

Вопросы ихтиологии, 2019, T. 59, № 4, стр. 375-404
Материалы к ревизии рода Photonectes (Melanostomiidae)
1 Институт проблем экологии и эволюции им. А.Н. Северцова РАН – ИПЭЭ
Москва, Россия
2 Институт океанологии им. П.П. Ширшова РАН – ИО РАН
Москва, Россия
* E-mail: prokartster@gmail.com
Поступила в редакцию 06.10.2018
После доработки 07.02.2019
Принята к публикации 07.02.2019
Аннотация
Критически проанализированы признаки внешней морфологии меланостомовых рыб рода Photonectes. Показано, что признаки, ранее применявшиеся для разграничения подродов, непригодны для этих целей. Подтверждено существование в составе рода только двух чётко диагностируемых подродов, Trachinostomias и Photonectes s. str., для которых даны ревизованные диагнозы на основании нового набора признаков. Вид P. uncinatus перемещен из подрода Trachinostomias в номинативный подрод. Кратко обсуждены основания для выделения групп видов в составе номинативного подрода и филогенетические отношения между этими группами. Описано три новых вида: P. distichodon, P. filipendulus и P. xenopogon spp. n. Указаны отличительные признаки ещё одного нового вида, из-за недостатка данных остающегося пока не описанным. Дана таблица для определения всех известных видов рода.
Род Photonectes Günther, 1887 является наименее разработанной в таксономическом отношении группой меланостомовых рыб, видовое разнообразие которой в настоящее время остаётся в значительной мере недооценённым, а принципы видовой диагностики и надвидовой классификации – недостаточно разработанными. Между тем наряду с Bathophilus Giglioli, 1882 это, вероятно, одна из наиболее диверсифицированных групп меланостомовых рыб, по-видимому, уступающая по числу видов лишь гипердиверсифицированному роду Eustomias Vaillant, 1888. С учётом настоящей публикации в составе рода содержится 34 номинальных вида, из которых 24 или 25 являются валидными (табл. 1). Кроме них мне известны ещё по меньшей мере четыре неописанных вида.
Таблица 1.
Список номинальных и валидных видов рода Photonectes
| Номинальный вид | Валидное название | Примечание |
|---|---|---|
| Подрод Trachinostomias | ||
| Eustomiasaguayoi Barbour, 1942 | P. margarita | |
| P.bifilifer Beebe, 1933 | P. parvimanus | |
| P. filipendulus sp. nova | P. filipendulus | |
| P.fimbria Regan et Trewavas, 1930 | P. parvimanus | |
| P.flagellatus Parr, 1927 | P. margarita | |
| P. (Trachinostomias) gorodinskii | P. gorodinskii | Отличия от P. parvimanus нуждаются в уточнении на более представительном материале |
| P.intermedius Parr, 1927 | P. margarita | Под названием “intermedius” описана молодь P. margarita |
| Echiostoma margarita Goode et Bean, 1896 | P. margarita | Возможно, сборный вид в настоящем понимании |
| P.monodactylus Regan et Trewavas, 1930 | P. margarita | |
| P. munificus Gibbs, 1968 | P. munificus | Родственные отношения с другими видами рода нуждаются в уточнении |
| P. parvimanus Regan et Trewavas, 1930 | P. parvimanus | |
| Echiostomarichardi Zugmayer, 1913 | P. margarita | |
| P. xenopogon sp. nova | P. xenopogon | |
| Подрод Photonectes s. str. | ||
| P. achirus Regan et Trewavas, 1930 | P. achirus | Группа “caerulescens”; наибольший известный экземпляр имеет SL 83.5 мм (Flynn, Klepadlo, 2012) |
| Lucifer albipennis Döderlein, 1882 | P. albipennis | Группа “albipennis”; возможно, сборный вид в настоящем понимании |
| P. barnetti Klepadlo, 2011 | P. barnetti | Группа “albipennis”; отличия от P. coffea нуждаются в уточнении |
| Melanostomias braueri Zugmayer, 1913 | P. braueri | Группа “braueri” |
| P. caerulescens Regan et Trewavas, 1930 | P. caerulescens | Группа “caerulescens” |
| P. coffea Klepadlo, 2011 | P. coffea | Группа “albipennis” |
| P. cornutus Beebe, 1933 | ? | Species inquirenda; синонимия с P. mirabilis недостаточно обоснована |
| P. corynodes Klepadlo, 2011 | P. corynodes | Известен только по ювенильному голотипу; близок к P. litvinovi |
| P. dinema Regan et Trewavas, 1930 | P. dinema | Группа “braueri”; известен только по молоди |
| P. distichodon sp. nova | P. distichodon | Группа “braueri” |
| P. gracilis Goode et Bean, 1896 | P. gracilis | Группа “caerulescens” |
| P. leucospilus Regan et Trewavas, 1930 | P. leucospilus | Группа “braueri”; известен только по молоди |
| P. litvinovi Prokofiev, 2014 | P. litvinovi | Близок к P. corynodes |
| P. (Dolichostomias) mirabilis Parr, 1927 | P. mirabilis | Группа “caerulescens” |
| P.ovibarba Regan et Trewavas, 1930 | P. braueri | Под названием “ovibarba” описана молодь P. braueri |
| P. paxtoni Flynn et Klepadlo, 2012 | P. paxtoni | Родственные отношения с другими видами Photonectes s. str. нуждаются в уточнении |
| P.paucidentatus Koefoed, 1956 | P. braueri | |
| P. phyllopogon Regan et Trewavas, 1930 | P. phyllopogon | Известен только по молоди |
| P. (Trachinostomias) uncinatus Prokofiev, 2015 | P. uncinatus | Группа “braueri” |
| P. venetaenia Prokofiev, 2016 | P. venetaenia | Группа “caerulescens” |
| Photonectes waitti Flynn et Klepadlo, 2012 | P. waitti | Группа “albipennis”; известен только по молоди, возможно, является ювенильной стадией вида P. coffea |
Внешне виды рода Photonectes отличаются от других меланостомовых рыб длинной изогнутой и выступающей вперед нижней челюстью, симфиз которой помещается в широкий промежуток между симфизными концами praemaxillaria. Подобное строение нижней челюсти отмечено еще только у рода Tactostoma Bolin, 1939, отличающегося от Photonectes намного более удлинённым телом, многорядными зубами в челюстях и рядом остеологических особенностей, свидетельствующих против их близкого родства (Fink, 1985). В качестве апоморфных признаков Photonectes указывалось наличие удлинённого заднего отростка anguloarticulare, удлинённое hypohyale и наличие плотного соединительнотканного хрящевого образования между коракоскапулярной пластинкой и cleithrum (Fink, 1985). Нужно, однако, отметить, что Финк (Fink, 1985) изучил только немногие виды рода и с разной степенью детализации, более того, типовой вид рода (P. albipennis) остался им неисследованным, в связи с чем монофилию рода нельзя считать доказанной. Сам Финк (Fink, 1985) отмечал, что существенные остеологические различия были выявлены им даже между экземплярами, считающимися конспецифичными (что не удивляет, так как, судя по списку материала, речь идет о P. margarita, под именем которого ранее смешивалось не менее четырёх разных видов). Несомненно, что полноценный морфологический анализ всех представителей рода приведёт к значительным изменениям существующих представлений о морфологической изменчивости и классификации данного рода.
В настоящее время проведение такого анализа затруднительно, так как ряд видов исключительно редки в коллекциях и рассеяны по многим музеям мира, а некоторые из них известны только по голотипам; ряд видов известен только по ювенильным стадиям; кроме того, вследствие особенностей строения кожи и подкожного соединительнотканного слоя многие виды сильно повреждаются в тралах, из-за чего оценка некоторых таксономически и филогенетически значимых признаков может быть произведена у них лишь в исключительных случаях. Вместе с тем морфологическая неоднородность фотонектов, даже только на основании признаков внешнего строения, была отмечена давно, что привело к выделению в составе этого рода до пяти подродов. Первые ревизии рода провели Парр (Parr, 1927) и Ригэн и Тревавс (Regan, Trewavas, 1930), выделившие подроды Dolichostomias Parr, 1927; Melanonectes Regan et Trewavas, 1930; Microchirichthys Regan et Trewavas, 1930; Photonectes s. str. и Trachinostomias Parr, 1927 на основании таких признаков, как наличие или отсутствие лучей грудного плавника и их число, наличие или отсутствие кожи на вертикальных плавниках и положение брюшных плавников по отношению к вершине рыла и основанию хвостового плавника. Позже было предложено синонимизировать подрод Microchirichthys с Trachinostomias (Morrow, Gibbs, 1964), и в таком виде подродовая классификация фотонектов просуществовала до самого последнего времени (Klepadlo, 2011). В указанной работе (Klepadlo, 2011) в качестве дополнительного признака для различения подродов была предложена величина промежутка между вершиной истмуса и первой парой фотофоров IP, но число и состав подродов остались неизменными. Однако позже мною было показано, что большинство признаков, положенных в основу подродового деления, являются либо плезиоморфными, либо варьирующими, иногда на индивидуальном уровне, что делает затруднительным разграничение подродов и не даёт оснований для признания их монофилетичными (Прокофьев, 2015). Кроме того, прежняя классификация не учитывала некоторых, возможно, филогенетически значимых признаков (например, наличие голубой светящейся ткани на теле, по которому Dolichostomias должен быть объединён с частью видов Photonectes s. str.). Поэтому было предложено сохранить в составе рода лишь два подрода (Trachinostomias и Photonectes s. str.), различая их на основании наличия или отсутствия кожи на вертикальных плавниках и промежутка между семью–восемью передними и двумя–четырьмя задними фотофорами IP, а подроды Melanonectes и Dolichostomias синонимизировать с номинотипическим подродом (Прокофьев, 2015).
Однако последующая работа с материалом и обнаружение новых экземпляров показали, что реальная ситуация не столь проста, как представлялось. Более того, оказалось, что такой “классический” признак, как захождение кожи на лучи вертикальных плавников, оказывается очень изменчивым в онтогенезе некоторых видов, в связи с чем одни экземпляры одного и того же вида (например, P. caerulescens) на этом основании могут быть отнесены к Trachinostomias, а другие – к Photonectes s. str. Поэтому я предпринял попытку критически пересмотреть признаки, использовавшиеся ранее в подродовой классификации фотонектов, и попытаться найди дополнительные критерии для характеристики совокупностей видов внутри этого рода. Результаты этого исследования изложены в настоящем сообщении. В связи с редкостью многих видов в коллекциях работа была ограничена лишь признаками внешней морфологии, поэтому она не претендует на завершённость. Помимо этого для демонстрации морфологического разнообразия критически значимых в систематике фотонектов структур потребовалось описание трёх новых видов, также включённое в статью. Еще один, по-видимому, новый вид оставлен пока в открытой номенклатуре из-за недостаточной для формального описания сохранности единственного ювенильного образца.
МАТЕРИАЛ И МЕТОДИКА
Материал, на основании которого выделяются новые виды, приведён при описании последних. Для сравнения и анализа признаков были изучены следующие экземпляры 18 видов Photonectes.
В работе использованы сокращения признаков: SL – стандартная длина; D, A, P, V – соответственно спинной, анальный, грудные и брюшные плавники и число лучей в них; vert. – общее число позвонков (включая уростилярный); сокращённые названия учреждений: ИО РАН – Институт океанологии РАН, г. Москва; ЗММУ – Зоологический музей Московского государственного университета; BMNH – Музей естественной истории, г. Лондон (Великобритания); MNHN – Национальный музей естественной истории, г. Париж (Франция); ZMUC – Зоологический музей Университета Копенгагена, Дания; другие сокращения: БМРТ – большой морозильный рыболовный траулер, БНТ – большой нейстонный трал, НИС – научно-исследовательское судно, НПС – научно-промысловое судно, ПС – планктонная сеть, РС – рыболовное судно, РТ – рыболовный траулер, РТАК – разноглубинный трал Айзекса–Кидда, РТАКСА – РТАК в модификации Самышева–Асеева, ст. – океанографическая станция. Коллекторы указаны так, как на этикетке. Использована стандартная номенклатура фотофоров стомиеобразных рыб (Morrow, Gibbs, 1964).
P. albipennis (44 экз.): ИО РАН – 2 экз. SL 170 и 193 мм, без этикетки; ИО РАН – 6 экз. SL 40–64 и 137 мм, 205–“3”, проба 371 (район Куросио?); ИО РАН – 1 экз. SL 242 мм, район Куросио, банка № 13, коллектор Соколовский (ТИНРО); ИО РАН – 1 экз. SL 160 мм, Восточно-Китайское море, 28°30ʹ с.ш., 130°00′ в.д., 100–0 м, глубина 840 м; ИО РАН – 6 экз. SL 143–219 мм, НПС “Фиолент”, рейс 4(9), трал 55, 04.09.1977 г.; ИО РАН – 1 экз. SL 245 мм, Индийский океан, 24°04ʹ ю.ш., 112°27ʹ в.д., пелагический трал 220–520 м, БМРТ “Таджикистан”, время 24.00, коллекторы Макаров, Гусев; ИО РАН – 2 экз. SL 185 и 188 мм, 09°07ʹ ю.ш., 131°02ʹ в.д., 450 м, БМРТ “Академик Берг”, 20.07.1967 г.; ИО РАН – 1 экз. SL 190 мм, 8°30ʹ ю.ш., 110°56ʹ в.д., о. Ява, 450–400 м, НПС “Профессор Месяцев”, рейс 7, трал 47, 27.04.1979 г.; ИО РАН – 4 экз. SL 68–170 мм, 31°02ʹ с.ш., 150°31ʹ в.д., НИС “Экватор”, ст. 251, 14.11.1968 г., разноглубинный трал 31 м, длина ваеров 350 м, скорость 3.8 узла, коллектор Комраков; ИО РАН – 1 экз. SL 156 мм, 32°36ʹ с.ш., 149°18ʹ в.д., 80–100 м, НИС “Экватор”, ст. 255, 15.12.1968 г.; ИО РАН – 2 экз. SL 203 и 251 мм, северная часть Тихого океана (данные о координатах лова не сохранились), НИС “Экватор”, трал 260, 22.01.1973 г.; ИО РАН – 1 экз. SL 180 мм, 01°08ʹ с.ш., 56°32ʹ в.д., НИС “Рифт”, рейс 2, ст. 26, проба 39, донный оттер-трал; ИО РАН – 1 экз. SL 209 мм, Индийский океан, возвышенность Боа-Паш, НПС “Фиолент”, рейс 3, трал 177; ИО РАН – 1 экз. SL 32.5 мм, 23°57ʹ9ʹʹ ю.ш., 112°14ʹ2ʹʹ в.д., НИС “Витязь”, рейс 31, ст. 4564, проба 212, сачок, слой облова – поверхность; ИО РАН – 1 экз. SL 36 мм, 12°21ʹ3ʹʹ ю.ш., 103°35ʹ1ʹʹ в.д., НИС “Витязь”, рейс 33, ст. 5005, проба 881, ПС, слой облова – поверхность; ИО РАН – 1 экз. SL 23.5 мм, 11°05ʹ5ʹʹ ю.ш., 140°04ʹ8ʹʹ з.д., НИС “Витязь”, рейс 34, ст. 5103, проба 193, ПС, слой облова – поверхность, на свет, время 21.30–21.50, 03.10.1961 г.; ИО РАН – 1 экз. SL 32 мм, 07°20ʹ ю.ш., 124°22ʹ в.д., НИС “Витязь”, рейс 57, ст. 7259, РТАК 62, проба 130; ИО РАН – 1 экз. SL 187 мм, 01°53ʹ ю.ш., 128°30ʹ в.д., НИС “Витязь”, рейс 57, ст. 7277, РТАК 73; ИО РАН – 2 экз. SL 105 и 150 мм, 26°23ʹ с.ш., 143°19ʹ в.д., НИС “Витязь”, рейс 57, ст. 7395, РТАК 135; ИО РАН – 1 экз. SL 170 мм, 29°09ʹ с.ш., 142°38ʹ в.д., НИС “Витязь”, рейс 57, ст. 7400, РТАК 140; ИО РАН № 780 – 2 экз. SL 158 и 163 мм, 25°29ʹ–25°35ʹ ю.ш., 35°09ʹ–35°01ʹ в.д., НИС “Витязь-II”, рейс 17, ст. 2631, 23.11.1988 г.; ЗММУ № 14590 – 5 экз. SL 161–230 мм, 08°29ʹ ю.ш., 59°35ʹ в.д., НПС “Звезда Крыма”, рейс 6, трал 3, 19.03.1976 г.
P. barnetti (3 экз.): ИО РАН – 1 экз. SL 112 мм, район Гаваев, банка Милуоки, горизонт лова 200–0 м, БМРТ “Академик Берг”, 09.05.1970 г.; ИО РАН – 1 экз. SL 160 мм, 03°02ʹ с.ш., 130°57ʹ в.д., НИС “Витязь”, рейс 57, ст. 7284, РТАК 78; ИО РАН – 1 экз. SL 124 мм, 01°37ʹ ю.ш., 143°57ʹ в.д., НИС “Дмитрий Менделеев”, рейс 18, ст. 1531, РТАК 22, трос 3800 м, проба 51, 09.02.1977 г.
P. braueri (6 экз.): MNHN № 1938–0133 – 1 экз. SL 26 мм, синтип P. ovibarba, 24°00ʹ с.ш., 76°00ʹ з.д.; ИО РАН – 1 экз. SL 22 мм, 07°45ʹ с. ш., 120°25ʹ в.д., горизонт лова 1500–0 м, глубина 4062 м, НИС “Витязь”, рейс 33, ст. 5028, проба 913, ринг-трал, время 22.00–22.50, 04–05.04.1961 г.; ИО РАН – 1 экз. SL 50 мм, 37°51ʹ с.ш., 43°55ʹ з.д., горизонт лова 550 м (вытравлено 1500 м троса), НИС “Петр Лебедев”, IV атлантическая экспедиция, проба 123А (IV–123A), РТАК, время 00.40–01.10; ИО РАН – 1 экз. SL 50 мм, 26°20ʹ6ʹʹ ю.ш., 90°02ʹ1ʹʹ в.д., НИС “Витязь”, рейс 31, ст. 4577, перлоновый ринг-трал 160 см, проба 248, 17–18.12.1959 г.; ИО РАН – 1 экз. SL 43 мм, 26°07ʹ ю.ш., 26°50ʹ з.д., 1500 и 1900 м, НИС “Академик Курчатов”, рейс 11, ст. 835, РТАК 19, проба 57, 16.11.1971 г.; ИО РАН – 1 экз. SL 225 мм, банка Сложная, НИС “Эврика”, трал 137, 05.04.1975 г.
P. caerulescens (3 экз.): ИО РАН – 1 экз. SL 114 мм, 14°17ʹ с.ш., 67°25ʹ з.д., горизонт лова 1500 м, НИС “Академик Курчатов”, рейс 14, ст. 1219, РТАК 23, время 05.45–09.32, 23.02.1973 г.; ИО РАН – 1 экз. SL 83 мм, 00°00ʹ1ʹʹ–00°09ʹ2ʹʹ ю.ш., 154°43ʹ4ʹʹ–154°22ʹ4ʹʹ з.д., НИС “Академик Курчатов”, рейс 17, ст. 1461; MNHN № 2003–1449 – 1 экз. SL 140 мм, Коралловое море, Новая Каледония (о-ва Луайоте), 24°58ʹ1.2ʹʹ ю.ш., 168°42ʹ0ʹʹ в.д., 1312–1520 м, рейс HALIPRO-2, ст. bt59, 18.11.1996 г.
P. coffea (6 экз.): ИО РАН – 1 экз. SL 125 мм, Андаманское море, РС “Орлик”, трал 39, проба 23, время 14.30, 16.03.1964 г.; ИО РАН – 1 экз. SL 120 мм, 00°01ʹ ю.ш., 175°55ʹ з.д., НИС “Витязь”, рейс 34, ст. 5139, РТАК, проба 347; ИО РАН SL ~160 мм, 00°01ʹ ю.ш., 175°55ʹ з.д., НИС “Витязь”, рейс 34, ст. 5139, РТАК, проба 350; ИО РАН – 2 экз. SL 55 и 70 мм, 03°08ʹ с.ш., 130°57ʹ в.д., 200 м, НИС “Витязь”, рейс 57, ст. 7285, РТАК № 79, проба 178, время 22.12–23.12, 25.03.1975 г. (экземпляры, уклоняющиеся к P. waitti); ИО РАН – 1 экз. SL 150 мм, 06°25ʹ с.ш., 140°40ʹ в.д., горизонт лова 500 м, НИС “Витязь”, рейс 57, ст. 7316, проба 212, из желудка Melanocetus johnsoni.
P. dinema (3 экз., изучены по фотографиям в разных проекциях): BMNH 1929.7.6.170-172 –синтипы SL ~ 24–28 мм из сборов НИС “Дана”, Северная Атлантика.
P. gorodinskii (1 экз.): ЗММУ № 23281 (из ИО РАН) – голотип, SL 180 мм, 07°55ʹ7ʹʹ с.ш., 153°45ʹ4ʹʹ з.д., НИС “Витязь”, рейс 34, ст. 5124, проба 273, 25.10.1961 г.
P. gracilis (1 экз.): ЗММУ № 23526 (из ИО РАН № 1702) – SL 155 мм, 12°23ʹ–12°33ʹ ю.ш., 48°07ʹ–48°05ʹ в.д., горизонт лова 720–700 м, НИС “Витязь-II”, рейс 17, ст. 2596, 11.11.1988 г.
P. leucospilus (2 экз., изучены по фотографиям в разных проекциях): BMNH 1929.7.6.173 – синтип SL ~ 25 мм, 14°52ʹ с.ш., 28°04ʹ з.д., НИС “Дана”, ст. 1161, трос 300 м; ZMUC 202063 – синтип SL 50 мм, 28°15ʹ с.ш., 56°00ʹ з.д., НИС “Дана”, ст. 1358, трос 2000 м.
P. litvinovi (1 экз.): ЗММУ № 23227 (из ИО РАН) – голотип, SL 150 мм, 02°20ʹ с.ш., 128°12ʹ в.д., горизонт лова 1000 м, НИС “Витязь”, рейс 57, ст. 7276, РТАК 72, проба 162, время 21.25–22.25, 23.03.1975 г.
P. margarita: в дополнение к 27 экз., перечисленным ранее (Прокофьев, 2015), изучено 8 экз.: ИО РАН – 2 экз. SL 207 и 220 мм, район Гавайских о-вов, сборы ТИНРО; ИО РАН – 1 экз. SL 181 мм, 00°01ʹ с.ш., 84°05ʹ з.д., горизонт лова 1000–0 м, НИС “Витязь”, рейс 35, ст. 5285, проба 414, время 22.40–00.20, 24–25.10.1962 г.; ИО РАН – 1 экз. SL 90.5 мм, 08°02ʹ с.ш., 142°16ʹ в.д., горизонт лова 200 м, НИС “Витязь”, рейс 57, ст. 7348, РТАК 109, проба 252; ИО РАН – 1 экз. SL 59 мм, Карибское море, горизонт лова 400 м, НИС “Витязь-II”, ст. 77, проба 6, РТАК 1, 02.05.1982 г.; ИО РАН – 1 экз. SL 45 мм, Карибское море, горизонт лова 500 м, НИС “Витязь-II”, ст. 150, проба 28, РТАК 5, время 19.35–20.35, 23.05.1982 г.; MNHN № 1991–6252 – 1 экз. SL 27 мм, 12°58ʹ ю.ш., 144°33ʹ в.д., горизонт лова 630 м, рейс Caride-I, ст. 9b, 13.09.1968 г.; MNHN № 2001-0594 – 1 экз. SL 270 мм, Индийский океан, 41°03ʹ ю.ш., 77°19ʹ58.8ʹʹ в.д., рейс KerAms, ст. c5, 04.08.2000 г.
P. mirabilis (1 экз.): ИО РАН – SL 83 мм, 35°29ʹ с.ш., 51°54ʹ з.д., горизонт лова 900–740 м, РТ “Атлант”, трал 201, 08.08.1976 г.
P. parvimanus (7 экз.): ИО РАН – 1 экз. SL 81 мм, 20°15ʹ ю.ш., 74°09ʹ з.д., НПС “Профессор Месяцев”, рейс 15, трал 275, 17.09.1984 г.; ИО РАН – 1 экз. SL 230 мм, 23°20ʹ7ʹʹ с.ш., 25°44ʹ з.д., РТАК 1000(0) м, НИС “Витязь-II”, рейс 9, ст. 1227, время 04.57–05.57, 25.05.1985 г.; ИО РАН – 1 экз. SL 163 мм, 29°41ʹ ю.ш., 164°39ʹ в.д., горизонт лова 200–0 м (550 м троса), НИС “Дмитрий Менделеев”, рейс 16, ст. 1249, РТАК 9, проба 18, время 00.15–01.15, 31.12.1975 г.; ИО РАН – 1 экз. SL 65 мм, 23°09ʹ ю.ш., 82°57ʹ з.д., НИС “Академик Курчатов”, рейс 4, проба 182; ИО РАН – 1 экз. SL 172 мм, 21°23ʹ2ʹʹ ю.ш., 81°38ʹ3ʹʹ з.д., НИС “Профессор Штокман”, рейс 18, ст. 1851, проба 33; ИО РАН – 1 экз. SL 140 мм, 32°03ʹ02ʹʹ с.ш., 168°01ʹ2ʹʹ з.д., НИС “Академик Мстислав Келдыш”, рейс 22, ст. 2332, РТАКСА, проба 18, время 19.40–21.50; ИО РАН – 1 экз. SL 204 мм, Срединно-Атлантический хребет, НИС “Академик Иоффе”, рейс 29, ст. 2188-1.
P. paxtoni (1 экз.): ИО РАН – SL 25 мм, 06°15ʹ5ʹʹ с.ш., 153°44ʹ7ʹʹ в.д., НИС “Витязь”, рейс 25, ст. 3663, 27–28.07.1957 г.
P. phyllopogon (6 экз.): ИО РАН – 1 экз. SL 23 мм, 04°28ʹ с.ш., 142°32ʹ в.д., горизонт лова ~180 м, НИС “Витязь”, рейс 50, ст. 6429, проба 54, РТАК, время 01.07–02.07, 03.05.1971 г.; ИО РАН – 1 экз. SL 38 мм, 02°02ʹ5ʹʹ–02°02ʹ3ʹʹ с.ш., 36°32ʹ2ʹʹ–36°31ʹ з.д., горизонт лова 700–0 м, НИС “Академик Сергей Вавилов”, рейс 43, ст. 2669, РТАКСА, время 23.04–00.39, 26–27.10.2016 г.; MNHN № 1993–3389 – 1 экз. SL 51 мм, 11°40ʹ1.2ʹʹ ю. ш., 143°10ʹ1.2ʹʹ в.д., 1300 м, рейс Caride-III, ст. 14; MNHN № 1993–3390 – 1 экз. SL 22 мм, 11°40ʹ1.2ʹʹ ю.ш., 143°10ʹ1.2ʹʹ в.д., 1300 м, рейс Caride-III, ст. 14; MNHN № 2003–2127 – 1 экз. SL 40 мм, Новая Каледония, 03°55ʹ1.2ʹʹ ю.ш., 170° в.д., 300 м, рейс Bora 1, ст. 6, 12.03.1965 г.; MNHN № 2017–0444 (из № 1991–6254) – 1 экз. SL 23 мм, 12°12ʹ ю.ш., 143°46ʹ1.2ʹʹ в.д., 1520 м, рейс Caride-I, ст. 17b.
P. uncinatus (1 экз.): ЗММУ № 23280 (из ИО РАН) – голотип SL 250 мм, возвышенность Риу-Гранди, горизонт лова 1070 м, НИС “Ихтиандр”, рейс 2188.XIII, трал 62, 20.02.1989 г.
P. venetaenia (1 экз.): ЗММУ № 23811 (из ИО РАН) – голотип SL 89 мм, 01°15ʹ ю. ш., 142°46ʹ в.д., горизонт лова 1000 м, НИС “Витязь”, рейс 57, ст. 7318, РТАК 93, проба 216, время 22.21–23.21, 05.04.1975 г.
P. waitti (26 экз.): ИО РАН – 1 экз. SL 76 мм, 205-“3”, проба 371 (район Куросио?); ИО РАН – 1 экз. SL 28 мм, НИС “Витязь”, рейс 25, ст. 3663, 28.07.1957 г., сачок закидной, проба 126; ИО РАН – 1 экз. SL 22.5 мм, 11°18ʹ5ʹʹ с.ш., 142°18ʹ в.д., НИС “Витязь”, рейс 27, ст. 4002, проба 312, плейстонный трал, слой облова – поверхность (глубина 9660 м), время 19.30–19.50, 21.05.1958 г.; ИО РАН – 1 экз. SL 41 мм, 04°24ʹ9ʹʹ–04°25ʹ1ʹʹ с.ш., 62°04ʹ1ʹʹ–62°04ʹ8ʹʹ в.д., НИС “Витязь”, рейс 33, ст. 4825, проба 224, ПС, слой облова – поверхность, время 21.45–22.00, 14.11.1960 г.; ИО РАН – 1 экз. SL 33 мм, 01°57ʹ1ʹʹ ю.ш, 83°03ʹ в.д., НИС “Витязь”, рейс 33, ст. 4911, проба 508, ПС, слой облова – поверхность, время 23.00–23.20, 10.01.1961 г.; ИО РАН – 1 экз. SL 30 мм, 07°45ʹ с.ш., 120°25ʹ в.д., горизонт лова 1500–0 м (над глубиной 4062 м), НИС “Витязь”, рейс 33, ст. 5028, проба 913, ринг-трал, время 22.00–02.50, 04–05.04.1961 г.; ИО РАН – 1 экз. SL 35 мм, 14°13ʹ ю.ш., 140°07ʹ8ʹʹ з.д., НИС “Витязь”, рейс 34, ст. 5105, проба 180, ПС, слой облова – поверхность, на свет, время 21.15–21.35, 29.09.1961 г.; ИО РАН – 1 экз. SL 38 мм, 01°59ʹ5ʹʹ с.ш., 176°01ʹ з.д., НИС “Витязь”, рейс 34, ст. 5137, проба 337, время 22.10–00.30, 07–08.11.1961 г.; ИО РАН – 2 экз. SL 29 и 30 мм, 01°59ʹ5ʹʹ с.ш., 176°01ʹ з.д., НИС “Витязь”, рейс 34, ст. 5137, проба 336, ПС, слой облова – поверхность, на свет, время 22.10–22.20, 07.11.1961 г.; ИО РАН – 1 экз. SL 37 мм, 05°59ʹ ю.ш., 176°13ʹ7ʹʹ з.д., НИС “Витязь”, рейс 34, ст. 5144, проба 365, ПС, слой облова – поверхность, на свет, время 00.45–01.00, 12.11.1961 г.; ИО РАН – 7 экз. SL 25–30 мм, 05°05ʹ с.ш., 77°07ʹ в.д., НИС “Витязь”, рейс 35, ст. 5249, проба 283, ПС, слой облова – поверхность, на свет, время 01.20–01.35, 30.11.1962 г.; ИО РАН – 3 экз. SL 27–31 мм, 00°01ʹ с.ш., 84°05ʹ з.д., НИС “Витязь”, рейс 35, ст. 5285, проба 420, ПС, на свет, время 19.40–19.55, 25.10.1962 г.; ИО РАН – 2 экз. SL 41 и ~ 44 мм, Бенгальский залив, 15°59ʹ с.ш., 89°37ʹ в.д., НИС “Витязь”, рейс 36, ст. 5308, сачок, слой облова – поверхность, время 18.30–19.00, 01.11.1964; ИО РАН – 1 экз. SL 26.5 мм, 07°50ʹ ю.ш., 120°30ʹ в.д., НИС “Витязь”, рейс 57, ст. 7254, проба 119, БНТ, слой облова – поверхность, время 03.20–03.40, 16.03.1975 г.; ИО РАН – 1 экз. SL 47 мм, 01°15ʹ ю.ш., 142°46ʹ в.д., НИС “Витязь”, рейс 57, ст. 7318, проба 217, БНТ, слой облова – поверхность, 05.04.1975 г.; ИО РАН – 1 экз. SL 27 мм, 01°25ʹ ю.ш., 142°48ʹ в.д., НИС “Витязь”, рейс 57, ст. 7319, БНТ, слой облова – поверхность, 06.04.1975 г.
Данные по P. achirus, P. corynodes и P. munificus, отсутствующие в доступном мне материале, взяты из их первоописаний (Regan, Trewavas, 1930; Gibbs, 1968; Klepadlo, 2011) и из переописания P. achirus (Flynn, Klepadlo, 2012). При анализе признаков и составлении определительной таблицы учтены данные первоописаний всех видов и основных таксономических работ по роду (Regan, Trewavas, 1930; Morrow, Gibbs, 1964; Klepadlo, 2011).
Методика изучения и терминология соответствуют общепринятой (Morrow, Gibbs, 1964); уточнения, касающиеся номенклатуры озубления челюстей и записи расположения фотофоров серии IP, подробно рассмотрены в соответствующих разделах анализа признаков. Для формального описания видов критически важными являются признаки озубления челюстей, строения подбородочного усика, наличие или отсутствие лучей Р, их число и строение, конфигурация фотофоров серии IP, а также форма и размеры посторбитального органа. При наличии на теле участков голубой светящейся ткани характер расположения последних является видоспецифичным. Дополнительное значение для видовой диагностики имеет степень развития жаберных лепестков, прежде всего, на ceratobranchiale-1 (хотя индивидуальная и экогеографическая внутривидовая изменивость по данному признаку нуждается в специальном изучении); наличие и характер расположения пятен белой светящейся ткани на голове и туловище; число нёбных зубов; положение на теле V и длина лучей в них (но эти лучи часто обломаны); число позвонков и другие меристические признаки. Из-за особенностей строения кожи и подкожной соединительной ткани у взрослых особей видов из подрода Trachinostomias и из группы “braueri” номинотипического подрода кожа у музейных образцов, как правило, сорвана, даже у добытых таким щадящим орудием лова, как трал Самышева–Асеева. При этом кожа всегда прочно соединена соединительнотканным тяжом, тянущимся по средней линии брюха так, что при её срыве лоскуты кожи обычно заворачиваются по ходу этого тяжа. В результате почти всегда можно подсчитать число органов в вентральном ряду фотофоров от истмуса до анального отверстия, однако невозможна оценка их числа в более задних отделах вентрального ряда и в боковом ряду, а также оценка наличия и характера расположения светящейся ткани на теле. Тем не менее я считаю возможным формальное описание новых видов по такому материалу при условии сохранности у него других структур из перечисленных выше, так как в противном случае таксономическое разнообразие в указанных группах окажется сильно недооценённым, что в свою очередь обесценит любые сравнительно-анатомические и филогенетические исследования данного рода. Особенно это касается подрода Trachinostomias, у видов которого признаки, исчезаемые с утратой кожи, достаточно однообразны, тогда как остеологические различия, судя по замечанию Финка (Fink, 1985), могут быть значительными. В указанных группах кожа лучше сохраняется у мальков, чем у взрослых рыб, однако структурные особенности (за исключением числа серийных фотофоров) могут изменяться в ходе онтогенеза, в связи с чем в качестве голотипов новых видов предпочтительнее выбирать взрослых рыб, даже при утрате у них кожи. На лучах вертикальных плавников кожа (при её срыве с туловища) почти всегда сохраняется в виде лоскутов в передней половине плавников и на межлучевой перепонке. Лучи Р, том случае когда они достаточно жёсткие, у коллекционных рыб также часто обломаны; в этом случае необходимо оценивать толщину их основания, благодаря которой можно предположить степень развития луча.
Подсчёт числа позвонков выполнялся по рентгенограммам в боковой проекции, по ним же проверялся подсчёт числа лучей в вертикальных плавниках. Пропорции посторбитального органа устанавливались по границам фотогенной железы. Пол и степень зрелости гонад устанавливались по макроскопическим признакам (Bucholtz et al., 2008). Все иллюстрации выполнены автором.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Обзор признаков, значимых для надвидовой классификации Photonectes
Озубление. Диагностически значимые отличия найдены в особенностях озубления praemaxillare и dentale (рис. 1) и числе зубов palatinum. В пределах рода Photonectes может быть выделено четыре типа озубления praemaxillare: 1) гетерогенный унисериальный (рис. 1а), 2) гомогенный унисериальный (рис. 1б–1д, 1и), 3) гетерогенный бисериальный (рис. 1е–1з, 1о–1р) и 4) гомогенный бисериальный (рис. 1к, 1л). Первые два типа характерны соответственно для молоди и взрослых особей видов подрода Trachinostomias, а последние два – для Photonectes s. str. в принимаемом здесь объеме. Озубление dentale может быть гомогенным (рис. 1б–1г, 1и, 1к) и гетерогенным (рис. 1м–1о, 1р), при этом оба варианта всегда сопряжены с таким же озублением praemaxillare.

Рис. 1.
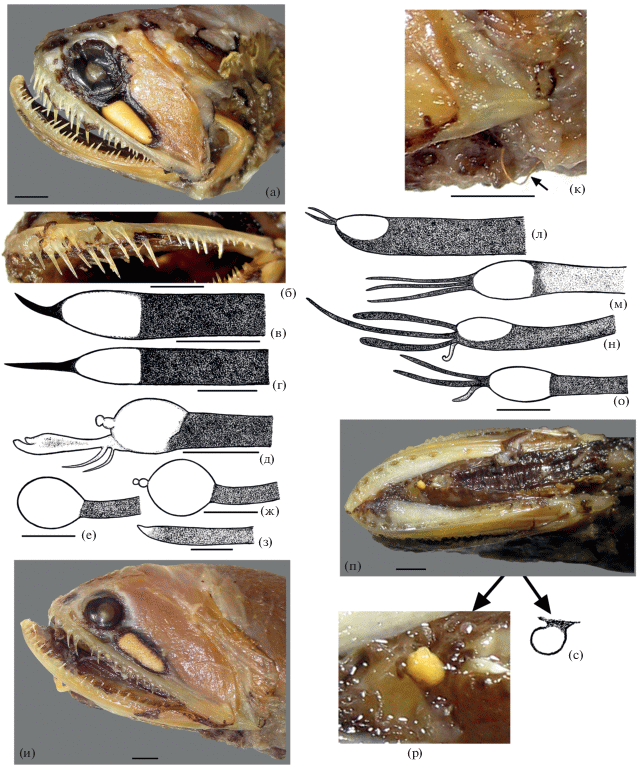
Особенности озубления Photonectes spp.: а – P. margarita SL 59 мм, НИС “Витязь-II”, ст. 77; б, в – P. margarita SL 192 мм, ЗММУ № 23290; г – P. albipennis SL 242 мм, Куросио, банка № 13; д – P. albipennis SL 170 мм, НИС “Витязь”, ст. 7400, praemaxillare; е – P. dinema SL 28 мм, BMNH 1929.7.6.172, praemaxillare; ж – P. albipennis SL 55 мм, Куросио, 205-“3”, praemaxillare; з – P. phyllopogon SL 38 мм, НИС “Сергей Вавилов”, ст. 2669, praemaxillare; и, к – P. albipennis SL 209 мм, НПС “Фиолент”, рейс 3, трал 177 (и – унисериальное озубление praemaxillare с левой стороны; к – бисериальное с правой стороны); л – P. albipennis SL 187 мм, НИС “Витязь”, ст. 7277, praemaxillare; м – P. albipennis SL 55 мм, Куросио, 205-“3” (выделены вершины отдельных премаксиллярных зубов, увел. ×2); н – P. dinema SL 28 мм, BMNH 1929.7.6.172, dentale; о, п – P. caerulescens SL 140 мм, MNHN № 2003–1449; р – P. braueri 225 мм SL, банка Сложная; с–ф – схемы озубления praemaxillare, основания зубов в ряду соединены прямой линией (с – P. caerulescens SL 140 мм; т – P. braueri SL 225 мм; у – P. uncinatus SL 250 мм; ф – P. braueri SL 50 мм); х–ч – строение вершин премаксиллярных зубов (х – P. caerulescens SL 140 мм, ц – P. braueri SL 50 мм, ч – P. phyllopogon SL 38 мм). Стрелками показаны зубы наружного ряда на praemaxillare. Масштаб: а–д, и–л, о–ч – 3 мм; е, н – 1 мм, ж, з, м – 1.5 мм.

Гомогенным я называю такой тип озубления челюстей, при котором все зубы имеют сопоставимые размеры и соседние зубы не различаются по величине. Разумеется, все зубы в челюсти не могут быть абсолютно одинаковыми, они мельче близ симфизов и к концу зубного ряда, но в целом озубление выглядит однородным (рис. 1б–1г). При гетерогенном озублении соседние зубы резко отличаются по длине. Все зубы в челюсти могут быть расположены строго в один ряд: если провести линию через центры оснований зубов, то она окажется прямой – унисериальный тип озубления. Однако в процессе настоящего исследования было выявлено, что у многих видов Photonectes от одного до четырёх зубов располагаются кнаружи от основного ряда, образуя второй (внешний) ряд зубов – бисериальный тип озубления. Когда зубы внешнего ряда располагаются напротив зубов внутреннего ряда (например, P. caerulescens, P. distichodon), двурядность озубления praemaxillare представляется очевидной (рис. 1о), однако в других случаях (P. braueri, P. uncinatus) зубы внешнего ряда располагаются против промежутков между зубами внутреннего ряда (рис. 1р). В этом случае может казаться, что чёткого подразделения на два ряда нет (как, например, было отмечено в первоописании P. uncinatus – Прокофьев, 2015. С. 137), но это не так. Если провести линии через центры оснований наружных и внутренних зубов, то они образуют две более или менее параллельные линии (рис. 1с–1у). Зубы внешнего ряда во всех изученных случаях разделены на две группы – переднюю и заднюю (в каждой из которых может быть один или два зуба). Зубы внешнего ряда всегда мельче наибольших зубов внутреннего ряда, поэтому при бисериальном типе гомогенность или гетерогенность озубления устанавливается по зубам внутреннего ряда.
Взрослые особи всех видов подрода Trachinostomias (табл. 1) имеют только гомогенное унисериальное озубление челюстей. Ювенильные особи были исследованы только у P. margarita, у них озубление praemaxillare резко гетерогенное, но также унисериальное (рис. 1а). Озубление dentale гетерогенное, причем заметно увеличены один–два зуба позади самых передних (присимфизных) зубов, тогда как все прочие зубы не сильно различаются по длине11 (рис. 1а). Гетерогенность озубления praemaxillare и dentale прослеживается у P. margarita до достижения по меньшей мере SL 70 мм (ЗММУ № 23293).
Гетерогенное бисериальное озубление praemaxillare установлено для P. braueri, P. caerulescens, P. dinema, P. distichodon, P. gracilis, P. leucospilus, P. litvinovi, P. mirabilis, P. paxtoni, P. phyllopogon, P. venetaenia и P. uncinatus. К сожалению, из всех этих видов я располагаю сведениями о возрастной изменчивости озубления praemaxillare только для P. braueri и P. caerulescens. Молодь P. braueri SL 22–50 мм имеет менее чёткое подразделение внутреннего и внешнего рядов, чем взрослый экз. SL 225 мм. У самых мелких рыб кнаружи смещен лишь один зуб, у малька P. braueri SL 50 мм (НИС “Витязь”, ст. 4577) слегка смещаются еще два зуба, но линия, соединяющая центры оснований смещённых кнаружи зубов, еще не параллельна таковой зубов внутреннего ряда, как это наблюдается у взрослых рыб (рис. 1ф). У имеющегося в моем распоряжении малька P. paxtoni SL 25 мм один зуб на praemaxillare смещён кнаружи. У экземпляров типовой серии P. dinema, представленной молодью SL 24–38 мм, озубление которых исследовал по моей просьбе Й. Нильсен (J. Nielsen, ZMUC), бисериальное озубление praemaxillare однозначно установлено для экземпляров ZMUC № 202061 и 202062, тогда как у остальных рыб (ZMUC № 202058-60) предположительно имеется только один ряд гетерогенных зубов (но подчеркивается, что сохранность челюстей плохая – “dentigerous bones are more or less damaged probably due to the many fangs being entangled in the net-meshes”: Й. Нильсен, личное сообщение). Из трёх синтипов этого вида, хранящихся в коллекции BMNH, у экземпляров SL 24 и 25 мм praemaxillare утрачено, а у экземпляра SL 28 мм озубление отчётливо бисериальное (рис. 1е). У неполовозрелого экземпляра P. caerulescens SL 83 мм (НИС “Академик Курчатов”, ст. 1461) только один зуб смещён кнаружи, причём в разной степени с правой и с левой сторон, тогда как половозрелые экземпляры, по крайней мере от 114 мм SL (НИС “Академик Курчатов”, ст. 1219), имеют чётко бисериальное озубление с двумя зубами в наружном ряду (рис. 1о, 1п). С другой стороны, малёк P. phyllopogon SL 38 мм уже имеет чётко бисериальное озубление praemaxillare (7 + 4) (рис. 1з), из чего можно предполагать, что дефинитивное озубление челюстей разными видами приобретается, возможно, на разных стадиях онтогенеза. Остальные из перечисленных выше видов представлены в моём материале половозрелыми рыбами с чётко бисериальным озублением praemaxillare. Таким образом, можно предполагать, что у мальков рассматриваемых видов сначала формируется гетерогенное унисериальное озубление челюстей, но уже на ранних стадиях (по меньшей мере при SL 20 мм) наблюдается формирование внешнего ряда, причём сначала кнаружи смещается зуб из передней группы, а затем – задней, и скорость преобразований в онтогенезе разных видов может различаться. Дефинитивным состоянием является гетерогенное бисериальное озубление praemaxillare. Озубление dentale у этих видов гетерогенное на всех возрастных стадиях.
Иначе осуществляется онтогенетическое преобразование озубления praemaxillare и dentale у видов группы “albipennis”, включающей P. albipennis, P. barnetti, P. coffea и P. waitti. У молоди P. albipennis и P. waitti SL 22–76 мм озубление praemaxillare и dentale гетерогенное и praemaxillare – бисериальное (с 1 + 1–2 зубами во внешнем ряду) (рис. 1ж, 1м). С ростом размер челюстных зубов уменьшается и при SL > 100 мм озубление становится гомогенным (рис. 1г, 1д, 1и–1л). Нужно отметить, что при этом на praemaxillare нередко удаётся различить две группы зубов, разделённых небольшим (часто незначительным) промежутком (рис. 1г), но зубы обеих групп не различаются по величине. У других рыб такой промежуток совершенно отсутствует (рис. 1и, 1к). В одних случаях присимфизные зубы praemaxillare не отделены промежутком от последующих и мало отличаются от них по величине (рис. 1л), в других один–два присимфизных зуба заметно мельче и отделены от последующих промежутком (рис. 1и). Во внешнем ряду у взрослых рыб имеется по одному–два зуба с каждой стороны (1 + 0 или 1 + 1 в передней и задней группах), по крайней мере до SL 245 мм (БМРТ “Таджикистан”, 24°04ʹ1 ю.ш., 112°27ʹ в.д.). Но у части взрослых рыб озубление praemaxillare становится гомогенным унисериальным (рис. 1г, 1д) (искусственная утрата внешних зубов здесь может быть уверенно исключена). Наименьший из имеющихся в моём распоряжении экземпляров с таким типом озубления имеет SL 188 мм (БМРТ “Академик Берг”, 09°07ʹ ю.ш., 131°02ʹ в.д.). Изредка встречаются рыбы, у которых praemaxillare с одной стороны имеет унисериальное (рис. 1и), а с другой – бисериальное (рис. 1к) озубление.
У P. barnetti озубление челюстей и последовательность его изменения в онтогенезе такое же, как описано выше, но зубы на praemaxillare и maxillare в целом несколько длиннее, чем у экземпляров других видов группы “albipennis” при тех же размерах. Но поскольку среди рыб, по строению подбородочного усика однозначно определяемых как P. albipennis, также наблюдается некоторая изменчивость в величине челюстных зубов, мне представляется, что по этой особенности уверенно разделить эти два вида всё-таки нельзя. И всё же, даже если зубы у P. barnetti в среднем более длинные, озубление praemaxillare тем не менее гомогенное и, как правило, бисериальное (унисериальное у экземпляра SL 124 мм, НИС “Дмитрий Менделеев”, ст. 1531). В оригинальном описании P. barnetti указано, что на praemaxillare и maxillare чередуются короткие и длинные клыки, но на схеме озубления все зубы изображены практически одноразмерными (Klepadlo, 2011. Fig. 3B). В моём материале все взрослые рыбы (SL 112–160 мм) с подбородочным усиком характерного для P. barnetti строения (включая топотипный экземпляр с банки Милуоки) имеют гомогенное озубление praemaxillare, maxillare и dentale. P. waitti известен только по неполовозрелым экземплярам (SL до 76 мм) с гетерогенным озублением praemaxillare и dentale. В оригинальном описании (Flynn, Klepadlo, 2012) рядность зубов praemaxillare не указана, но, судя по приводимой фотографии голотипа, они бисериальные. В моём материале все экземпляры этого вида имеют гетерогенное бисериальное озубление praemaxillare и гетерогенное озубление dentale, подобные соразмерным малькам P. albipennis. Таким образом, онтогенез озубления praemaxillare и dentale у видов группы “albipennis” существенно отличается от такового у P. braueri, P. caerulescens и близких к ним видов. Уже у наименьших из исследованных мальков (SL 22.5–23.5 мм) озубление praemaxillare чётко бисериальное, как и dentale гетерогенное, но по достижении половой зрелости гетерогенное озубление praemaxillare и dentale сменяется гомогенным, а внешний ряд зубов на praemaxillare у части рыб исчезает.
Конечно, для уверенной оценки филогенеза озубления челюстей у Photonectes пока что недостаточно фактических данных, но всё же мне представляется допустимой следующая рабочая гипотеза. Детали озубления челюстей различных родов меланостомовых рыб остаются малоизученными, однако формирование внешнего ряда более мелких зубов отмечено, помимо Photonectes, и у некоторых других, не связанных близким родством родов (например, у Chirostomias, Echiostoma, Malacosteus). Известен и высокоспециализированный по признакам озубления род, имеющий многочисленные ряды зубов на praemaxillare и dentale (Tactostoma). Гомо- и гетерогенный тип озубления praemaxillare и dentale также распределён довольно мозаично. Вместе с тем наиболее типичным для стомиатоидных рыб является гетерогенное унисериальное озубление praemaxillare и dentale, характерное, в частности, для наиболее примитивных представителей группы (Astronesthes, Borostomias). Хотя не известно ни одного вида Photonectes, у которого бы в дефинитивном состоянии сохранялось гетерогенное унисериальное озубление praemaxillare, этот тип озубления характерен для молоди видов Trachinostomias. Во взрослом состоянии в этом подроде он сменяется гомогенным унисериальным озублением. Внешний ряд зубов на praemaxillare у видов Trachinostomias никогда не формируется. Напротив, гетерогенное озубление praemaxillare и dentale и бисериальное озубление praemaxillare, по-видимому, являются исходным у подрода Photonectes s. str., но в группе видов “albipennis” вторично осуществляется переход к гомогенному озублению praemaxillare и dentale и наблюдается тенденция к унисериальности озубления praemaxillare. Гомогенное и в отдельных случаях унисериальное (на praemaxillare) озубление челюстей у видов группы “albipennis” возникает параллельно и независимо от такового у Trachinostomias, в пользу чего свидетельствует тот факт, что у видов группы “albipennis” на ранних онтогенетических стадиях озубление praemaxillare всегда бисериальное, тогда как у Trachinostomias – унисериальное. Наблюдаемое у видов группы “albipennis” только на ранних онтогенетических стадиях гетерогенное озубление praemaxillare и dentale является дефинитивным для видов комплекса “braueri–caerulescens”, с какими-то из представителей которого виды группы “albipennis”, очевидно, филогенетически связаны. Таким образом, специфика озубления подродов Trachinostomias и Photonectes s. str. (в принимаемом здесь объеме) должна была сформироваться на самых ранних этапах кладогенеза группы. Вероятно, предковая форма Photonectes обладала гетерогенным унисериальным типом озубления, сохраняющимся у существующих видов лишь у молоди Trachinostomias, на основании которого сформировалось, с одной стороны, дефинитивное гомогенное унисериальное озубление Trachinostomias, а с другой, гетерогенное бисериальное озубление P. braueri, P. caerulescens и родственных им видов. От последних происходят виды группы “albipennis”, вторично приобретающие гомогенное озубление praemaxillare и dentale в дефинитивном состоянии.
Помимо числа рядов и соотношения размеров соседствующих зубов в челюстях представляет интерес строение вершин отдельных зубов, прежде всего на praemaxillare. У молоди всех видов фотонектов близ вершины зуба имеется хорошо развитый дополнительный зубчик (“barb”) (рис. 1м, 1х). У мальков Trachinostomias (P. margarita), по меньшей мере до 70 мм SL, дополнительные зубчики хорошо развиты на всех зубах praemaxillare и vomer, на многих зубах maxillare, dentale и palatinum. У взрослых особей всех видов Trachinostomias вершины всех зубов простые, заострённые (рис. 1б, 1в). У видов подрода Photonectes s. str. дополнительные зубчики обычно сохраняются и во взрослом состоянии. У ювенильных особей P. braueri выявлена изменчивость в степени развития дополнительных зубчиков: иногда (особь SL 50 мм, НИС “Витязь”, ст. 4577) зубчик в виде закруглённой лопасти слабо развит (рис. 1ц), присутствует на всех премаксиллярных зубах, но на других костях вершины зубов простые. У другого малька (SL 45 мм, НИС “Академик Курчатов”, ст. 835) все премаксиллярные и сошниковые зубы с отчётливыми дополнительными зубчиками, очень слабые зубчики присутствуют также на отдельных зубах maxillare и dentale. У взрослых P. braueri и P. distichodon на praemaxillare и maxillare имеются отдельные зубы с более или менее выраженными зубчиками, тогда как остальные зубы с простыми вершинами. У P. caerulescens, P. dinema, P. leucospilus, P. mirabilis, P. paxtoni,22 P. venetaenia, P. uncinatus и молоди видов группы “albipennis” дополнительный зубчик хорошо развит на всех премаксиллярных зубах (рис. 1м), а у видов группы “albipennis” иногда и на некоторых зубах maxillare. У половозрелых экземпляров видов группы “albipennis” этот дополнительный зубчик обычно также сохраняется, но у части рыб исчезает, причём его исчезновение коррелирует с максимальным уменьшением размеров зубов и с утратой внешнего ряда на praemaxillare. Зубы нижней челюсти у этих видов только с простыми вершинами, но сошниковые и нёбные зубы также могут нести дополнительные зубчики. Только у P. gracilis вершины зубов, по-видимому, простые (но у единственного имеющегося в моём распоряжении экземпляра у части зубов они обломаны). У P. phyllopogon при SL 38 мм вершины всех премаксиллярных зубов крючковидно загнуты назад, но довольно слабый дополнительный зубчик присутствует лишь на некоторых из них (рис. 1з, 1ч).
Таким образом, у видов Trachinostomias дополнительные зубчики на зубах исчезают в процессе развития, тогда как у видов номинотипического подрода они сохраняются, по крайней мере на премаксиллярных зубах. Вместе с тем степень их развития у разных видов Photonectes s. str. весьма неоднородна и, по-видимому, есть виды, у которых они также могут быть совсем утрачены (P. gracilis).
Число зубов на palatinum составляет (4)5–6 у видов подрода Trachinostomias, у P. braueri и P. distichodon; как исключение и только с одной стороны у P. margarita встречно три зуба. Нёбные зубы у этих видов более или менее расставлены, так что длина их ряда больше длины наибольшего зуба. У всех остальных видов на нёбных костях обычно имеется два или три сближенных основаниями зуба; как исключение и только с одной стороны у отдельных ювенильных экземпляров группы “albipennis” отмечено 4 зуба33. Однако редукция числа нёбных зубов, вероятнее всего, происходит неоднократно, поскольку имеющий лишь по два нёбных зуба P. uncinatus по совокупности признаков стоит ближе всего к P. braueri и P. distichodon, а не к какому-либо из видов с малым числом нёбных зубов.
Посторбитальный и суборбитальный органы. Форма и/или размеры посторбитального органа могут быть видоспецифичными, хотя изменчивость этих признаков недостаточно изучена. Размеры посторбитального органа подвержены половому диморфизму и, по крайней мере у некоторых видов (P. xenopogon), по-видимому, также значительной онтогенетической изменчивости. Вместе с тем я обнаружил признак, который стабилен у обоих полов во всех возрастах и, вероятно, может считаться филогенетически значимым. Он отмечен только у видов, имеющих рисунок из голубой светящейся ткани на теле (что само по себе является очень своеобразной особенностью), и является дополнительным аргументом в пользу возможной монофилии этой линии, уже предполагавшейся ранее (Прокофьев, 2015).
Масса светящейся железы у меланостомовых рыб снаружи окружена кольцом чёрной ткани. У P. caerulescens, P. gracilis, P. mirabilis и P. venetaenia передненижний угол этого окаймления сильно выдается вперёд, достигая вертикали середины глаза (рис. 2а). Судя по приводимой фотографии, хотя и плохого качества, такой же выступ имеется и у P. achirus, однако здесь он не заходит за вертикаль заднего края глаза (но изображён малёк SL 32.3 мм со слабо развитым посторбитальным органом: Flynn, Klepadlo, 2012. Fig. 4). У других видов рода передние верхний и нижний углы либо одинаково закруглены, либо нижний лишь едва выдается вперёд по сравнению с верхним (рис. 2б, 2в). Оказалось возможным проанализировать возрастную изменчивость данного признака у P. braueri – вида, наиболее сходного с “голубыми” фотонектами по характеру озубления челюстей и степени захождения кожи на вертикальные плавники. У молоди этого вида (SL 50 мм) передненижний угол окаймления посторбитальной железы слабо выступает вперёд, слегка заходя за вертикаль заднего края глаза, тогда как у взрослых рыб передний край окаймления вертикальный. В то же время у малька близкого к P. braueri вида P. leucospilus при SL ~ 25 мм передний край посторбитального органа вертикальный, с одинаково закруглёнными верхним и нижним углами. У видов Trachinostomias и группы “albipennis” строение посторбитального органа одинаковое на всех возрастных стадиях и изменяется лишь его размер (увеличивается с ростом). У самок Photonectes длина посторбитального органа меньше, чем у самцов.
Рис. 2.
Детали строения Photonectes spp.: а–в – посторбитальный и суборбитальный (so) органы (а – P. caerulescens, MNHN № 2003–1449, стрелкой обозначен передненижний выступ окаймления железы посторбитального органа; б – P. margarita, ЗММУ № 23290; в – P. albipennis, НИС “Витязь”, ст. 7400); г–и – схемы расположения фотофоров серии IP (г – P. margarita, д – P. uncinatus, e – P. gracilis, ж – P. caerulescens, з – P. braueri, и – P. albipennis, взрослый); к, л – расположение органов IP у мальков P. albipennis SL 32.5 мм, НИС “Витязь”, ст. 4564 (к) и P. leucospilus SL 25 мм, BMNH 1929.7.6.173 (л) (стрелкой показан промежуток между передней и задней группами IP); м–о – варианты строения Р, схематично (м – P. distichodon, н – P. xenopogon, о – P. filipendulus); п–ф – характер захождения кожи на лучи D и А (п – P. caerulescens SL 140 мм, MNHN № 2003–1449; р – P. caerulescens SL 114 мм, НИС “Академик Курчатов”, ст. 1219; с – P. phyllopogon SL 38 мм, НИС “Сергей Вавилов”, ст. 2669; т – P. braueri SL 50 мм, НИС “Пётр Лебедев”, IV–123А; у – P. albipennis SL 32.5 мм, НИС “Витязь”, ст. 4564; ф – P. albipennis SL 170 мм, НИС “Витязь”, ст. 7400; белыми стрелками обозначены основания лучей, черными – верхняя граница кожи на том же луче). Масштаб: а–и, м–ф – 3 мм; к, л – 2 мм.

Рис. 3.
Детали строения Photonectes spp.: а–л – P. margarita, варианты строения усика (а – ЗММУ № 23294, б – ЗММУ № 23283, в – SL 220 мм, район Гавайских о-вов, сборы ТИНРО; г – ЗММУ № 23293, SL 73 мм; д – НИС “Витязь”, ст. 5285; е – ЗММУ № 23290, ж – ЗММУ № 23285, з – НИС “Витязь”, ст. 7348; и – ЗММУ № 23293 SL 91 мм; к – ЗММУ № 23293, SL 80 мм; л – ЗММУ № 23295); м–о – P. parvimanus, варианты строения усика (м – малёк, SL не указана, по: Regan, Trewavas, 1930. Fig. 119A ); н – SL 81 мм, НПС “Профессор Месяцев”, рейс 15, трал 275; о – SL 163 мм, НИС “Дмитрий Менделеев”, ст. 1249); п – P. albipennis с укороченными жаберными лепестками на 1–й дуге, НИС “Витязь”, ст. 5005; р – P. albipennis со слабо пигментированным усиком, НИС “Витязь”, ст. 5103. Масштаб: а – 1.5 мм, л – 2 мм; б–к, м–р – 1 мм; линейка общая для з–к.
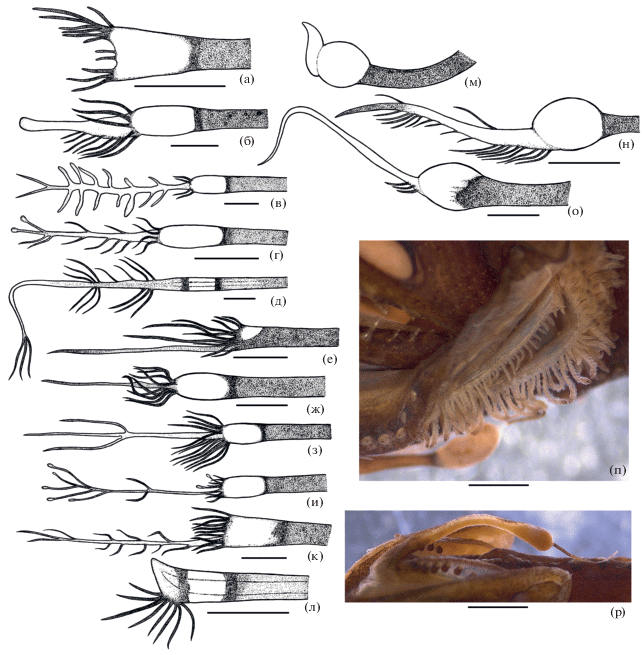
Рис. 4.
Детали строения Photonectes spp.: а–в – голотип, г – паратип P. distichodon sp. n. (а – вид головы сбоку, б – озубление praemaxillare; в, г – подбородочный усик); д–з – P. braueri, варианты строения усика (д – SL 22 мм, НИС “Витязь”, ст. 5028; е – SL 50 мм, НИС “Пётр Лебедев”, IV–123A; ж – SL 43 мм, НИС “Академик Курчатов”, ст. 835; з –SL 225 мм, банка Сложная); и–с – P. filipendulus sp. n. (и – вид головы сбоку, голотип; к – Р (показан стрелкой), голотип; л–о – подбородочный усик (л – голотип; м – паратип ИО РАН № 3590; н – паратип ИО РАН № 3589; о – паратип ИО РАН № 3588); п–с – аберрантное строение усика нетипового экземпляра). Масштаб: ф, б, и, к, п–с – 3 мм; в, з, л–о – 1 мм, г–ж – 0.5 мм; линейка общая для л–о.
Иногда в качестве диагностического признака рода Photonectes указывается отсутствие суборбитального органа (Klepadlo, 2011), однако я обнаружил его у всех видов как небольшой округлый фотофор под нижним краем глаза на уровне его середины (рис. 2а–2в). Ранее Финк отмечал утрату суборбитального органа только “у некоторых филогенетически продвинутых видов Eustomias” (Fink, 1985. P. 94).
Серии фотофоров IP и BR. Наличие или отсутствие промежутка в ряду IP в характеристике разных видов фотонектов отмечалось давно (Regan, Trewavas, 1930), хотя детально расположение органов в этом ряду описано и сравнено не было. Клепадло (Klepadlo, 2011) добавила к характеристике подродов величину промежутка от вершины истмуса до первой пары фотофоров IP – ряд протягивается на всю длину истмуса (Trachinostomias) либо начинается примерно от середины его длины (остальные подроды).
Анализ характера расположения фотофоров IP у различных видов Photonectes показал, что имеет место скоррелированная зависимость между величиной передних и задних фотофоров в этой серии, промежутков между отдельными органами в серии и расстояния от первой пары органов до вершины истмуса, на основании которой чётко разделяются подроды Trachinostomias и Photonectes s. str. в принимаемом здесь объеме. Внутри Photonectes s. str. выявлена некоторая изменчивость взаиморасположения органов в задней части серии IP. У видов Trachinostomias (P. filipendulus, P. margarita, P. parvimanus, P. xenopogon) все органы IP мелкие, одинакового размера, расположены на равных промежутках друг от друга, и их первая пара отстоит от вершины истмуса на расстояние, равное промежутку между органами первой и второй пары (рис. 2г). Межвидовая и онтогенетическая изменчивость по данным признакам отсутствует. Число органов в серии варьирует от 8 до 11 (Klepadlo, 2011), но, как правило, их 10 (у P. munificus – 8). У P. munificus органы в серии описаны без промежутка, положение первой пары относительно вершины истмуса не известно (Gibbs, 1968; Klepadlo, 2011).
В первоописании P. uncinatus расположение органов IP отмечено не было, однако этот вид был описан в составе подрода Trachinostomias, в диагнозе которого и в определительной таблице было указано отсутствие промежутка в серии IP (Прокофьев, 2015). Переисследование голотипа этого вида выявило у него довольно своеобразное расположение органов в данной серии (рис. 2д). Передние семь органов серии у него уменьшены в размерах и сближены, так что величина промежутков между ними не превышает их диаметра (самые передние органы чуть более расставлены, чем задние). Восьмой фотофор в серии несколько крупнее и отстоит от предыдущих на промежуток, равный 1.5 его диаметра, а от девятого – на промежуток, равный трём своим диаметрам. Два последних фотофора в серии разделены промежутком в полтора своих диаметра. Таким образом, промежутки между задними органами серии у P. uncinatus увеличены, но расстояние между 8-м и 9-м органами значительно превышает соседние промежутки, в связи с чем формула ряда может быть записана как 8 + 2. Расстояние от вершины истмуса до 1-й пары органов IP у P. uncinatus слегка превышает суммарную длину восьми передних фотофоров IP и промежутков между ними. Следовательно, по расположению органов IP этот вид не соответствует Trachinostomias и должен быть перемещён в подрод Photonectes s. str., где он наиболее близок к P. braueri.
У всех видов подрода Photonectes s. str. первая пара фотофоров IP смещена назад, так что промежуток между ней и вершиной истмуса равняется суммарной длине не менее чем шести–восьми передних фотофоров и промежутков между ними, а иногда даже немного превышает длину всего IP-ряда. В расположении органов IP-серии наблюдается некоторая межвидовая изменчивость. У P. caerulescens и P. gracilis передние шесть или семь органов сближены, так что промежутки между ними меньше диаметра фотофора, а три последних органа расставлены так, что формула ряда записывается как 6 + 1 + 1 + 1 или 7 + 1 + 1 + 1. При этом у исследованного мною экземпляра P. gracilis (6 + 1 + 1 + 1) расстояние между последним и предпоследним органами в серии больше (около трёх диаметров фотофора), чем промежутки между 6-м и 7-м и 7-м и 8-м органами в серии (около двух диаметров фотофора) (рис. 2). К сожалению, я не располагаю материалом для подтверждения стабильности данного состояния у P. gracilis. У P. caerulescens отмечена некоторая изменчивость по данному признаку: у одних рыб три последних фотофора расположены на равных промежутках (рис. 2ж), а у других промежуток между двумя последними органами соизмерим с их диаметром, тогда как промежуток между 7-м и 8-м и 8-м и 9-м органами явственно его превышает (формула записывается как 7 + 1 + 2).
Рис. 5.
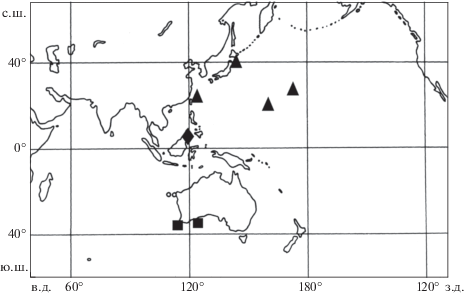
Распространение Photonectes distichodon (◼), P. filipendulus (▲) и P. xenopogon spp. n. (◆).
Видам P. braueri, P. dinema, P. distichodon, P. leucospilus, P. litvinovi, P. mirabilis, P. phyllopogon и P. venetaenia свойственно чёткое подразделение ряда IP на две группы – переднюю, состоящую из восьми (реже семи) сближенных органов, промежутки между которыми не превышают их диаметра, и заднюю, состоящую из двух столь же сближенных органов, но отстоящих от последнего фотофора передней группы на расстояние не менее двух диаметров органа (рис. 2з, 2и). Формула ряда записывается как 8 + 2 (или 7 + 2). Фотофоры задней группы могут быть несколько крупнее передних (что обычно проявляется у крупных рыб) или одинаковой с ними величины. Судя по исследованным малькам P. braueri, P. dinema и P. leucospilus (SL 22–50 мм), онтогенетическая изменчивость у этих видов отсутствует, органы расположены у них так же, как и у крупных рыб (P. braueri SL 225 мм). У видов группы “albipennis” (P. albipennis, P. barnetti, P. coffea, P. waitti) в дефинитивном состоянии (неясно для P. waitti, известного только по малькам) расположение органов такое же, как описано выше, но отмечены внутривидовые вариации числа органов в группах (обычно 8 + 2, реже 7 +2, как исключение 6 + 2, 8 + 3 или 7 + 1 + 2). Однако у мальков видов группы “albipennis” вплоть до SL 50–60 мм все органы IP расположены на равном расстоянии друг от друга (рис. 2к). Таким образом, в онтогенезе этих видов формирование промежутка в серии IP происходит только на поздних стадиях, что отличает их от видов группы “braueri”, имеющих этот промежуток уже на ранних стадиях (рис. 2л). Очевидно, молодь видов группы “albipennis” проходит стадию предкового состояния признака, когда смещение ряда назад от вершины истмуса уже произошло, но промежуток в задней части ряда еще не сформировался. У видов группы “braueri” эта стадия выпадает или, по меньшей мере, формирование дефинитивного состояния завершается уже по достижении SL 20 мм (рыбы меньших размеров не известны).
Кроме того, необходимо отметить разницу в числе фотофоров BR, которое у видов Trachinostomias составляет 9–14, у большинства видов комплекса “braueri–caerulescens” (кроме P. achirus и P. phyllopogon) – 8–10, у P. achirus и P. phyllopogon – 6–8, а у видов группы “albipennis” – лишь 5–7. Редукцию числа органов BR, очевидно, следует рассматривать в качестве апоморфной особенности, но, вероятнее всего, как и другие редуктивные преобразования в роде Photonectes (число нёбных зубов, лучей Р), она осуществляется неоднократно и независимо. Если в отношении P. phyllopogon (взрослые особи которого не описаны) еще можно предполагать филогенетическое родство с видами группы “albipennis”, то P. achirus, для которого указано 6–8 органов BR (Flynn, Klepadlo, 2012), совершенно очевидно близкородствен P. caerulescens (у последнего, по моим данным, BR 8–10). Возрастной изменчивости по числу органов BR на моём материале не прослеживается.
Число фотофоров в других группах может различаться у разных видов, но закономерностей, позволяющих объединять виды в естественные группы на основании числа и расположения органов в сериях, не выявлено.
Грудной плавник (Р). Наличие или отсутствие Р, число и степень развития его лучей (рис. 2м–2о) традиционно считаются важным признаком надвидовой классификации рода (Regan, Trewavas, 1930; Morrow, Gibbs, 1964; Klepadlo, 2011), но, на мой взгляд, он имеет диагностическое значение только на уровне видов, тогда как его филогенетический вес и, соответственно, значение для надвидовой классификации ничтожны. Число лучей Р у Photonectes варьирует от 0 до 3, при этом все переходы можно наблюдать в пределах подрода Trachinostomias. У P. (T.) gorodinskii и P. (T.) parvimanus два луча Р (судя по толщине сохранившихся оснований, у первого вида они, по-видимому, были развиты лучше, чем у второго). Как исключение, у отдельных особей P. parvimanus может быть встречен зачаточный третий луч Р, иногда только с одной стороны (НПС “Профессор Месяцев”, рейс 15, трал 275). У видов группы “braueri” из номинотипического подрода (P. braueri, P. dinema, P. distichodon, P. uncinatus) также два небольших луча Р (ранее на этом основании они выделялись в особый подрод Melanonectes). Наличие двух небольших лучей Р, очевидно, является исходным состоянием у Photonectes. У других видов Trachinostomias наблюдается либо сильное развитие верхнего луча (нижний при этом сохраняется, но остается слабо развитым либо утрачивается) (P. margarita, P. xenopogon), либо редукция плавника сначала до единственного крошечного луча (P. filipendulus), а затем и его полная утрата (у P. munificus – Gibbs, 1968). У всех прочих видов Photonectes s. str. Р отсутствует. Очевидно, что редукция Р осуществляется в подродах Trachinostomias и Photonectes s. str. независимо.
Кожа на вертикальных плавниках. Захождение кожи на вертикальные плавники (D и А) традиционно трактуется как специфическая (и основная) черта Trachinostomias (включая Microchirichthys (Regan, Trewavas, 1930)), отделяющая его от других совокупностей видов (Morrow, Gibbs, 1964; Klepadlo, 2011; Прокофьев, 2015). Между тем тщательное исследование экземпляров, находящихся в моём распоряжении, показало, что по этому признаку у разных видов наблюдается сложная изменчивость и при придании ему определяющего значения в диагностике подродов (как это всегда делалось ранее) возможны ошибки в определении родственных отношений видов. Например, P. uncinatus, характеризующийся полностью покрытыми кожей вертикальными плавниками, при первоописании был отнесён на этом основании к подроду Trachinostomias, хотя по совокупности других признаков он оказывается ближе всего к P. braueri. Более того, оказалось, что у некоторых видов (P. caerulescens) изменчивость по данному признаку такова, что одни экземпляры должны быть классифицированы по нему как Trachinostomias, а другие – как Photonectes s. str. (рис. 2п, 2р).
Было установлено, что видам Trachinostomias (P. margarita, P. parvimanus) свойственно захождение кожи не менее чем на 2/3 длины лучей D и А на всех стадиях онтогенеза. У взрослых рыб кожа, вероятно, доходит до кончиков лучей, однако проследить этот признак оказывается возможным исключительно редко, так как плавниковые лучи почти всегда сильно повреждаются. Среди других видов захождение кожи на вертикальные плавники вплоть до кончиков лучей отмечено ещё только у P. uncinatus. Однако у крупного экземпляра P. braueri (НИС “Эврика”, банка Сложная) кожа, по-видимому, также заходит не менее чем на 2/3 длины лучей D и А, но далее у него лучи сильно повреждены и размочалены, так что кожа на них могла быть просто содрана. Вообще, как правило, крупные экземпляры видов Trachinostomias и видов группы “braueri” повреждены настолько, что кожа на плавниках сохраняется лишь в виде обрывков, из-за чего оценивать степень захождения кожи на лучи не представляется возможным.
У P. caerulescens наблюдаются резкие различия по этому признаку между экземплярами SL 114 и 140 мм (оба являются половозрелыми рыбами, других значимых различий между ними не выявлено). У наиболее крупного экземпляра кожа практически целиком покрывает лучи D и А (на задних лучах она частично содрана, но, судя по самым последним лучам этих плавников, могла доходить практически до их кончиков) (рис. 2п). В то же время у экземпляра SL 114 мм кожа в задней половине D и A доходит лишь до середины длины лучей (рис. 2р). Однако даже это противоречит диагнозу подрода Photonectes s. str., к которому данный вид относится и для которого указывается отсутствие кожи на вертикальных плавниках (Morrow, Gibbs, 1964; Klepadlo, 2011). Нужно заметить, что исходная формулировка данного признака звучала несколько иначе (“membrane of dorsal and anal fins thick and black, only tips of rays visible in adults vs. thin, rays conspicuous”: Regan, Trewavas, 1930. P. 120–125), однако и при такой формулировке крупные экземпляры P. braueri и P. caerulescens в моей коллекции должны быть отнесены к Trachinostomias, а не к Melanonectes и Photonectes s. str., куда они помещаются в указанной выше классификации (Regan, Trewavas, 1930). Нужно также отметить, что у молоди Trachinostomias лучи вертикальных плавников гораздо лучше различимы, чем у взрослых, при сохранной коже, однако связано это не со степенью захождения кожи на плавники, а со степенью развития подкожного слоя студенистой соединительной ткани (см. далее), которая у молоди развита гораздо хуже, в связи с чем кожа плотно охватывает лучи.
У молоди всех видов Photonectes (за исключением Trachinostomias в принимаемом здесь объёме) кожа заходит лишь на основания лучей D и А, оставляя лучи открытыми не менее чем на 2/3 их длины (рис. 2с–2у). У молоди видов комплекса “braueri–caerulescens” при тех же размерах кожа на плавниках развита сильнее, чем в группе “albipennis”. В дальнейшем, по-видимому, у видов комплекса “braueri–caerulescens” происходит нарастание кожи на лучи, тогда как у видов группы “albipennis” лучи остаются совершенно свободными от кожи даже у самых крупных рыб (рис. 2ф).
Кроме того, следует отметить существенные различия в степени развития подкожного слоя соединительной ткани у различных представителей рода Photonectes. Он исключительно сильно развит у взрослых особей видов подрода Trachinostomias и видов группы “braueri” из номинотипического подрода. В связи с этим кожа у коллекционных экземпляров почти всегда сорвана. Напротив, у молоди всех видов рода подкожный слой развит довольно слабо и при SL < 80 мм кожа обычно остается сохранной. У видов группы “albipennis” подкожный слой остаётся слабо выраженным даже у самых крупных рыб, а сама кожа толще и грубее, в связи с чем материал хорошо сохраняется в тралах. Виды с голубой светящейся тканью на теле (группа “caerulescens”) занимают по данному признаку промежуточное положение между двумя вышеуказанными совокупностями. Функциональное значение данной особенности (гидродинамическое, защитное?) не вполне ясно. Представляет интерес сравнительное гистологическое исследование кожи и подкожной соединительной ткани у разных видов рода.
Число позвонков. По литературным данным (Morrow, Gibbs, 1964; Richards, 2006), число позвонков у видов Photonectes составляет 49–64. Эти данные не учитывают P. munificus, у которого 67 позвонков. По моему материалу пределы колебания числа позвонков составляют 48–63 (включая уростиль). В первоописании P. gorodinskii для этого вида было указано 53 или 54 позвонка, но рентгеноснимок рыбы, длительное время хранившейся в формалине, в передней части туловища был нечётким. Переделанный после четырёх лет хранения в спирту рентгеноснимок голотипа чётко показал наличие у него 57 позвонков (включая уростиль) (D 19, A 20, vert. 57, хвостовых 14). Таким образом, реальное число позвонков у этого вида, хотя и наименьшее среди Trachinostomias, не так резко отличается от других видов данного подрода, как казалось ранее. После исключения из состава данного подрода P. uncinatus (54 позвонка) оказывается, что между двумя принимаемыми мною подродами фотонектов имеются четкие (с хиатусом) различия по число позвонков – 48–55 у Photonectes s. str. и 57–67 у Trachinostomias.
Помимо перечисленных признаков, возможно, диагностическое значение на уровне групп видов имеет длина V, лучи которых очень длинные у молоди P. margarita (достигают хвостового стебля и часто заходят даже за конец С), тогда как у ювенильных и взрослых экземпляров видов из групп “albipennis” и “caerulescens” V сравнительно короткие, не заходящие своими концами за начало А. К сожалению, у всех остальных изученных представителей рода, даже ювенильных стадий, концы V обломаны, в связи с чем оценить степень его изменчивости между видами не представляется возможным.
Ричардс (Richards, 2006) указывает на различия между личинками подродов Photonectes s. str. и Trachinostomias в дорсальной меланофорной пигментации (соответственно один или три–семь меланофоров на миомер), но конкретные виды им не перечисляются.
Подытоживая сказанное, можно заключить, что ранее высказанное мною мнение о существовании в составе рода Photonectes лишь двух подродов (номинативного и Trachinostomias) (Прокофьев, 2015) полностью подтверждается проведённым здесь анализом, однако признаки, которые должны быть положены в основу выделения этих подродов, следует полностью пересмотреть. Вид P. uncinatus следует переместить из подрода Trachinostomias в номинотипический подрод, в пределах которого он наиболее близок к P. braueri и P. distichodon. Внутри Photonectes s. str. возможно объединение ряда видов в группы, по крайней мере, часть из которых, возможно, монофилетична. Однако эти предположительно монофилетические группы (“albipennis” и “caerulescens”), по-видимому, имеют общих предков с более генерализованными видами Photonectes s. str., в связи с чем на настоящем уровне изученности группы нет причин для более дробного таксономического деления последнего.
Подрод Trachinostomias Parr, 1927
Trachinostomias: Parr, 1927. P. 105 (типовой вид – Echiostoma margarita Goode et Bean, 1896; по последующему обозначению: Eschmeyer, 2018).
Microchirichthys: Regan, Trewavas, 1930. P. 120, 124 (типовой вид – Photonectes parvimanus Regan et Trewavas, 1930, по последующему обозначению: ZR (Pisces), 1930. P. 48, цит. по: Eschmeyer, 2018).
Диагноз. Озубление praemaxillare и dentale гетерогенное у молоди, гомогенное у взрослых рыб; озубление praemaxillare унисериальное на всех стадиях онтогенеза; в дефинитивном состоянии все зубы только с простыми вершинами. Фотофоры IP отстоят друг от друга на равные промежутки, длина которых значительно превышает диаметр фотофора; первая пара фотофоров IP отстоит от вершины истмуса на расстояние, соизмеримое с длиной промежутка между IP-1 и IP-2. BR 9–14. Кожа заходит на D и А, как правило, достигая концов лучей, как у молоди, так и у взрослых рыб. Vert. 57–67.
Состав. См. табл. 1.
Замечания. P. munificus отнесён к этому подроду на основании данных первоописания (Gibbs, 1968), однако он сильно уклоняется от остальных видов по многим признакам, а особенности озубления челюстей и расположения фотофоров IP у него не описаны (но на рисунке голотипа зубы в челюстях изображены мелкими и однородными). Прочие виды данного подрода характеризуются довольно однообразным набором морфологических особенностей, различаясь между собой, главным образом, строением Р и подбородочного усика (рис. 3а–3о). Материал, относимый в настоящее время к P. margarita, характеризуется значительной изменчивостью по этим признакам, позволяющей предполагать его сборный характер. Ювенильным особям этого вида свойственно наличие уплощённой с боков, конической или почти прямоугольной луковицы типа “intermedius” (рис. 3а). Взрослые формы с известной долей условности можно разделить по строению усика на три группы: 1) усик вовсе без луковиц или с придатком позади (проксимальной) луковицы (несущим филаменты второго порядка), столь же развитым, как и ствол (сюда относятся типы всех остальных номинальных видов, помещаемых в синонимию P. margarita) (рис. 3б–3д); 2) усик с терминальной луковицей, любые придатки которой намного тоньше ствола (рис. 3е–3к), и 3) усик без луковицы, но с терминальным коническим расширением, имеющим прозрачное “окно”, внутри которого виден ствол (светящийся?) (рис. 3л). Отсутствие чётких границ между этими группами и других признаков, подтверждающих их выделение, заставляет меня рассматривать их как варианты одного полиморфного вида. Корреляции между типами строения усика и числом лучей Р на моём материале не выявлено.
Подрод Photonectes s. str.
Lucifer: Döderlein, 1882. P. 26 (nom. praeocc., non Thompson, 1830 (Crustacea), nec Reichenbach, 1849 (Aves); типовой вид – Lucifer albipennis Döderlein, 1882, по монотипии).
Photonectes: Günther, 1887. P. 212 (nom. nov. pro Lucifer Döderlein, 1882).
Dolichostomias: Parr, 1927. P. 106, 111 (типовой вид – Photonectes gracilis Goode et Bean, 1896, по последующему обозначению: Eschmeyer, 2018).
Melanonectes: Regan, Trewavas, 1930. P. 119, 120 (типовой вид – Photonectes dinema Regan et Trewavas, 1930, по последующему обозначению: ZR (Pisces), 1930. P. 48, цит. по: Eschmeyer, 2018).
Photonectoides: Koefoed, 1956. P. 11 (типовой вид – Photonectoides paucidentatus Koefoed, 1956, по монотипии).
Диагноз. Озубление praemaxillare и dentale у молоди всегда гетерогенное, у взрослых рыб – гетерогенное или гомогенное; озубление praemaxillare, как правило, бисериальное, редко переходит в унисериальное на поздних стадиях онтогенеза (группа “albipennis”), у видов комплекса “braueri–caerulescens”, возможно, может быть унисериальным на самых ранних онтогенетических стадиях (SL < 35 мм); в дефинитивном состоянии все или часть зубов praemaxillare (иногда и на других костях) обычно с дополнительным зубчиком у вершины, редко все зубы простые (P. gracilis ?). Первая пара фотофоров IP далеко отстоит от вершины истмуса; первые 6–8 фотофоров этой серии сильно сближены между собой, отделены промежутком от последующих (у молоди видов из группы “albipennis” до SL 50–60 мм все фотофоры этой серии разделены равными промежутками). BR 6–10. У молоди кожа покрывает только основания D и А; у взрослых рыб лучи вертикальных плавников от совершенно свободных от кожи (группа “albipennis”) до покрытых ею до самых кончиков (P. uncinatus). Vert. 48–55.
Состав. См. табл. 1.
Замечания. По характеру онтогенетических изменений в признаках озубления челюстей, в степени захождения кожи на вертикальные плавники и в расположении фотофоров в ряду IP виды номинотипического подрода распадаются на две группы – комплекс “braueri–caerulescens” и группу видов “albipennis”. Для комплекса “braueri–caerulescens” характерно гетерогенное озубление praemaxillare и dentale и бисериальное озубление praemaxillare в дефинитивном состоянии; захождение с ростом рыб кожи на лучи вертикальных плавников так, что крупные рыбы становятся не отличимыми по этому признаку от представителей подрода Trachinostomias, и формирование промежутка между передними и задними фотофорами IP уже на ранних стадиях онтогенеза. Видам группы “albipennis” свойственно изменение в ходе онтогенеза гетерогенного озубления praemaxillare и dentale на гомогенное и тенденция к унисериализации премаксиллярного озубления, полностью свободные от кожи вертикальные плавники у взрослых рыб и сохранение сплошного ряда фотофоров IP вплоть до поздних мальковых стадий (SL ~ 50 мм). Кроме того, видам группы “albipennis” свойственно меньшее число фотофоров BR (5–7), чем большинству видов комплекса “braueri–caerulescens” (8–10), но у входящих в состав последнего P. achirus и P. phyllopogon число органов BR также может уменьшаться до 6. К группе “albipennis” принадлежат P. albipennis, P. barnetti, P. coffea и P. waitti.
Различия между видами группы “albipennis” нуждаются в дальнейшем изучении. Различия в строении усика между P. albipennis, P. barnetti и P. coffea скорее количественные, нежели качественные, тогда как другие признаки, предлагавшиеся для разделения этих видов (Klepadlo, 2011), не поддерживаются нашим материалом. В частности, указание на разницу в числе лучей D между P. albipennis и P. barnetti (12–13 против 14–15), возможно, объясняется ошибкой старых описаний первого вида (Döderlein, 1882; Günther, 1887), на которую указывали еще Парин и Соколовский (1976). Во всяком случае, в моём материале все виды группы “albipennis” обычно имеют 14–16 лучей D, их наибольшее число у экземпляров с усиком, типичным для P. albipennis, достигает 18. Различия в длине жаберных лепестков, кажется, хороший видовой признак, но его экогеографическая изменчивость внутри вида также не исключается, во всяком случае, в моём материале есть экземпляр, у которого при типичном для P. albipennis строении усика жаберные лепестки укорочены (рис. 3п), что соответствует P. barnetti по данным Клепадло (Klepadlo, 2011). Напротив, у топотипного экземпляра P. barnetti с банки Милуоки (БМРТ “Академик Берг”, 09.05.1970 г.) жаберные лепестки оказались длинными. Экземпляры со всеми тремя типами усика могут быть встречены в одной и той же водной массе. В изученном материале найден экземпляр, у которого ствол усика на большей части своей длины с резко ослабленной пигментацией (рис. 3р). С другой стороны, учитывая неоднородность изученного материала по P. albipennis в деталях озубления челюстей, длине усика, пигментации его структур, степени развития его дистального филамента (от очень длинного до отсутствующего) и светящейся ткани близ основания терминальной луковицы, нельзя полностью исключать возможную сборную природу этого материала.
Комплекс “braueri–caerulescens” может быть разделён на группы видов “braueri” и “caerulescens”. Группа “braueri” характеризуется только плезиоморфными признаками, а именно наличием Р с двумя небольшими лучами, отсутствием голубой светящейся ткани на теле и передненижнего выступа посторбитального органа (точнее, пигментированной кожи, окаймляющей фотогенную железу). В этой группе виды P. braueri, P. distichodon и P. uncinatus известны по половозрелым рыбам; у них подбородочный усик очень мал. При этом для P. braueri известна также молодь, строение усика которой свидетельствует о том, что у этого вида редукция усика происходит с ростом рыбы. Два других вида данной группы (P. dinema и P. leucospilus) известны только по малькам, имеющим хорошо развитый подбородочный усик. Однако если в ходе последующих исследований удастся выявить у этих видов редукцию усика в процессе развития, то эта особенность может оказаться апоморфией группы “braueri”.
Группа “caerulescens” включает виды, характеризующиеся наличием голубой светящейся ткани на теле, образующей характерный для каждого вида рисунок, и наличием длинного передненижнего выступа чёрной кожи, окружающей светящуюся железу посторбитального органа. Оба этих признака, очевидно, являются апоморфными и свидетельствуют в пользу монофилии данной группы. Помимо этого видам группы “caerulescens” свойственна утрата Р, скорее всего, осуществляющаяся у них независимо от таковой у видов группы “albipennis”. Но, хотя монофилия группы “caerulescens” и выглядит на настоящем уровне изученности наиболее обоснованной, эта группа совершенно очевидно выводится из группы “braueri”, монофилия которой пока не может быть доказанной. Усик у взрослых рыб из группы “caerulescens” развит лучше, чем у таковых в группе “braueri”, однако сведения об онтогенетической изменчивости длины усика у видов группы “caerulescens” отсутствуют в связи с исключительной редкостью рыб. В состав группы “caerulescens” входят P. achirus, P. caerulescens, P. gracilis, P. mirabilis и P. venetaenia.
Кроме того, я отношу к комплексу “braueri–caerulescens” ещё пять номинальных видов, которых нельзя поместить в какую-либо группу. Все они утрачивают Р. Из них P. corynodes и P. litvinovi, по-видимому, близки между собой и сходны с P. mirabilis по строению подбородочного усика, имеющего боковую ветвь, но отличаются от последнего полным отсутствием святящейся ткани на голове, туловище и внутри рта. P. cornutus представляет собой загадочный вид, который, по имеющимся описаниям (Beebe, 1933; Morrow, Gibbs, 1964), полностью подходит под характеристику P. mirabilis, за исключением отсутствия у него полосы голубой светящейся ткани на туловище. Можно предположить, что этот признак был пропущен при описании P. cornutus и таким образом правы Гиббс и Бэрнетт (Gibbs, Barnett, 1990), поместившие этот вид в синонимию P. mirabilis. Однако после описания P. corynodes и P. litvinovi, характеризующихся сходным строением усика при полном отсутствии у них светящейся ткани, в таком предположении нет абсолютной уверенности. Необходимо переисследование голотипов P. cornutus и P. mirabilis, до результатов которого я предлагаю считать P. cornutus species inquirenda. Ещё два вида, P. paxtoni и P. phyllopogon, известны мне только по молоди44, и для уточнения их филогенетического положения необходимо исследование взрослых рыб.
Таким образом, хотя внутри номинотипического подрода удается достаточно уверенно выделить диагностируемые группы видов, чёткой границы между этими группами на настоящем уровне изученности провести нельзя. Поэтому, несмотря на то что группа “braueri” соответствует подроду Melanonectes, для группы “caerulescens” подродовым названием стало бы Dolichostomias (оно же пригодно для обозначения в качестве особого подрода всего комплекса “braueri–caerulescens”), а группа “albipennis” представляет собой номинотипический подрод в наиболее узком его понимании, я не вижу необходимости придавать этим группам таксономический ранг, тем более что при этом часть видов окажется с неясным подродовым статусом. Надо полагать, что детальный сравнительно-анатомический анализ всех видов рода позволит разрешить проблемы взаимоотношений между этими группами и их филогенетический статус, однако в обозримом будущем такой анализ представляется мне недоступным из-за отсутствия возможности детального анатомирования многих видов. На данный момент я предполагаю, что подроды Trachinostomias и Photonectes s. str. в принятом здесь объеме имеют наиболее давнее время обособления и длительную историю независимого развития и нельзя исключать, что более глубокие исследования дадут основания для разделения их в ранге близких родов. Внутри подрода Photonectes s. str. группа видов “albipennis” могла обособиться раньше группы “caerulescens”, но обе эти группы на настоящем уровне изученности представляются кроновыми филетическими линиями, ведущими своё происхождение от форм, близких к видам группы “braueri” или к P. paxtoni, характеризующихся наибольшим числом примитивных особенностей.
ОПИСАНИЯ НОВЫХ ВИДОВ
(рис. 2м, 4, 5)
Материал. Голотип, ИО РАН № 3584, SL 167 мм, 35°33ʹ6ʹʹ ю.ш., 116°31ʹ6ʹʹ в.д., НИС “Дмитрий Менделеев”, рейс 16, ст. 1378, 02.03.1976 г., РТАК № 49, горизонт лова 200–0 м, время 02.30–03.30. Паратип, ИО РАН № 3585, SL 175 мм, 34°43ʹ2ʹʹ ю.ш., 123°53ʹ7ʹʹ в.д., НИС “Дмитрий Менделеев”, рейс 16, ст. 1374, 28.02.1976 г., орудие лова ОРН-18, горизонт лова 1825–0 м, на горизонте 13.40–14.40, окончание траления 17.30.
Диагноз. Вид рода Photonectes с двумя лучами Р, бисериальным типом озубления praemaxillare, 4 (5?) зубами в наружном ряду на praemaxillare, с 51 позвонком, без голубой светящейся ткани на теле, с крошечным подбородочным усиком у взрослых рыб (1.4–2.4% SL при SL 167–175 мм), имеет чёткую терминальную луковицу, не выступающую за границы ствола и несущую интенсивно пигментированный терминальный филамент.
Описание. D 18–19, A 20–21, P 2, V 7; vert. 51 (хвостовых 13–14). Результаты некоторых измерений представлены в табл. 2. Фотофоров в сериях BR 9–10, IP 8 + 2, PV 24–25, OV 21–са.23, VAL около 9(?), точное число органов в сериях VAV и АС подсчитать нельзя; до ануса 9 органов VAV, далее назад кожа большей частью содрана. Органы BR расположены на равном расстоянии или первые три сближены, последний всегда сильно уменьшен. Первые восемь органов IP сближены, отделены промежутком от двух последних, более расставленных; первая пара органов отстоит от вершины истмуса на промежуток, равный длине ряда из 6.5 передних фотофоров IP. Посторбитальный орган (рис. 4а) почти конической формы, сужающийся назад, на его передний край слегка налегает узкая кожная складка. Максимальная высота посторбитального органа втрое меньше длины. Установить наличие или отсутствие пятен светящейся ткани на рыле и за жаберным отверстием не представляется возможным из-за повреждения кожи. Светящихся органов дна ротовой полости нет. Лучи Р довольно слабые, у голотипа сохранились почти полностью, из них нижний хуже развит, длина верхнего луча соизмерима с длиной промежутка из пяти противолежащих фотофоров PV. У паратипа сохранились только основания лучей Р равной толщины. Кожа в области вертикальных плавников сорвана.
Таблица 2.
Морфометрические признаки трёх видов Photonectes
| Признак | P. distichodon sp. n. | P. filipendulus sp. n. | P. xenopogon sp. n. | ||||
|---|---|---|---|---|---|---|---|
| голотип | паратип | голотип, самка | паратипы, самцы | нетиповой экземпляр | голотип | паратип | |
| SL, мм | 167 | 175 | 235 | 138–190 | 155 | 104 | 168 |
| В % SL | |||||||
| Максимальная высота тела | 12.0 | 10.9 | 10.6 | 10.9–13.1 | 12.9 | 11.5 | 10.1 |
| Минимальная высота тела | 1.8 | 1.7 | 1.0 | 1.0–2.0 | 2.6 | 1.9 | 2.1 |
| Предорсальное расстояние | 82.6 | 82.9 | 84.3 | 83.3–85.0 | 84.5 | 85.6 | 83.9 |
| Преанальное расстояние | 79.6 | 81.7 | 84.3 | 82.6–85.0 | 84.5 | 83.7 | 82.1 |
| Превентральное расстояние | 66.2 | 66.3 | 72.3 | 68.4–70.6 | 69.0 | 66.4 | 66.1 |
| Длина головы | 18.0 | 14.3 | 13.2 | 15.0–16.1 | 14.8 | 14.4 | 14.3 |
| Длина рыла | 1.8 | 1.7 | 2.1 | 1.7–2.2 | 2.3 | 2.4 | 2.1 |
| Горизонтальный диаметр глаза | 3.0 | 2.9 | 1.9 | 2.4–2.9 | 2.6 | 2.9 | 2.7 |
| Ширина межглазничного промежутка | 4.2 | 4.3 | 4.3 | 3.9–4.6 | 4.5 | 3.9 | 4.2 |
| Длина верхней челюсти | 12.0 | повреждена | 11.5 | 12.1–13.7 | 12.9 | 12.5 | 11.9 |
| Длина усика | 2.4 | 1.4 | 3.0 | 3.5–9.2 | луковица без ствола | 4.8 | повреждён |
| Длина посторбитального органа | 3.6 | 3.4 | 2.6 | 2.9–3.6 | 3.2 | 1.0 | 2.4 |
Рыло короче глаза. Озубление praemaxillare бисериального типа, зубы на maxillare и dentale разноразмерные (рис. 4а, 4б). Зубов praemaxillare 9–11 + 4(5?), в наружном ряду две пары зубов разделены широким промежутком (у голотипа на одной praemaxillare за последним зубом имеется недоокостеневшая закладка 5-го зуба, но неясно, замещает ли она 4-й зуб или формирует отдельный зуб). Наибольшие зубы praemaxillare в полтора раза длиннее диаметра зрачка и во столько же превышают длину наибольших зубов maxillare и dentale, последние соизмеримы по величине. На maxillare 11–14 расставленных зубов, за которыми расположена гребёнка из мелких частых зубов, до семи передних зубов наиболее крупные, постепенно увеличивающиеся в длине от 1-го к 7-му, самые задние зубы значительно меньшей величины.
На dentale 28–32 зуба, резко различающихся по величине, наиболее длинные зубы перемежаются несколькими более короткими. На сошнике 3 + 3 (голотип) или 2 + 2 длинных зуба с каждой стороны образуют косой ряд, в котором зубы прогрессивно удлиняются от наружного к внутреннему. На palatinum четыре–шесть разноразмерных и неравно расставленных зуба. Первая жаберная дуга несёт сильные зубчики, в большинстве своём собранные в группы по два–три; жаберные лепестки в верхней половине ceratobranchiale-1 укороченные. Подбородочный усик очень мал, немного короче рыла, его ствол интенсивно пигментирован. Терминальная луковица хорошо выражена, но совершенно не выступает за границы ствола, на вершине несёт единственный сильно пигментированный филамент, у голотипа составляющий примерно половину длины луковицы, а у паратипа – равный ей (рис. 4в, 4г).
Этимология. Видовой эпитет образован от греческих слов “distichus” (двурядный) и “odus” (зуб) по характерному признаку озубления praemaxillare.
Сравнительные замечания. Новый вид очень сходен с P. braueri (Zugmayer, 1913), от которого отличается наличием хорошо развитого пигментированного терминального филамента луковицы усика у взрослых рыб и бóльшим числом зубов в наружном ряду на praemaxillare (4, возможно 5, против 1–2).
Результаты изучения онтогенетической изменчивости строения усика у P. braueri в целом совпадают с данными предшествующих авторов (Morrow, Gibbs, 1964; Gibbs, 1984), но нужно отметить, что наименьший из имеющихся в моём распоряжении экземпляров (SL 22 мм, НИС “Витязь”, ст. 5028) имеет помимо маленького шаровидного терминального придатка луковицы, характерного для молоди этого вида SL 22–31 мм (Regan, Trewavas, 1930; Gibbs, 1984), также крупный плоский вытянутый почти не пигментированный придаток (неплотное скопление меланофоров присутствует только в его дистальной половине) с двумя непигментированными филаментами в основании (рис. 4д). Наличие такого придатка у молоди P. braueri ранее не отмечалось. Он весьма напоминает соответствующий придаток у P. parvimanus Regan et Trewavas, 1930, но у последнего никогда не бывает терминального шаровидного придатка луковицы. К сожалению, этот малёк находится в очень плохой сохранности, так что большинство других признаков, пригодных для различения этих видов, у него установить невозможно (в частности, неясно озубление praemaxillaria), однако его вертикальные плавники совершенно не покрыты кожей, что позволяет определить его как P. braueri. Возможно, наличие подобного придатка свойственно особям самых ранних стадий онтогенеза и в последующем он утрачивается, однако более мелкие особи P. braueri до сих пор не известны. У рыб SL 50 мм (НИС “Петр Лебедев”, IV–123A; НИС “Витязь”, ст. 4577) имеется крупная шаровидная луковица без придатков (рис. 4е), терминальный шаровидный придаток, описанный для молоди SL 22–31 мм (Regan, Trewavas, 1930), у них утрачен, но имеется у экз. SL 43 мм, НИС “Академик Курчатов”, ст. 835: рис. 4ж). Наибольший из имеющихся в моём распоряжении экземпляров P. braueri (SL 225 мм, НИС “Эврика”, Угловое поднятие, банка Сложная) совершенно утрачивает луковицу, но вершина усика у него не пигментирована и имеет крючковатую форму (рис. 4з). Таким образом, на всех стадиях онтогенеза придатки луковицы у P. braueri имеют иную форму и никогда не бывают интенсивно пигментированными, как у описываемого нового вида.
Ранее P. braueri уже указывался для Юго-Западной Пацифики (Paxton et al., 1989; Paulin et al., 1989), но морфологического описания этого материала проведено не было, в связи с чем весьма вероятно, что под этим названием мог указываться P. distichodon. В моих материалах экземпляры P. braueri имеются из Атлантического (между 38° с.ш. и 27° ю.ш.), Индийского океанов и внутренних морей Индо-Австралийского архипелага (07°45ʹ с.ш., 120°25ʹ в.д.); для последних этот вид отмечается впервые.
От другого близкого вида, P. uncinatus, вероятно, также связанного с водами южной субтропической конвергенции, новый вид хорошо отличается относительно небольшим, почти коническим посторбитальным органом (3.4–3.6% SL против 5.8% SL, гантелевидной формы), наличием терминальной луковицы и филамента усика, а также бóльшим числом нёбных зубов (4–6 против 2).
Наличие длинного терминального филамента луковицы характерно для P. dinema Regan et Trewavas, 1930 и P. leucospilus Regan et Trewavas, 1930, но у них он не пигментирован и имеет иное строение (Regan, Trewavas, 1930. Fig. 115A, B ).
(рис. 2о, 4, 5)
Photonectes cf. margarita: Прокофьев, 2015. С. 134, 135. Рис. 2.
Материал. Голотип, ИО РАН № 3586, SL 235 мм, Тихий океан, Гавайский район, 31° с.ш., 174° в.д., НПС “Радуга”, ст. 16. Паратипы (4 экз.): ИО РАН № 3587, SL 174 мм, Тихий океан, 40°19′5ʹʹ с.ш., 135°49′5ʹʹ з.д., НИС “Витязь”, ст. 4191, проба 198, трал Айзекса–Кидда, глубина места 4460 м, горизонт лова 302–326 м, трос 900–1085 м, 08.12.1958 г., время 23.25–00.00; ИО РАН № 3588, SL 138 мм, 29°58′ с.ш., 120°43′ з.д., НИС “Витязь”, ст. 4217, проба 246, трал Айзекса–Кидда, глубина места 4090 м, горизонт лова ~240 м, 26.12.1958 г., время 02.50–03.08; ИО РАН № 3589, SL 190 мм, 25° с.ш., 165° в.д., горизонт лова 100 м, 30.06.1966 г.; ИО РАН № 3590, SL 153 мм, 40°19′5ʹʹ с.ш., 135°49′5ʹʹ з.д., НИС “Витязь”, рейс 29, ст. 4191, проба 197, трал Айзекса–Кидда, трос 1060–810 м, 08.12.1958 г., время 22.25–22.45.
Дополнительный нетиповой материал: 1 экз. SL 155 мм, пойман вместе с паратипом № 3590.
Диагноз. Вид рода Photonectes с одним очень маленьким и слабым лучом Р (иногда утрачен?), унисериальным типом озубления praemaxillare, с 59–62 позвонками, с посторбитальным органом, составляющим 2.9–3.6% (самцы) или 2.6% (самки) SL, с коротким или умеренно длинным подбородочным усиком (3.0–9.2% SL при SL 138–235 мм), имеющим маленькую, не выступающую за границы ствола терминальную луковицу, от вершины которой отходят два–четыре одинаково мелких пигментированных филамента.
Описание. D 16–19, A 18–22, P 0?–1, V 7; vert. 59–62 (хвостовых 14). Некоторые измерения приведены в табл. 2. Фотофоров в сериях BR 11–12, IP 10, PV 35, VAV ca.12 (до ануса – 9); точное число органов в сериях OV, VAL и АС подсчитать нельзя из-за срыва кожи. Органы BR расположены на равном расстоянии или самые передние и/или самые задние органы сближены, все – одинаково мелкие. Все органы IP расположены через одинаковые промежутки; первая пара органов отстоит от вершины истмуса на расстояние, соизмеримое с длиной промежутка между IP-1 и IP-2. Посторбитальный орган (рис. 4и) овально-конической формы, слабо сужающийся назад; складка кожи, прикрывающая его передний край, слабо выражена. Установить наличие или отсутствие пятен светящейся ткани на рыле и за жаберным отверстием не представляется возможным из-за повреждения кожи. Светящихся органов дна ротовой полости нет. Луч Р очень слабый, короткий, мягкий, его длина много меньше длины головы (рис. 4к). У паратипов № 3587 и 3589 никаких следов Р обнаружить не удалось, но нельзя исключать, что он был искусственно утрачен, так как кожа в этой области у рыб сильно повреждена. Кожа заходит на вертикальные плавники.
Рыло незначительно короче, равно или слегка превышает диаметр глаза. Озубление praemaxillare унисериального типа, зубы в челюстях однородные, на maxillare и dentale более мелкие в задних частях соответствующих костей (рис. 4и). Зубов praemaxillare 5–6, наибольшие из них не более чем в полтора раза превышают длину наибольших зубов maxillare и dentale, последние соизмеримы по величине или зубы несколько мельче на dentale. На maxillare 9–16 расставленных зубов, за которыми расположена гребёнка из мелких частых зубов; на dentale зубов 20–32; на сошнике 1 + 1 или 2(3) + 2, на palatinum по 4–7 разноразмерных зубов с каждой стороны. Первая жаберная дуга несёт сильные длинные зубчики; жаберные лепестки по всей длине ceratobranchiale-1 длинные. Подбородочный усик в длину составляет от трети до половины длины головы или несколько больше, с очень маленькой терминальной луковицей, не выступающей за границы ствола, иногда пигментированной снизу, несущей на вершине от двух до четырёх коротких, как правило, одинаковых (у паратипов № 3588 и 3589 один из филаментов короче остальных, но такой же толщины в основании) пигментированных филаментов (рис. 4л–4о). У паратипа № 3587 усик, по-видимому, повреждён или регенерирует, филаментов луковицы не обнаружено. У нетипового экземпляра SL 155 мм в области прикрепления подбородочного усика найдено маленькое округлое, вероятно, светящееся образование, практически без ствола (рис. 4п–4с).
Этимология. Название вида образовано от латинских слов “filis” (нить) и “pendulus” (обвисший) и связано с одним из отличительных признаков вида – наличием у него слабого луча Р (в отличие от жёсткого луча у P. margarita).
Сравнительные замечания. Новый вид чрезвычайно близок к P. margarita (Goode et Bean, 1895), но отличается от последнего слабо развитым (иногда совсем отсутствующим?) лучом Р, строением терминальных филаментов усика и более крупным посторбитальным органом у обоих полов (2.6% у самок и 2.9–3.6% у самцов против соответственно 0.6–1.5 и 1.6–2.8%, как правило, не более 2.5% у P. margarita). У P. margarita всегда имеется один длинный луч Р, по-видимому, значительно превышающий длину головы (изредка присутствует второй, вероятно, гораздо более короткий). У коллекционных экземпляров этот луч, как правило, в той или иной степени обломан, но как раз по этой особенности они и могут быть отделены, так как у нового вида луч Р настолько слаб и мал, что сохраняется практически целиком (кроме того, у P. margarita луч Р в основании всегда намного толще, чем у нового вида, что хорошо заметно при прямом сравнении экземпляров). У двух паратипов (№ 3587 и 3589) Р не найден, но эта область у них сильно повреждена, так что нельзя исключать, что Р здесь искусственно утрачен. В то же время необходимо отметить, что в других случаях Р может сохраняться при полном срыве кожи, так что нельзя исключать и его полной редукции у части экземпляров. У P. margarita никогда не наблюдается редукции луча Р.
Подбородочный усик у нового вида характеризуется маленькой луковицей, несущей от двух до четырёх пигментированных филаментов, либо одинаково развитых, либо один из филаментов короче остальных (рис. 4л–4о). Эта особенность хорошо отличает новый вид от P. margarita, у которого при всём разнообразии вариантов строения усика всегда имеется один крупный филамент, с которым (либо с терминальным концом луковицы, хотя иногда луковица может вовсе отсутствовать) связано различное число филаментов более низкого порядка. Таким образом, можно сказать, что у P. filipendulus все терминальные филаменты одного порядка, тогда как у P. margarita имеется один крупный филамент первого порядка и многочисленные филаменты второго порядка (рис. 3б–3к). Экземпляры, которые можно было бы счесть промежуточными, не известны.
Перечисленные особенности уже были отмечены ранее (Прокофьев, 2015), но тогда уклоняющиеся экземпляры были обозначены как P. cf. margarita. Однако нахождение дополнительных материалов подтверждает стабильность данных отличий, в связи с чем я считаю необходимым выделить эту форму в самостоятельный вид.
Особняком стоит экземпляр SL 155 мм (НИС “Витязь”, ст. 4191), который не включён в типовую серию вида. Он довольно сильно повреждён, но имеет один маленький слабо развитый луч Р, из известных видов фотонектов характерный только для Р. filipendulus, вместе с одним из паратипов которого он был пойман. Однако в области прикрепления подбородочного усика у этого экземпляра найдено маленькое округлое, вероятно, светящееся образование, практически без ствола (рис. 4п–4с). Неясно, является ли это аберрацией, связанной с регенерацией оторванного усика. Один из паратипов Р. filipendulus (№ 3587) имеет короткий усик с маленькой терминальной луковицей без филаментов, предположительно регенерировавшей. Но ствол у него вполне развит. Для уточнения ситуации, наблюдаемой у экземпляра SL 155 мм, необходимы новые материалы.
(рис. 2н, 5, 6а–6г)
Photonectes sp. 1: Парин и др., 1977. С. 95 (part.: только ст. 7243).
Photonectes (Melanonectes) dinema: Прокофьев, 2014. С. 382 (ошибочное определение).
Материал. Голотип, ИО РАН № 3591, SL 104 мм, 08°40′ с.ш., 119°48′ в.д., НИС “Витязь”, рейс 57, ст. 7243, РТАК, горизонт лова 500 м, 01–02.03.1975 г. Паратип, ИО РАН № 3592, SL 168 мм, пойман вместе с голотипом.
Диагноз. Вид рода Photonectes с одним длинным и крепким лучом Р, состоящим из двух плотно соединённых половинок, унисериальным типом озубления praemaxillare, с 59–60 позвонками, с посторбитальным органом, составляющим 1.0% (неполовозрелый самец) или 2.4% (половозрелый самец) SL, со сравнительно коротким подбородочным усиком (4.8% SL при SL 110 мм), имеющим удлинённую, не выступающую за границы ствола терминальную луковицу со внутренней пигментацией, несущую плоский, непигментированный, асимметрично раздвоенный на вершине терминальный филамент.
Описание. D 18, A 23, P 1, V 7; vert. 59–60 (хвостовых 13). Некоторые измерения см. в табл. 2. Фотофоров в сериях BR 9, IP 10, VAV са.12; в остальных сериях точное число органов подсчитать нельзя из-за повреждения кожи. Органы BR одинаково мелкие и расположены на равном расстоянии друг от друга. Все органы IP расставлены на одинаковые промежутки; первая пара органов отстоит от вершины истмуса на расстояние, соизмеримое с длиной промежутка между IP-1 и IP-2. У обоих экземпляров имеется суборбитальный орган, но у голотипа он заметно крупнее; напротив, посторбитальный орган гораздо меньше, чем у паратипа55 (рис. 6а, 6б). Посторбитальный орган удлинённо-овальный, несколько сужающийся назад. Установить наличие или отсутствие пятен светящейся ткани на голове и за жаберным отверстием не представляется возможным из-за повреждения кожи. Светящихся органов дна ротовой полости нет. Луч Р длинный и крепкий, состоит из двух плотно соединённых половинок, которые при отламывании конца луча могут расходиться, создавая иллюзию двух лучей66. Кожа в области вертикальных плавников большей частью сорвана, но у паратипа перед А имеется лоскут кожи, который, будучи расправленным, не оставляет сомнений в том, что он целиком покрывал переднюю часть этого плавника.
Рис. 6.
Детали строения Photonectes spp.: а–г – P. xenopogon sp. n. (а, б – посторбитальный орган (а – голотип, б – паратип); в – озубление челюстей, голотип; г – подбородочный усик, голотип); д – P. margarita ЗММУ № 23289, подбородочный усик; е–з – Photonectes sp. indet. (е – вид головы сверху и praemaxillaria, стрелкой показано пятно белой светящейся ткани кнутри от глаза; ж – нижняя челюсть и подбородочный усик; з – луковица усика); и – P. phyllopogon, НИС “Сергей Вавилов”, ст. 2669, подбородочный усик. Масштаб: а–в, д–ж – 3 мм, г – 1 мм; з, и – 0.5 мм.
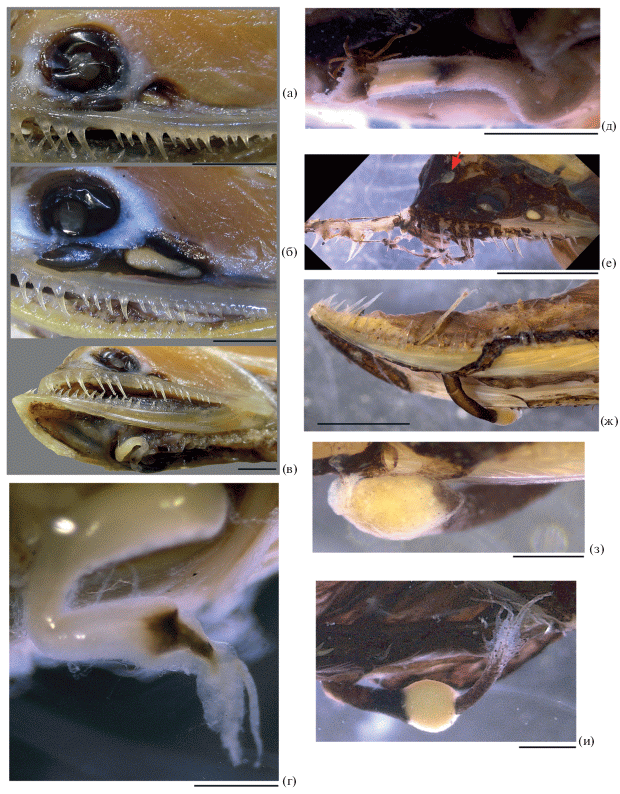
Рыло короче глаза. Озубление praemaxillare унисериального типа, зубы в челюстях однородные, на maxillare и dentale уменьшаются по величине к задним концам соответствующих костей (рис. 6в). Зубов praemaxillare 6, у голотипа имеется небольшой промежуток между тремя передними и тремя задними зубами, у паратипа два передних зуба сильнее отстоят от последних четырёх; самые передние зубы короче последующих. Наибольшие зубы praemaxillare почти вдвое превышают диаметр зрачка и примерно в полтора раза больше наибольших зубов maxillare и dentale, последние соизмеримы по величине. На maxillare 13–18 расставленных зубов, за которыми расположена гребёнка из мелких частых зубов; на dentale 27–30 зубов; у голотипа на сошнике 3 + 3, у паратипа 1 + 1 зуб; на palatinum по 5 зубов с каждой стороны. Первая жаберная дуга несёт многочисленные сильные зубчики; жаберные лепестки по всей длине ceratobranchiale-1 длинные. Подбородочный усик составляет около трети длины головы, его ствол практически не пигментирован (возможно, верхний слой эпидермиса стерт). Терминальная луковица удлинённая, не выступает за границы ствола, с интенсивной внутренней пигментацией характерной формы (рис. 6г), на вершине несёт единственный плоский непигментированный раздваивающийся на конце филамент, длина которого соизмерима с длиной луковицы.
Этимология. Видовой эпитет образован от греческих слов “xenos” (чуждый) и “pogon” (борода), даётся в связи с тем, что по строению усика новый вид похож не на родственных ему представителей подрода Trachinostomias, а на далекий по другим признакам вид P. dinema.
Сравнительные замечания. Типовые экземпляры описываемого вида ранее были отнесены мною к P. dinema на основании сходного строения подбородочного усика (Regan, Trewavas, 1930. Fig. 115B ), отличающегося лишь наличием у нового вида внутренней пигментации луковицы, для P. dinema не отмеченной, и асимметричной формой терминального филамента. Однако по другим признакам (унисериальное против бисериального озубление praemaxillare, однородные против разноразмерных зубы maxillare и dentale, гораздо большее число позвонков (59–60 против 50), отсутствие промежутка между передними и задними фотофорами IP) новый вид резко отличается от P. dinema, но близок P. margarita, P. filipendulus и P. parvimanus, усики которых при всей их изменчивости существенно различаются по строению (рис. 3а–3о, 4л–4о). Наличие внутренней пигментации луковицы усика является уникальным признаком нового вида в пределах рода и исключает сомнения в его самостоятельности.
Парин с соавторами (1977) отнесли типовые экземпляры нового вида вместе с тремя другими пойманными в 57-м рейсе НИС “Витязь” (станции 7192 и 7374) к обозначенному ими в открытой номенклатуре Photonectes sp. 1, по моему мнению, основанному на сборном материале. Рыбы со станций 7192 и 7374 (ЗММУ № 23286 и 23289) резко отличаются от P. xenopogon отсутствием внутренней пигментации усика и наличием множественных пигментированных нитевидных придатков у её передненижнего угла (против плоского непигментированного терминального придатка у P. xenopogon). Хотя наличие “пигментированной оси” указывается Париным с соавторами для всех рыб, в действительности таковая отсутствует у рыб со станций 7192 и 7374, а имеется только наружная пигментация (рис. 6д). В моём материале имеется ещё несколько экземпляров таких рыб (ЗММУ № 23287, 23295, 23299), характеризующихся усиком, лишённым нормально развитой луковицы, но с прозрачным “окном” на дистальном расширенном конце, внутри которого виден тяж, возможно, светящейся ткани; спереди и сзади это “окно” ограничено наружной пигментацией, у некоторых рыб эти участки пигмента могут быть соединены широкой полосой также наружного пигмента, в других случаях не выраженной. Усик несёт группу пигментированных дистальных нитевидных придатков. Я отношу эти экземпляры к P. margarita, хотя более представительные материалы, возможно, дадут основания для их таксономического обособления от рыб, имеющих нормальную терминальную луковицу или вовсе без таковой. Однако они определенно не конспецифичны P. xenopogon.
(рис. 6е–6з)
Материал. ИО РАН, SL 52 мм, 31°02ʹ с.ш., 150°31ʹ в.д., НИС “Экватор”, ст. 251, 14.11.1968 г., разноглубинный трал 31 м, длина ваеров 350 м, скорость 3.8 узла, коллектор Комраков.
Замечания. Этот малёк найден в пробе с 4 экз. P. albipennis и, по-видимому, принадлежит к неописанному виду, характеризующемуся резко гетерогенным озублением praemaxillare и dentale, но унисериальными зубами на praemaxillare (всего их около 8) (рис. 6е, 6ж); наличием дополнительных зубчиков у вершин премаксиллярных зубов; коротким подбородочным усиком с крупной луковицей, несущей небольшой плоский лопастевидный придаток (рис. 6ж, 6з); наличием пятен белой светящейся ткани над глазом и над жаберным отверстием (рис. 6е); BR 12; укороченными жаберными лепестками в верхней части ceratobranchiale-1; D и А, не покрытыми кожей. Область Р сильно повреждена, как и истмус, но первая пара органов IP, по-видимому, отстоит от вершины истмуса на расстояние двух промежутков между IP-1 и IP-2 или чуть более. Подродовая принадлежность этого малька не вполне ясна. От видов номинотипического подрода он отличается увеличенным числом органов BR. По строению подбородочного усика он соответствует ранним малькам P. parvimanus, но полностью не покрытые кожей вертикальные плавники при SL 52 мм и наличие пятна белой светящейся ткани над глазом исключают отнесение его к этому виду. Последний признак, по-видимому, уникален для нового вида. Среди других фотонектов пятна белой светящейся ткани на голове имеются у P. dinema, P. leucospilus и некоторых видов из группы “caerulescens”, но последние должны быть исключены из сравнения, так как имеют голубую светящуюся ткань на теле, отсутствующую у рассматриваемого вида. У P. dinema и P. leucospilus пара пятен белой светящейся ткани сближена и расположена по средней линии рыла у его вершины (против расставленных пятен в межглазничном промежутке); кроме того, у этих видов отсутствуют пятна светящейся ткани над жаберным отверстием, не более 10 органов BR и совершенно иное строение придатка луковицы усика. Плохая сохранность истмуса не позволяет полноценно оценить строение IP-серии фотофоров. Из-за невозможности провести полноценное сравнение единственного известного экземпляра нового вида с другими видами рода он оставлен пока без видового названия.
КЛЮЧ ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ РОДА PHOTONECTES
(P. cornutus и P. sp. indet. не включены в определительную таблицу из-за недостатка данных)
1(2) Озубление praemaxillare и dentale гетерогенное у молоди и гомогенное у взрослых рыб, praemaxillare – унисериальное на всех стадиях онтогенеза; BR 9–14; первая пара фотофоров IP отстоит от вершины истмуса на расстояние, соизмеримое с длиной промежутка между IP-1 и IP-2; все органы IP отстоят друг от друга на равные промежутки; кожа покрывает более половины длины лучей D и А77 ….………………………….............3
2(1) Озубление praemaxillare гетерогенное бисериальное, dentale – гетерогенное; если озубление челюстей гомогенное, а praemaxillare – унисериальное, то фотофоров BR 5–7; расстояние от IP-1 до вершины истмуса намного превышает длину промежутка между IP-1 и IP-2; передние 6–8 фотофоров IP более сближены и отделены промежутком от последующих, если же этот промежуток не выражен (бывает только у молоди!), то кожа заходит лишь на основания лучей D и А ………………………………………………….12
3(4) IV 49–50, OV 38, VAL 2–4, органы VAV разбиты на три группы; vert. 67; Р 0 ……… P. munificus
4(3) IV не более 45, OV менее 32, VAL 11–14, все органы VAV отстоят на равные промежутки; vert. 57–63; Р (0?)1–2(3) ……………………………………… 5
5(6) Луковица усика с внутренней пигментацией, несёт плоский терминальный филамент, раздвоенный на конце (рис. 6г) …………… P. xenopogon
6(5) Луковица усика, если имеется, без внутренней пигментации; ее придатки, если имеются, иной формы (рис. 3а–3о, 4л–4о) …………………… 7
7(8) Р 1–2(3), если один, то луч длинный и крепкий; если имеется луковица усика с терминальными придатками, то последние многочисленные и/или неодинаковой величины и формы (как на рис. 3а–3о) ……………………...................... 9
8(7) Р 1, крошечный, гибкий, возможно, у некоторых рыб совсем утрачен; луковица усика маленькая, с двумя–четырьмя короткими нитевидными терминальными филаментами ………..P. filipendulus
9(10,11) Vert. 57; IV 40; P 2, лучи гибкие и тонкие, но длинные (равновеликие?); усик с маленькой терминальной луковицей без придатков …………………………………………… ..P. gorodinskii
10(9,11) Vert. 58–63; IV 43–48; P 2(3), лучи гибкие, равновеликие (очень редко встречающийся 3-й луч всегда зачаточный); усик с крупной шаровидной терминальной луковицей, снабжённой плоским субтерминальным придатком, как правило (кроме самых мелких рыб), длинным и несущим в основании пигментированные филаменты второго порядка (рис. 3м–3о) ……… P. parvimanus
11(9,10) Vert. (58)60–63; IV 42–45; P 1, длинный и крепкий, редко два (в этом случае второй луч развит гораздо слабее); терминальная луковица усика, если имеется, маленькая округлая либо уплощённая конической формы, несёт нитевидные (иногда в дополнение к ним и более толстые цилиндрические) филаменты (рис. 3а–3л) …………. P. margarita
12(13) Р 2 ………………………………….................14
13(12) Р 0 ……………………………………….......... 23
14(15) Луковица усика с удлинённым терминальным придатком; на вершине рыла имеется пара округлых пятен белой светящейся ткани [зубов palatinum 1–3] ……………………………........... 16
15(14) Луковица усика с крошечным придатком или без него (у взрослых рыб может быть полностью редуцирована); нет пятен белой светящейся ткани на рыле (не известно для P. distichodon) ……………………....................................... 19
16(17) Тело более высокое, его максимальная высота не более чем в 1.2 раза меньше расстояния от заднего края глаза до нижнечелюстного сустава при SL ~ 24–28 мм; луковица усика сильно вытянута в длину, её терминальный придаток на вершине раздвоен; жаберные лепестки длинные …………………………………………... P. dinema
17(16) Тело более прогонистое, его максимальная высота в 1.4 раза меньше расстояния от заднего края глаза до нижнечелюстного сустава при SL ~ 25 мм; луковица усика овальная или почти шаровидная, её терминальный придаток оканчивается маленькой луковичкой; жаберные лепестки укороченные ………………….. P. leucospilus
19(20) Посторбитальный орган очень длинный (5.8% SL у самки), гантелевидной формы; зубов palatinum 2 …………………………………..... P. uncinatus
20(19) Посторбитальный орган небольшой (3.3–3.6% SL), по форме близок к коническому или треугольному; зубов palatinum 4–6 ………… 21
21(22) В наружном ряду на praemaxillare один–три зуба (1 + 1 или 1 + 2); подбородочный усик у взрослых рыб с полностью редуцированной луковицей и придатками ……………………….... P. braueri
22(21) В наружном ряду на praemaxillare четыре зуба (2 + 2); подбородочный усик у взрослых рыб с маленькой, но хорошо выраженной луковицей, несущей интенсивно пигментированный терминальный филамент ……………….......... P. distichodon
23(24) Голубая светящаяся ткань на теле имеется …………………………………………………………... 25
24(23) Голубая светящаяся ткань на теле отсутствует ………………………………………………………... 33
25(26) На боках тела от жаберного отверстия до основания С проходит срединная полоса голубой светящейся ткани; на брюхе голубая светящаяся ткань представлена продольным рядом пятнышек непосредственно над органами PV, соединяющимся с таким же рядом на противоположной стороне тела поперечными рядами пятнышек, расположенными между органами PV; превентральное расстояние менее 57% SL ………………………… P. gracilis
25(26) Посредине боков тела нет полосы голубой светящейся ткани; на брюхе голубая светящаяся ткань расположена иначе; превентральное расстояние более 60% SL …………………............. 27
27(28) Подбородочный усик с маленькой луковичкой в основании и крупной луковицей, несущей длинный терминальный придаток с дополнительными луковичками и коротким боковым отростком; имеется три пары пятен светящейся ткани в дне рта; пятно белой(?) светящейся ткани перед 1-м фотофором BR; полоска голубой светящейся ткани на истмусе с каждой стороны снутри от органов IP, широко разобщённая с продольным рядом пятнышек голубой ткани (могут сливаться в сплошную полосу?) между рядами фотофоров OV и PV вдвое ближе к PV …..................................................P. mirabilis
28(27) Подбородочный усик с одной крупной луковицей или без таковой, с толстым пигментированным стволом и тонким непигментированным придатком, утолщённым на конце; нет пятен светящейся ткани в дне рта и перед фотофорами BR (кроме P. achirus); расположение скоплений голубой ткани иное ……………………….................29
29(30) Голубая светящаяся ткань образует две параллельные полосы вдоль срединной линии на истмусе, на уровне 1–2-го фотофоров IP переходящих на боковые стороны тела снаружи от IP, и дополнительную нерезкую полоску над самыми передними фотофорами PV [BR 9; подбородочный усик без луковицы] ……………………….. P. venetaenia
30(29) Голубая светящаяся ткань образует полосу по срединной линии брюха между соседними рядами фотофоров PV, отдающую ответвления наружу между фотофорами PV, и продольным рядом пятнышек непосредственно над органами PV (его наличие не указано для P. achirus, но для него указано скопление под OV-1–2: Flynn, Klepadlo, 2012. Р. 266)………………………….31
31(32) Подбородочный усик с развитой луковицей; одна пара пятен светящейся ткани в дне рта; есть светящаяся ткань перед BR-1; BR 6–8……………………………………………....P. achirus
32(31) Подбородочный усик без луковицы; нет светящейся ткани в дне рта и перед BR-1; BR 8–10 ………………………………........P. caerulescens
33(34) Подбородочный усик несёт боковой отросток ……………………………………………………….. 35
34(33) Подбородочный усик без бокового отростка ……………………………………………………..... 37
35(36) Подбородочный усик без луковицы; длина рыла втрое короче диаметра глаза; зубов dentale 29; А 19 ………………………………….. P. litvinovi
36(35) Подбородочный усик с крупной луковицей; длина рыла соизмерима с диаметром глаза; зубов dentale 16; А 16 …………………… P. corynodes
37(38) Подбородочный усик короткий, не более половины длины нижней челюсти, с очень крупной овальной или шаровидной луковицей без придатков или с листовидным светящимся придатком; нет пятен белой светящейся ткани над жаберным отверстием …………………………... 39
38(37) Подбородочный усик обычно длиннее нижней челюсти, а если короткий, то несёт две последовательно расположенные луковицы; терминальная луковица усика с нитевидным филаментом; одно–два пятна белой светящейся ткани над жаберным отверстием …………………............ 41
39(40) Подбородочный усик с крупной овальной луковицей без придатков; VAV 15–16 ………P. paxtoni
39(40) Подбородочный усик с крупной шаровидной луковицей, несущей интенсивно пигментированный терминальный филамент, оканчивающийся непигментированным (светящимся) листовидным придатком с перистым дистальным краем (рис. 6и); VAV 11–12 …………… P. phyllopogon
41(42) Ствол усика сплошь пигментированный, оканчивается единственной луковицей, несущей длинный терминальный пигментированный или непигментированный филамент, часто с дополнительными светящимися тельцами ……..P. albipennis
42(41) Усик с двумя последовательно расположенными луковицами либо с единственной луковицей и с непигментированным (светящимся?) участком примерно посредине ствола …........... 43
43(44) Усик с участком, лишённым пигмента на дорсальной и латеральных поверхностях данного отрезка ствола, но пигментированным вентрально [у взрослых рыб премаксиллярные и максиллярные зубы длиннее, чем у P. coffea (сравнивать одноразмерных рыб!)] ……............... P. barnetti
44(43) Усик с двумя последовательно расположенными луковицами (проксимальная луковица иногда практически не выдаётся за границы ствола, но в таком случае непигментированный участок полностью охватывает данный отрезок ствола) …………………………………………………....... 45
45(46) Усик не короче длины головы; длина участка ствола между луковицами не менее чем вдвое (обычно много больше) превышает длину дистальной луковицы; жаберные лепестки на cera-tobranchiale-1 длинные ………………………... P. coffea
46(45) Усик короче длины головы; длина участка ствола между луковицами обычно не превышает длину дистальной луковицы (как исключение, в 1.5 раза больше её длины); жаберные лепестки в верхней части ceratobranchiale-1 обычно более или менее укорочены (иногда очень слабо, редко совсем не укорочены) ………………... P. waitti
(Отношения между P. coffea и P. waitti нуждаются в дальнейшем изучении. Различия в длине усика и промежутка между луковицами могут быть обусловлены возрастной изменчивостью и P. waitti может оказаться лишь ювенильной стадией P. coffea. Наименьший из исследованных мною особей P. coffea (SL 55 мм, НИС “Витязь”, ст. 7285) имеет промежуток между луковицами, лишь вдвое превышающий длину дистальной луковицы (у всех более крупных рыб он намного больше). Однако у наибольшего из известных экземпляров P. waitti (SL 76 мм, 205–“3”, проба 371) он соизмерим с длиной дистальной луковицы, как и у более мелких особей. В моём материале имеется лишь 1 экз. P. waitti (НИС “Витязь”, ст. 5028, SL 30 мм), у которого промежуток между луковицами превышает (в 1.5 раза) длину дистальной луковицы. Степень укорочения жаберных лепестков в верхней половине ceratobranchiale-1 у P. waitti значительно варьирует, изредка (НИС “Витязь”, ст. 4825 и часть рыб со ст. 5249) они могут быть одинаковой длины по всей длине дуги).
БЛАГОДАРНОСТИ
Я глубоко признателен Дж. Маклейну, К. Уэббу (J. Maclaine, K. Webb, BMNH) и Й. Нильсену (J. Nielsen, ZMUC) за присланные фотографии и данные по синтипам P. dinema и P. leucopogon.
ФИНАНСИРОВАНИЕ РАБОТЫ
Изучение видового состава ихтиофауны мезопелагиали выполнено в рамках темы госзадания № 0149-2018-0009 при поддержке Российского научного фонда (грант № 19-14-00026), изучение морфологии и систематики меланостомовых рыб – в рамках темы госзадания № 0109-2018-0076. Статья написана при частичной поддержке гранта РФФИ № 18-04-00019.
Список литературы
Парин Н.В., Соколовский А.С. 1976. Видовой состав семейства Melanostomiatidae (Osteichthyes) в зоне течения Куросио // Тр. ИО АН СССР. Т. 104. С. 237–249.
Парин Н.В., Беккер В.Э., Бородулина О.Д. и др. 1977. Глубоководные пелагические рыбы западной тропической части Тихого океана и внутренних морей Индо-Малайского архипелага // Там же. Т. 107. С. 68–188.
Прокофьев А.М. 2014. Новые и редкие виды глубоководных пелагических рыб из семейств Opisthoproctidae, Melanostomiidae, Oneirodidae и Linophrynidae // Вопр. ихтиологии. Т. 54. № 4. С. 379–386.
Прокофьев А.М. 2015. Материалы к ревизии подрода Trachinostomias (Melanostomiidae: Photonectes). Морфологическая изменчивость P. margarita с описанием двух новых видов // Там же. Т. 55. № 2. С. 131–138.
Beebe W. 1933. Deep-sea stomiatoid fishes. One new genus and eight new species // Copeia. № 4. P. 160–175.
Bucholtz R.H., Tomkiewicz J., Dalskov J. 2008. Manual to determine gonadal maturity of herring (Clupea harengus L.) // DTU Aqua-report 197-08. Charlottenlund: Natl. Inst. Aquat. Res., 45 p.
Döderlein L. 1882. Ein Stomiatide aus Japan // Arch. Naturgeschichte. V. 48. № 1. P. 26–31.
Fink W.L. 1985. Phylogenetic interrelationships of the stomiid fishes (Teleostei: Stomiiformes). Michigan: Miscell. Publ. Mus. Zool. Univ. № 171. 127 p. + i–vii.
Flynn A.J., Klepadlo C. 2012. Two new species of Photonectes (Teleostei: Stomiidae) from the Indo-Pacific, and a re-examination of P. achirus // Mem. Mus. Victoria. V. 69. P. 259–267.
Gibbs R.H. 1968. Photonectes munificus, a new species of melanostomiatid fish from the south Pacific subtropical convergence, with remarks on the convergence fauna // Contrib. Sci. Los Angeles Co. Mus. № 149. P. 1–6.
Gibbs R.H. 1984. Melanostomiidae // Fishes of the north-eastern Atlantic and the Mediterranean. V. 1 / Eds. Whitehead P.J.P. et al. Paris: UNESCO. P. 341–365.
Gibbs R.H., Barnett M.A. 1990. Melanostomiidae // Check–list of the fishes of the eastern tropical Atlantic (CLOFETA). V. 1 / Eds. Quero J.C. et al. Lisbon; Paris: JNICTSEI; UNESCO. P. 308–337.
Günther A. 1887. Report on the deep-sea fishes collected by H.M.S. Challenger during the years 1873-76 // Rep. Sci. Res. Voy. H.M.S. Challenger. V. 22. Pt. 57. 268 p. + i–lxv.
Eschmeyer W.N. (ed.) 2018. Catalog of Fishes: Genera, Species, References. (http://research.calacademy.org/research/ichthyology/catalog/fishcatmain.asp)
Klepadlo C. 2011. Three new species of the genus Photonectes (Teleostei: Stomiiformes: Stomiidae: Melanostomiinae) from the Pacific Ocean // Copeia. № 2. P. 201–210.
Koefoed E. 1956. Isospondyli. 1. Gymnophotodermi and Lepidophotodermi from the “Michael Sars” North Atlantic deep-sea expedition 1910 // Sci. Res. M. Sars N. Atlantic Deep-Sea Exped. 1910. V. 4. Pt. 2. № 5. P. 1–21.
Morrow J.E., Gibbs R.H. 1964. Melanostomiatidae // Fishes of the Western North Atlantic. V. 1. Pt. 4. New Haven: Sears Found. P. 351–522.
Parr A.E. 1927. The Stomiatoid fishes of the suborder Gymnophotodermi (Astronesthidae, Melanostomiatidae, Idiacanthidae) with a complete review of the species // Bull. Bingham Oceanogr. Collect. Yale Univ. V. 3. Art. 2. 123 p.
Paulin C., Stewart A., Roberts C., McMillan P. 1989. New Zealand fish: a complete guide // Natl. Mus. New Zealand Miscell. Ser. № 19. 279 p.
Paxton J.R., Hoese D.F., Allen G.R., Hanley J.E. 1989. Pisces. Petromyzontidae to Carangidae. Zoological Catalogue of Australia. V. 7. Canberra: Austral. Government Publ. Service, 665 p.
Regan C.T., Trewavas E. 1930. The fishes of the families Stomiatidae and Malacosteidae // Dana Rept. V. 6. P. 1–143.
Richards W.J. 2006. Melanostomiidae: scaleless dragonfishes // Early stages of Atlantic fishes: an identification guide for the Western Central North Atlantic / Ed. Richards W.J. Boca Raton: CRC Press. P. 283–288.
Дополнительные материалы отсутствуют.
Инструменты
Вопросы ихтиологии