест производителей (◻) горбуши Oncorhynchus gorbusha в бассейне р. Поронай в 2003−2014 гг.
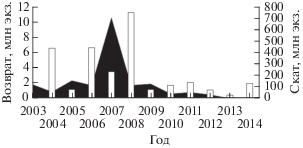
Рис. 7.
Заполнение производителями горбуши Oncorhynchus gorbusha нерестилищ бассейна Пороная (◻) и рек западного побережья зал. Терпения (◼) в 2011−2014 гг.
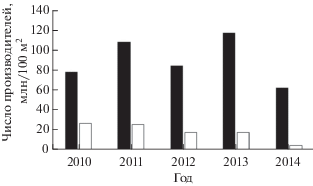
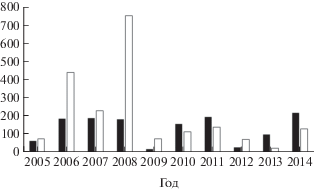
Реки западного побережья залива, впадающие севернее м. Тихий (Макаровский район), более короткие, выраженного горно-предгорного типа; по гидрогеологическим характеристикам они несопоставимы с Поронаем (протяжённой рекой, имеющей на большей части русла выраженный равнинный тип), как несопоставимы и уловы горбуши по отношению к площади нерестилищ. Несопоставим и нерестовый фонд. В современный период средняя численность производителей горбуши на нерестилищах рек западного побережья выше, чем на нерестилищах Пороная (рис. 7), что свидетельствует о более благоприятной ситуации с воспроизводством данного вида. В последние годы выше и численность мигрирующей молоди горбуши (рис. 8), несмотря на значительно меньшую площадь нерестового фонда.
Рис. 8.
Численность молоди горбуши Oncorhynchus gorbusha, мигрировавшей из бассейна Пороная и рек западного побережья зал. Терпения в 2005−2014 гг., млн экз.; обозначения см. на рис. 7.
Летняя кета, в отличие от осенней формы, откладывает икру на нерестилища, омываемые подрусловым потоком, т.е. практически на тех же участках русла, где нерестится горбуша (Лапшина и др., 2012). В прошлом веке наиболее обширные нерестилища летней кеты отмечали в р. Лонгари; в основном русле Пороная они простирались от устья р. Вальза до истоков Пороная, некоторое количество нерестилищ находилось в притоках Вальза, Житница, Онорка, Северная Хандаса и Сергеевка, в меньшей степени – в реках Южная Хандаса, Седьмая, Девятая и Десятая речки (неопубликованные данные Главгосрыбвода). В настоящее время численность летней формы низка, её мониторинг на нерестилищах не ведут, современная площадь и локализация нерестилищ неизвестны.
Нерестилища осенней формы кеты в бассейне Пороная приурочены к местам выхода грунтовых вод в основном русле Пороная, на участке от р. Чёрная до устья р. Вальза, и в притоках нижнего и среднего течения (Житница, Буюклинка, Леонидовка, Орловка, Ельная, Матросовка и др.). Из притоков верхнего течения осеннюю кету отмечали в р. Вальза, ключах и ручьях Рыбоводный, Костин, Каменка, Вторая речка и Берёзовый. Численность производителей на естественных нерестилищах в современный период чрезвычайно низкая, точные площади функционирующих нерестилищ не известны.
Основной запас осенней кеты зал. Терпения формируется на двух крупных ЛРЗ − Буюкловском и Побединском, расположенных в бассейне Пороная. Летнюю кету в небольших объёмах разводят на Побединском ЛРЗ (Лапшина и др., 2012). В 2004−2014 гг. суммарный выпуск молоди кеты с этих ЛРЗ варьировал в пределах 35.9−60.3 (47.65) млн экз. На западном побережье залива функционируют четыре ЛРЗ – на реках Лазовая, Нитуй, Пугачевка, Тихая; из них один (на р. Лазовая) выпускает кету, а на трёх разводят преимущественно горбушу; выпуск её молоди в 2004−2014 гг. составлял 11.8−42.6 (28.83) млн экз.
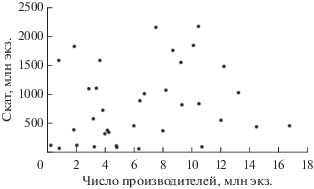
Анализ зависимости родители−потомки показал, что максимальный урожай молоди приходится преимущественно на диапазон численности родительского стада 3.63−10.42 млн экз.; при заходе на нерест более 10.42 млн производителей величина пополнения снижается (рис. 9).
Рис. 9.
Соотношение между величиной пропуска производителей горбуши Oncorhynchus gorbusсha на нерестилища в 1978−2014 гг. и последующим скатом молоди в реках зал. Терпения.
Симу и кижуча зал. Терпения в основном используют в качестве прилова либо как объекты любительского и спортивного рыболовства. Вылов симы невелик и в настоящее время варьирует от 0.3 до 1.7 т. Нерестилища симы расположены в тех же реках, где и нерестилища горбуши, но значительно выше их по течению.
Основные нерестилища кижуча расположены в верхнем течении р. Поронай (выше 270 км от устья) и в среднем течении р. Вальзы, в меньшем количестве он заходит на нерест в притоки нижнего и среднего течения Пороная (Каменка, Буюклинка, Житница, Матросова Верхняя и др.) (Гриценко и др., 1987). Вылов кижуча за последние пять лет варьируется от 358 до 808 т.
ЗАКЛЮЧЕНИЕ
Воспроизводство горбуши − наиболее многочисленного вида тихоокеанских лососей в зал. Терпения − в настоящее время сосредоточено в сравнительно небольших горно-предгорных реках западного побережья (Макаровский городской округ). Основной водоток северного побережья зал. Терпения – р. Поронай − практически потерял значимость для воспроизводства данного вида. Даже в наиболее продуктивных в недавнем прошлом левобережных притоках Пороная (ключи Холодный, Березовый), имеющих выраженные горно-предгорные характеристики и обладающих высоким потенциалом воспроизводства горбуши, её производители на протяжении последних лет встречаются единично. Сравнительно эффективное воспроизводство горбуши в отдельные годы может происходить в реках бассейна оз. Невское. Осенняя форма кеты, второго по численности вида тихоокеанских лососей в рассматриваемом районе, может эффективно размножаться только в сравнительно крупных реках на участках русла, имеющих выраженный равнинный тип. Естественное воспроизводство осенней кеты в бассейне Пороная отмечается только в отдельные годы в незначительных количествах; запас формируется за счёт разведения на лососёвых рыборазводных заводах. Вместе с тем в бассейне Пороная наблюдается некоторое увеличение численности редкой летней формы кеты, что даёт надежду на её сохранение. По результатам анализа многолетних данных, оптимальная величина пропуска производителей горбуши для заполнения нерестилищ зал. Терпения составляет 3.63−10.42 млн экз.
БЛАГОДАРНОСТИ
Авторы выражают искреннюю благодарность всем коллегам, совместно с которыми выполняли исследования на реках бассейна зал. Терпения. Особая признательность А.В. Захарову (СахНИРО) за многолетнее сотрудничество в полевых работах на реках бассейна р. Поронай, оз. Невское и реках Макаровского городского округа. Отдельное спасибо А.М. Каеву (СахНИРО) за конструктивную критику и замечания.
Список литературы
Антонов А.А. 2005. Биология и динамика численности горбуши зал. Анива (о. Сахалин): Автореф. дис. … канд. биол. наук. Владивосток: ТИНРО-центр, 24 с.
Атлас Сахалинской области. 1967. М.: Изд-во ГУГК при Совмине СССР, 135 с.
Воловик С.П. 1967. Структура нерестовых стад и эффективность естественного воспроизводства горбуши на южном Сахалине: Автореф. дис. … канд. биол. наук. Калининград: КТИРПХ, 25 с.
Воловик С.П., Ландышевская А.Е., Смирнов А.И. 1972. Материалы по эффективности размножения горбуши на южном Сахалине // Изв. ТИНРО. Т. 81. С. 69−90.
Голованов И.С. 1982 О естественном воспроизводстве горбуши Oncorhynchus gorbusha (Walbaum) на северном побережье Охотского моря // Вопр. ихтиологии. Т. 22. Вып. 4. С. 568−575.
Гриценко О.Ф. 2002. Проходные рыбы острова Сахалин (систематика, экология, промысел). М.: Изд-во ВНИРО, 248 с.
Гриценко О.Ф. Ковтун А.А., Косткин В.К. 1987. Экология и воспроизводство кеты и горбуши. М.: Агропромиздат, 166 с.
Гриценко О.Ф., Гришин А.Ф., Захаров А.В., Шелепаха Г.Н. 1989. Воспроизводство горбуши Oncorhynchus gorbuscha (Walbaum) о-ва Сахалин // Вопр. ихтиологии. Т. 29. Вып. 3. С. 377−386.
Двинин П.А. 1952. Лососи южного Сахалина // Изв. ТИНРО. Т. 37. С. 69−108.
Ефанов В.Н. 2003. Организация мониторинга и моделирование запасов популяций рыб (на примере горбуши Oncorhynchus gorbuscha Walb.). Южно-Сахалинск: Изд-во СахГУ, 134 с.
Живоглядов А.А. 2014. Рыбы малых и средних рек острова Сахалин: пространственное распределение, структура и динамика // Вопр. ихтиологии. Т. 54. № 1. С. 1–11.
Живоглядов А.А., Никитин В.Д., Промашкова О.А., Прохоров А.П. 2011. Некоторые подходы к изучению видовой и пространственной структуры сообществ рыб равнинной части русла р. Поронай // Тр. СахНИРО. Т. 12. С. 55−71.
Живоглядова Л.А., Даирова Д.С., Лабай В.С. 2012. Состав, структура и сезонная динамика макрозообентоса рек восточного Сахалина // Изв. ТИНРО. Т. 171. С. 199−209.
Живоглядов А.А., Антонов А.А., Руднев В.А., Ким Хе Юн. 2013. О вариациях выживаемости эмбрионально-личиночных стадий горбуши Oncorhynchus gorbuscha и кеты Oncorhynchus keta на нерестилищах рек о. Сахалин // Вопр. рыболовства. Т. 14. № 2. С. 242–258.
Заварина Л.О. 2008. Биология и динамика численности кеты (Oncorhynchus кета) северо-восточного побережья Камчатки: Автореф. дис. … канд. биол. наук. Петропавловск-Камчатский: КамчатНИРО, 25 с.
Золотухин С.Ф. 2003. Нерестовый фонд и современный статус популяций лососей в Приморском крае: Автореф. дис. … канд. биол. наук. Владивосток: ТИНРО-центр, 24 с.
Иванков В.Н. 1972. Особенности экологии и структура популяций осенней кеты различных районов Сахалина // Уч. зап. ДВГУ. № 60. С. 27–35.
Иванков В.Н. 1993. Популяционная организация у тихоокеанских лососей с коротким пресноводным периодом жизни // Вопр. ихтиологии. Т. 33. № 1. С. 78–83.
Иванков В.Н., Андреев В.Л. 1972. Экология и моделирование популяций горбуши южных Курильских островов // Уч. зап. ДВГУ. Т. 60. С. 3−26.
Инструкция о порядке проведения обязательных наблюдений за дальневосточными лососевыми на КНС и КНП бассейновых управлений рыбоохраны и стационарах ТИНРО. 1987. Владивосток: Изд-во ТИНРО, 23 с.
Каев A.M. 2001. Распространение осенней кеты в связи с особенностями водоносных комплексов Сахалина и Курильских островов // Чтения памяти B.Я. Леванидова. Вып. 1. С. 344–349.
Каев А.М. 2007. Биологические основы рационального промысла лососей в сахалино–курильском регионе // Вопр. рыболовства. Т. 8. № 4 (32). С. 713–733.
Каев А.М. 2008. Результаты мониторинга нерестовых подходов горбуши в Сахалинской области в 2008 г. // Бюл. № 3 реализации «Концепции дальневосточной бассейновой программы изучения тихоокеанских лососей». С. 24–29.
Каев А.М. 2009. Мониторинг состояния запасов горбуши в основных районах ее промысла в Сахалинской области в 2009 г. // Бюл. № 4 реализации “Концепции дальневосточной бассейновой программы изучения тихоокеанских лососей”. С. 28–33.
Каев А.М. 2011а. Оценка эффективности прогнозирования и управления промыслом горбуши в Сахалино-Курильском регионе // Изв. ТИНРО. Т. 167. С. 32–53.
Каев А.М. 2011б. Методическое руководство по количественному учету покатной молоди горбуши и кеты в малых реках методом выборочных обловов. Южно-Сахалинск: Изд-во СахНИРО, 16 с.
Каев А.М., Игнатьев Ю.И. 2006. Заводское разведение лососей в Сахалинской области // Рыб. хоз-во. № 6. С. 57–60.
Каев А.М., Игнатьев Ю.И. 2015. Развитие заводского разведения тихоокеанских лососей в Сахалино-Курильском регионе и его значение для промысла // Тр. ВНИРО. Т. 153. С. 95–104.
Каев А.М., Авдеев Д.В., Антонов А.А. и др. 2010. Результаты мониторинга подходов горбуши в основных районах ее промысла в Сахалинской области в 2010 г. // Бюл. № 5 реализации “Концепции дальневосточной бассейновой программы изучения тихоокеанских лососей”. С. 30–35.
Казаков Н.А., Генсиоровский Ю.В. 2007. Влияние вертикального градиента осадков на характеристики гидрологических, лавинных и селевых процессов в низкогорье // Геоэкология. Инженер. геология. Гидрогеология. Геокриология. № 4. С. 342–347.
Ким Х.Ю., Антонов А.А. 2002. Уровень заполнения рек производителями как один из факторов становления численности горбуши в заливе Анива // Сб. науч. тр. молодых ученых МГТА. Вып. II. С. 52−57.
Кляшторин Л.Б., Любушин А.А. 2005. Циклические изменения климата и рыбопродуктивности. М.: Изд-во ВНИРО, 235 с.
Котенев Б.Н., Кровнин А.С., Кловач Н.В. и др. 2014. Влияние климато-океанологических факторов на состояние основных запасов горбуши в 1950–2015 гг. // Тр. ВНИРО. Т. 158. С. 114−161.
Лабай В.С., Живоглядова Л.А., Полтева А.В. и др. 2015. Водотоки острова Сахалин: жизнь в текучей воде. Южно-Сахалинск: Изд-во Сахалин. обл. краевед. музея, 236 с.
Лакин Г.Ф. 1980. Биометрия. М.: Высш. шк., 293 с.
Лапшина А.Е., Игнатьев Ю.И., Кузнецова Л.Д., Латушкина Е.В. 2012. Опыт искусственного воспроизводства летней кеты в Сахалинской области и Хабаровском крае // Науч. тр. Дальрыбвтуза. № 27. С. 32−44.
Леванидов В.Я. 1964. О связи между плотностью заполнения нерестилищ и эффективностью нереста амурских лососей // Изв. ТИНРО. Т. 55. С. 65−74.
Линдберг Г.У. 1953. Закономерности распределения рыб и геологическая история дальневосточных морей // Очерки по общим вопросам ихтиологии. М.; Л.: Изд-во АН СССР. С. 47–57.
Макеев С.С. 2010. Регулирование заполнения нерестовых рек производителями лососей. Южно-Сахалинск: Сахалинрыбвод, 52 с. (http://smak02.ru/images/files/nauka)
Методические рекомендации по учету численности тихоокеанских лососей в реках Сахалинской области. 2013. Южно-Сахалинск: Изд-во СахНИРО, 31 с.
Никифоров С.Н. 2001. Ихтиофауна пресных вод Сахалина и ее формирование: Автореф. дис. … канд. биол. наук. Южно-Сахалинск: СахГУ, 25 с.
Никифоров С.Н., Гришин А.Ф. 1989. Распределение рыб в пресноводных водоемах Сахалина и возможный генезис ихтиофауны в северо-западной части острова // Вопр. ихтиологии. Т. 20. Вып. 5. С. 746–753.
Никифоров С.Н., Гришин А.Ф., Захаров А.В., Шелепаха Г.Н. 1997. Состав ихтиофауны и распределение рыб в бассейнах рек Поронай и Тымь (Сахалин) // Там же. Т. 37. № 3. С. 329–337.
Плохинский Н.А. 1961. Биометрия. Новосибирск: Изд-во АН СССР, 362 с.
Правдин И.Ф. 1966. Руководство по изучению рыб. М.: Пищ. пром-сть, 370 с.
Ресурсы поверхностных вод СССР. 1973. Т. 18. Дальний Восток. Вып. 4. Сахалин и Курилы. Л.: Гидрометеоиздат, 264 с.
Риккер В.Е. 1971. Сопоставление двух кривых воспроизводства // Рыб. хоз-во. № 3. С. 16–21.
Рухлов Ф.Н. 1968. Речной период жизни сахалинской горбуши // Там же. № 12. С. 15−17.
Рухлов Ф.Н. 1982. Жизнь тихоокеанских лососей. Южно-Сахалинск: Дальневост. книж. изд-во, 110 с.
Рухлов Ф.Н. 2007. Хроники сахалинского рыболовства. Южно-Сахалинск: Изд-во СахНИРО, 144 с.
Сафронов С.Н. 2000. Экологические группы и пространственное распределение рыб малых рек острова Сахалин // Чтения памяти проф. В.В. Станчинского. Вып. 3. С. 59–63.
Сафронов С.Н., Никифоров С.Н. 1995. Видовой состав и распространение ихтиофауны пресных и солоноватых вод Сахалина // Матер. ХХХ науч.-метод. конф. преподавателей ЮСГПИ. Южно-Сахалинск: Изд-во ЮСГПИ. С. 112–124.
Сафронов С.Н., Никифоров С.Н. 2003. Список рыбообразных и рыб пресных и солоноватых вод Сахалина // Вопр. ихтиологии. Т. 43. № 1. С. 42–53.
Сафронов С.Н., Литенко Н.Л., Пешеходько В.М. и др. 2000. Эколого-биоценотическая характеристика и качество вод внутренних водоемов острова Сахалин // Чтения памяти профессора В.В. Станчинского. Вып. 3. С. 321−329.
Сафронов С.Н., Никитин В.Д., Заварзина Н.К. и др. 2008. Видовая структура, численность и промысел рыб озера Невское (о. Сахалин) // Тр. СахНИРО. Т. 10. С. 208–235.
Семко Р.С. 1939. Камчатская горбуша // Изв. ТИНРО. Т. 16. 111 с.
Таранец А.Я. 1937а. Краткий определитель рыб советского Дальнего Востока и прилежащих вод // Там же. Т. 11. С. 1–200.
Таранец А.Я. 1937б. Материалы к познанию ихтиофауны советского Сахалина // Там же. Т. 12. С. 5–44.
Таранец А.Я. 1939. Исследования нерестилищ кеты и горбуши в р. Иски // Рыб. хоз-во. № 12. С. 1−4.
Фельдман М.Г., Шевляков Е.А., Артюхина Н.Б. 2016. Оценка ориентиров пропуска производителей тихоокеанских лососей Oncorhynchus в бассейнах рек восточной и юго-восточной Камчатки // Исследования водных биологических ресурсов Камчатки и северо-западной части Тихого океана. Вып. 41. С. 51–80.
Шмидт П.Ю. 1905. Рыбные промыслы Дальнего Востока. Морские промыслы острова Сахалин. СПб.: Тип. Г.П. Пожарова, 458 с.
Шунтов В.П. 2010. О программах комплексных исследований тихоокеанских лососей на период 2010−2014 гг. // Бюл. № 4 реализации “Концепции дальневосточной бассейновой программы изучения тихоокеанских лососей”. С. 183−199.
Шунтов В.П. 2016. Концептуальные заметки об управлении биологическими ресурсами, рациональном и устойчивом рыболовстве // Вопр. рыболовства. Т. 17. № 1. С. 5–19.
Wickett W.P. 1958. Review of certain environmental factors affecting the production of pink and chum salmon // J. Fish. Res. Board Can. V. 15. № 5. P. 1103−1126.
Дополнительные материалы отсутствуют.
Инструменты
Вопросы ихтиологии