

Вопросы ихтиологии, 2020, T. 60, № 5, стр. 617-620
Об обнаружении нерестилища щитоносного ската Bathyraja parmifera в северо-западной части Японского моря
В. В. Панченко 1, *, Д. В. Антоненко 2
1 Национальный научный центр морской биологии Дальневосточного отделения РАН − ННЦМБ ДВО РАН
Владивосток, Россия
2 Тихоокеанский филиал Всероссийского научно-исследовательского института рыбного хозяйства
и океанографии – ТИНРО
Владивосток, Россия
* E-mail: vlad-panch@yandex.ru
Поступила в редакцию 23.09.2019
После доработки 30.09.2019
Принята к публикации 30.09.2019
Аннотация
Впервые сообщается об обнаружении нерестилища щитоносного ската Bathyraja parmifera в северо-западной части Японского моря в координатах 42°21′1″ с.ш. 131°16′6″ в.д., на глубине 216−224 м при придонной температуре 1.0°С. На нерестилище обнаружены производители и яйцевые капсулы.
Скаты подотряда Rajoidei широко распространены в северной части Тихого океана. Щитоносный скат Bathyraja parmifera – широкоборeальный, преимущественно приазиатский вид (Долганов, 1999; Mecklenburg et al., 2002; Парин и др., 2014). В российских водах Японского моря он является монодоминантным видом скатов, на долю которого приходится >90% биомассы обитающих здесь представителей подотряда (Вдовин и др., 2004; Соломатов 2004; Соколовский и др., 2007).
В настоящем сообщении впервые приводятся данные по обнаруженному в северо-западной части Японского моря нерестилищу щитоносного ската.
МАТЕРИАЛ И МЕТОДИКА
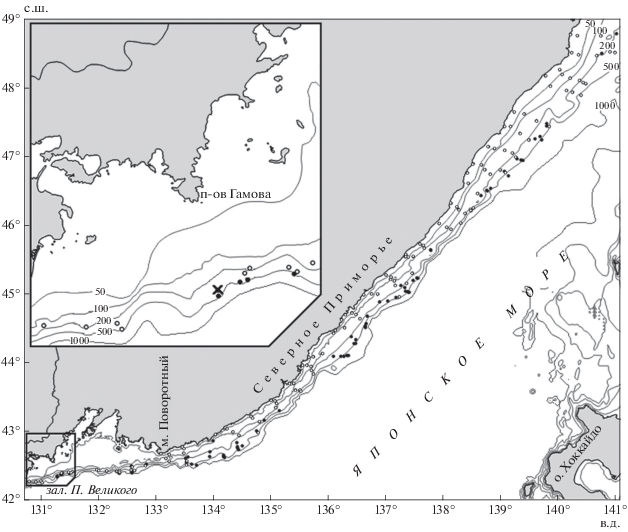
Материалом для работы послужили данные, собранные авторами при выполнении учётной траловой съёмки на научно-исследовательском судне ТИНРО “Бухоро” 18.06−09.08.2007 г. у материкового побережья Японского моря (рисунок). Проведено 214 ловов донным тралом ДТ/ТМ-27.1/24.4 м на глубинах 12−623 м. Всего в период исследований отмечено и промерено (полная длина − TL) 278 особей щитоносного ската. Из массового улова яйцевых капсул скатов была взята выборка (250 шт.) для определения доли пустых капсул и оболочек, содержащих развивающихся эмбрионов. Карта распределения построена с помощью программы Surfer.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
В северо-западной части Японского моря щитоносный скат отмечается в широком диапазоне глубин от 40 до 830 м, однако в шельфовой зоне обитает лишь незначительная часть особей (Антоненко и др., 2007). В период наших исследований он встречался от 105 до 618 м – глубины, близкой к максимальной из исследованных.
В середине июня на траверзе п-ова Гамов в одном тралении помимо рыб и беспозвоночных было отмечено массовое присутствие капсул скатов. Постановка трала на грунт осуществлена в координатах 42°21′1″ с.ш. 131°16′6″ в.д., выборка – 42°22′2″ с.ш. 131°17'7'' в.д. (рисунок). Глубина на месте траления варьировала от 216 м в начале до 224 м в конце. За 30 мин лова было поймано ~76 тыс. капсул, 10% из которых содержали яйца с развивающимися эмбрионами.
Несомненно, что тралением у п-ова Гамова был охвачен район массового нереста щитоносного ската. Об этом свидетельствует массовость обнаруженных яиц, схожесть их с литературными характеристиками капсул этого вида (Ebert, 2005; Hoff, 2019) и нахождение на нерестилище производителей щитоносного ската. Во время траления были пойманы семь взрослых особей: два самца TL 110 и 115 см и пять самок TL 96−118 см, а также неполовозрелый самец длиной 52 см. Половая зрелость самцов этого вида наступает при TL 79.5−91.6 см, самок – 80.9−94.5 см (Долганов, 1998; Ebert, 2005; Орлов, 2006). По данным Хоффа (Hoff, 2008), нерест щитоносного ската проходит преимущественно в летний период, что объясняет нахождение производителей на нерестилище во время выполнения съёмки.
На нерестилище плотность капсул с эмбрионами составила 190 тыс. шт/км2. В Беринговом и северной части Японского моря отмечены нерестилища и с более высокой концентрацией развивающихся в яйцевых оболочках эмбрионов скатов рода Bathyraja, хотя на разных участках этот показатель может сильно варьировать, а нерестилища могут быть локализованы на ограниченных площадках (Hoff, 2010; Hunt et al., 2011). Высокая плотность яиц на нерестовых участках скатов в этих районах обусловлена одновременным развитием генераций нескольких лет. В восточной части беринговоморских вод инкубационный период щитоносного ската длится 3.5 года, в связи с чем на нерестилище одновременно развиваются три когорты эмбрионов (Hoff, 2008).
Длительность инкубационного периода обусловлена температурным режимом вод. У разных видов североатлантических скатов, развивающихся при относительно высокой температуре, он варьирует от 2 до 8 мес. (Wourms, 1977). Так, у Raja eglanteria в экспериментальных условиях при постоянной температуре 20°C потомство вылупляется в среднем через 12 недель (Luer et al., 2007). При слабоположительной температуре эмбрионы развиваются гораздо медленней. У глубоководного ската B. spinosissima, нерестящегося среди вулканических источников Галапагосского рифа, период инкубации при температуре 2.7−2.8°C длится, по-видимому, более 4 лет (Salinas-de-León et al., 2018). На нерестилище щитоносного ската в восточной части Берингова моря, где развитие эмбрионов продолжается 3.5 года, температура несколько выше. За 14-месячный период наблюдений она варьировала в пределах 4.1−5.0°C (в среднем 4.4°C) (Hoff, 2008). Столь высокая температура глубоководной части моря обусловлена непрерывным поступлением относительно тёплых тихоокеанских вод (Хен, 1988). На обнаруженном нами в северо-западной части Японского моря нерестилище щитоносного ската температура воды была ниже (1.0°C), что подразумевает более продолжительный период инкубации. Следует отметить, что в другом районе Японского моря, у западного побережья Хоккайдо, на расположенном на близкой глубине (245−271 м) нерестилище ската Смирнова B. smirnovi, которого иногда рассматривают синонимом щитоносного ската (Долганов, Королев, 2006), температурный фон соответствует отмеченному нами у материкового побережья на нерестилище щитоносного ската: в июле 1.3, в течение года 0.9−2.1°С (Hunt et al., 2011). Подобный годовой градиент температуры характерен для глубин >200 м и в северо-западной части Японского моря (Зуенко, 2008).
Район нахождения обнаруженного нами нерестилища, известный под названием Гамовские свалы, обладает сложным для проведения работ изрезанным скалистым рельефом. В связи с этим на других изобатах в районе нерестилища была найдена возможность для выполнения только одного траления на глубине 300−340 м (рисунок) при придонной температуре воды 0.8°С длительностью 25 мин. В улове здесь была отмечена только одна особь щитоносного ската TL 104 см и единичные оболочки пустых капсул. Остаётся неясным, были они отложены в этом месте или же перенесены течениями или орудиями лова с расположенного на меньшей глубине нерестилища. В любом случае очевидно, что массового нереста скатов в этом месте не происходит.
Рис. 1.
Пространственное распределение щитоносного ската Bathyraja parmifera в северо-западной части Японского моря в июне−августе 2007 г.: (⚪) – траления без улова вида, (⚫) – траления с уловом вида, (×) – траление на нерестилище, (—) − изобаты.
Обращает на себя внимание полное отсутствие на нерестилище молоди скатов. Минимальный размер отмеченных на других участках особей щитоносного ската составлял, как правило, не менее 24 см (исключение – одна особь TL 22 см), хотя в северо-западной части Японского моря встречается его молодь TL ≥ 17 см (Антоненко и др., 2007). Отсутствие мальков можно связать с отсутствием во время проведения работ массового вылупления. Вероятно, в водах Японского моря мальки щитоносного ската, как и на других участках ареала, выходят из яйцевых капсул в основном в холодный период года. Например, в восточной части Берингова моря массовое вылупление щитоносного ската приурочено к осенне-зимнему периоду (Hoff, 2008). Места же концентраций вылупившихся мальков и подрастающей молоди не совпадают (Hoff, 2016).
В выполненных в последующие годы нескольких траловых съёмках подобных массовых поимок капсул скатов не наблюдалось, хотя их некоторое количество в уловах в районе расположения нерестилища присутствовало в каждой съёмке. Площадь нерестилища может быть небольшой. По данным подводных наблюдений, концентрации яиц скатов могут тянуться на дне узкой полосой, в непосредственной близости от которой могут присутствовать лишь единичные капсулы (Hunt et al., 2011). Охватить тралением такой участок сложно, учитывая снос трала течениями, характерными для Гамовских свалов. К тому же в дальнейшем для уменьшения риска нанести ущерб популяции время траления в районе расположения нерестилища скатов, как правило, сокращалось в 1.5−2.0 раза.
В работах Хоффа (Hoff, 2009, 2010), основанных на большом временнóм отрезке данных по нерестилищам нескольких видов из разных районов, показано, что скаты рода Bathyraja стабильно используют для нереста одни и те же участки в течение многих лет. О стабильности расположения нерестового участка с благоприятными для развития эмбрионов факторами среды пишет и Хант с соавторами (Hunt et al., 2011). Местом нереста скатов обычно служат покрытые мягким грунтом или каменистые участки подводных хребтов, расположенные в высокопродуктивных зонах и имеющие циркуляцию водных масс, благоприятную для развития в яйцевых капсулах эмбрионов. Очевидно, подобной зоной является обнаруженная нами в районе Гамовских свалов площадка. Для этого изрезанного скалистого района характерны высокие концентрации гидробионтов, в частности, ценного в промысловом отношении вида – гребенчатой креветки Pandalus hypsinotus (Кобликов, Корнейчук, 2015). О стабильности обнаруженного нами нерестилища щитоносного ската свидетельствует массовое присутствие пустых оболочек яиц, среди которых находились как довольно свежие, хорошо сохранившиеся, так и старые, покрытые обрастателями разлагающиеся капсулы.
Наши данные подтверждают имеющиеся в литературе сведения об использовании щитоносным скатом для нереста расположенные в высокопродуктивных зонах участки подводных хребтов. Более детальная типизация расположения нерестилищ требует дальнейших исследований.
Список литературы
Антоненко Д.В., Пущина О.И., Соломатов С.Ф. 2007. Распределение и некоторые черты биологии щитоносного ската Bathyraja parmifera (Rajidae, Rajiformes) в северо-западной части Японского моря // Вопр. ихтиологии. Т. 47. № 3. С. 311−319.
Вдовин А.Н., Измятинский Д.В., Соломатов С.Ф. 2004. Основные результаты исследований рыб морского прибрежного комплекса Приморья // Изв. ТИНРО. Т. 138. С. 168−190.
Долганов В.Н. 1998. Размножение скатов семейства Rajidae дальневосточных морей России // Там же. Т. 124. С. 425–428.
Долганов В.Н. 1999. Географическое и батиметрическое распространение скатов семейства Rajidae в дальневосточных морях России и сопредельных водах // Вопр. ихтиологии. Т. 39. № 3. С. 428–430.
Долганов В.Н., Королев М.Р. 2006. О валидности скатов группы “parmifera” рода Bathyraja (Rajidae, Rajoidei) // Изв. ТИНРО. Т. 147. С. 179−182.
Зуенко Ю.И. 2008. Промысловая океанография Японского моря. Владивосток: Изд-во ТИНРО-центр, 227 с.
Кобликов В.Н., Корнейчук И.А. 2015. Состояние запасов и перспективы промысла глубоководных креветок в водах Приморья // Вопр. рыболовства. Т. 16. № 1. С. 96−106.
Орлов А.М. 2006. К обоснованию промысловой меры дальневосточных скатов (сем. Rajidae) на примере массовых западноберинговоморских видов // Тр. ВНИРО. Т. 146. Методические аспекты исследований рыб морей Дальнего Востока. С. 252−264.
Парин Н.В., Евсеенко С.А., Васильева Е.Д. 2014. Рыбы морей России: аннотированный каталог. М.: Т-во науч. изд. КМК, 733 с.
Соколовский А.С., Дударев В.А., Соколовская Т.Г., Соломатов С.Ф. 2007. Рыбы российских вод Японского моря: аннотированный и иллюстрированный каталог. Владивосток: Дальнаука, 200 с.
Соломатов С.Ф. 2004. Характеристика ихтиофауны морских вод северного Приморья (Японское море) // Изв. ТИНРО Т. 138. С. 205−219.
Хен Г.В. 1988. Сезонная и межгодовая изменчивость вод Берингова моря и её влияние на распределение и численность гидробионтов: Автореф. дис. … канд. геогр. наук. М.: ВНИРО, 24 с.
Ebert D.A. 2005. Reproductive biology of skates, Bathyraja (Ishiyama), along the eastern Bering Sea continental slope // J. Fish Biol. V. 66. P. 618–649. https://doi.org/10.1111/j.1095-8649.2005.00628.x
Hoff G.R. 2008. A nursery site of the Alaska skate (Bathyraja parmifera) in the eastern Bering Sea // Fish. Bull. V. 106. № 3. P. 233–244.
Hoff G.R. 2009. Embryo developmental events and the egg case of the Aleutian skate Bathyraja aleutica (Gilbert) and the Alaska skate Bathyraja parmifera (Bean) // J. Fish Biol. V. 74. P. 483–501. https://doi.org/10.1111/j.1095-8649.2008.02138.x
Hoff G.R. 2010. Identification of skate nursery habitat in the eastern Bering Sea // Mar. Ecol. Prog. Ser. V. 403. P. 243–254. https://doi.org/10.3354/meps08424
Hoff G.R. 2016. Identification of multiple nursery habitats of skates in the eastern Bering Sea // J. Fish Biol. V. 88. № 5. P. 1746–1757. https://doi.org/10.1111/jfb.12939
Hoff G.R. 2019. Where do baby skates come from? (https:// www.afsc.noaa.gov/Education/pdfs/Eggcase_brochure.pdf)
Hunt J.C., Lindsay D.J., Shahalemi R.R. 2011. A nursery site of the golden skate (Rajiformes: Rajidae: Bathyraja smirnovi) on the Shiribeshi Seamount, Sea of Japan // Mar. Biodiv. Rec. V. 4. P. 1–7. https://doi.org/10.1017/S1755267211000728
Luer C.A., Walsh C.J., Bodine A.B., Wyffels J.T. 2007. Normal embryonic development in the clearnose skate, Raja eglanteria, with experimental observations on artificial insemination // Environ. Biol. Fish. V. 80. P. 239−255. https://doi.org/10.1007/s10641-007-9219-4
Mecklenburg C.W., Mecklenburg T.A., Thorsteinson L.K. 2002. Fishes of Alaska. Bethesda, Maryland: Amer. Fish. Soc., 1037 p.
Salinas-de-León P., Phillips B., Ebert D. et al. 2018. Deep-sea hydrothermal vents as natural egg-case incubators at the Galapagos Rift // Sci. Rept. V. 8. № 1. Article № 1788. https://doi.org/10.1038/s41598-018-20046-4
Wourms J.P. 1977. Reproduction and development of chondrichthyan fishes // Amer. Zool. V. 17. P. 379−410.
Дополнительные материалы отсутствуют.
Инструменты
Вопросы ихтиологии