

Вопросы ихтиологии, 2021, T. 61, № 2, стр. 167-176
Распределение, размерно-половой состав Gymnocanthus pistilliger (Cottidae) у западного побережья Камчатки в летний период и современное состояние его запасов
А. А. Матвеев 1, *, А. А. Баланов 2, В. В. Панченко 2
1 Камчатский филиал Всероссийского научно-исследовательского института
рыбного хозяйства и океанографии – КамчатНИРО
Петропавловск-Камчатский, Россия
2 Национальный научный центр морской биологии Дальневосточного отделения РАН – ННЦМБ ДВО РАН
Владивосток, Россия
* E-mail: matveev.a.a@kamniro.ru
Поступила в редакцию 03.03.2020
После доработки 13.03.2020
Принята к публикации 17.03.2020
Аннотация
В Охотском море, у западного побережья Камчатки, нитчатый шлемоносец Gymnocanthus pistilliger в летний период 2017 г. встречался на глубинах от 14 (минимальная глубина тралений) до 64 м. Районы максимальных концентраций G. pistilliger по массе и численности совпадают лишь частично. Наибольшие средние уловы по биомассе отмечены между 55° и 56° с.ш., где также зарегистрирована высокая численность вида. Максимальные средние уловы по численности получены южнее – между 53° и 54° с.ш. Наиболее обычен и обилен этот вид на глубинах до 40−50 м. Длина особей в уловах варьирует в пределах 6–26 (в среднем 18.3) см. Соотношение самцов к самкам – 0.4 : 1. Наименьшие размеры особей обоих полов отмечены в диапазоне глубин 14−20 м; с возрастанием глубины средние размеры самок и самцов увеличиваются. В летний период на западнокамчатском шельфе G. pistilliger встречается в широком диапазоне температуры: 0.4–12.1°С. Наибольшие уловы вида в основной части батиметрического интервала (20–50 м) приурочены к температуре 1.0–3.5°С, а в мелководной зоне (14–20 м) – 3.5–6.0°С. По результатам летних работ 2017 г. у западного побережья Камчатки учтённая численность G. pistilliger составила 146 млн экз., биомасса – 11.2 тыс. т.
Среди представителей семейства Cottidae виды рода Gymnocanthus наиболее широко распространены (Андрияшев, 1954; Wilson, 1973). Одним из шести видов этого рода, обитающих в Северном полушарии, является нитчатый шлемоносец Gymnocanthus pistilliger. Арктическо-бореальный вид, широко распространённый в Северо-Западной Пацифике от южной части Японского моря до юго-восточного побережья Аляски, в Беринговом и Охотском морях, вдоль тихоокеанского побережья Курильских о-вов и о-ва Хоккайдо, а также в Чукотском море (Линдберг, Красюкова, 1987; Allen, Smith, 1988; Борец, 2000; Mecklenburg et al., 2002; Новиков и др., 2002; Федоров и др., 2003; Yamazaki et al., 2013). Считается промысловым видом, однако промыслом используется слабо, оставаясь перспективным объектом рыболовства (Токранов, 1987; Панченко, 2013).
Целенаправленные исследования распределения и других аспектов биологии нитчатого бычка проводились в российских водах Японского моря, преимущественно в зал. Петра Великого (Вдовин и др., 1994; Вдовин, Дударев, 2000; Шелехов, Панченко, 2007; Панченко, 2012, 2013; Панченко и др., 2020), у восточного побережья Берингова моря (Hoff, 2000) и в прикамчатских водах (Токранов 1981а, 1981б, 1985а, 1985б, 1987, 1993). При этом в литературе до сих пор встречаются противоречивые данные о предельных глубинах его обитания (Андронов, Датский, 2014; Матвеев, Терентьев, 2016; Баланов, Матвеев, 2018). Наибольшей численности этот вид достигает у западного побережья Камчатки, в водах Охотского моря (Токранов, 2017).
Имеющиеся в нашем распоряжении данные позволяют дополнить и уточнить сведения о пространственном и батиметрическом распределении, описать размерный и половой состав, а также оценить состояние запасов нитчатого шлемоносца у западного побережья Камчатки в летний период.
МАТЕРИАЛ И МЕТОДИКА
В основу работы положен материал, собранный на шельфе Западной Камчатки в Охотском море во время донной траловой съёмки на НИС “ТИНРО” 22.06−31.07.2017 г. Траления выполняли в пределах Западно-Камчатской и Камчатско-Курильской рыбопромысловых подзон (50°58′− 57°28′ с.ш.; граница между подзонами проходит по 54° с.ш.) по стандартной сетке станций (рис. 1). Работы выполняли в светлое время суток, со средней скоростью траления 2.9 узла. Орудием лова служил донный трал типа ДТ 27.1 (горизонтальное раскрытие 16.3 м, длина верхней подборы 27.1 м, периметр трала 33.7 м). Всего проведено 255 учётных тралений на глубинах от 14 до 409 м. Так как нитчатый шлемоносец в период съёмки не встречался глубже 64 м, сведения по его встречаемости, уловах, температуре обитания и оценки запасов рассчитаны для диапазона глубин 14–64 м. В этом диапазоне выполнено 133 траления (табл. 1).
Рис. 1.
Схема траловых станций (⚫) у западного побережья Камчатки, июнь–июль 2017 г.: 54° с.ш. – граница между Западно-Камчатской и Камчатско-Курильской промысловыми подзонами, (–) – изобаты 20, 50 и 100 м.
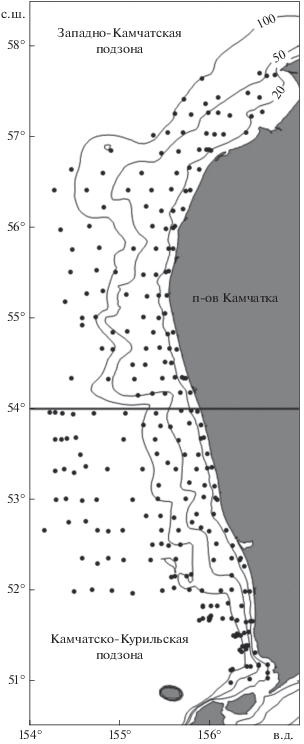
Таблица 1.
Число тралений, выполненных на шельфе Западной Камчатки на глубинах 14−64 м в июне–июле 2017 г.
| Диапазон глубин, м | Расположение участков траления, ° с.ш. | |||||||
|---|---|---|---|---|---|---|---|---|
| 51–52 | 52–53 | 53–54 | 54–55 | 55–56 | 56–57 | 57–58 | 51–58 | |
| 14.0–20.0 | 3 | 4 | 8 | 5 | 4 | 5 | 2 | 31 |
| 20.1–30.0 | 3 | 3 | 4 | 3 | 5 | 5 | 3 | 26 |
| 30.1–40.0 | 5 | 5 | 5 | 4 | 2 | 1 | 1 | 23 |
| 40.1–50.0 | 4 | 3 | 1 | 4 | 3 | 3 | 3 | 21 |
| 50.1–64.0 | 7 | 5 | 7 | 3 | 3 | 4 | 3 | 32 |
| 14.0–64.0 | 22 | 20 | 25 | 19 | 17 | 18 | 12 | 133 |
Разбор улова и ихтиологические исследования выполняли по общепринятым методикам (Борец, 1997). Ранее были установлены надёжные диагностические признаки G. pistilliger (Баланов, Матвеев, 2018), что позволило идентифицировать особей этого вида с высокой точностью. У рыб измеряли полную длину (TL)11 и массу; всего промерено 3310 экз. G. pistilliger, из них 2361 самок и 949 самцов. Расчёт запасов проводили методом сплайн-аппроксимации с использованием компьютерной геоинформационной системы ГИС “КартМастер”, разработанной ВНИРО для обработки данных разнообразных биоресурсных съёмок, в том числе и траловых (Бизиков и др., 2007). Для расчёта численности и биомассы принят коэффициент уловистости, равный 0.5 (Борец, 1985). В тексте и таблицах данные по уловам приводятся в расчёте на 1 ч траления (кг/ч и экз/ч).
Температуру придонного слоя воды измеряли CTD-зондом SBE 19plus V2 (Sea-Bird Electronics, Inc., США). Результаты фоновых наблюдений обработаны с использованием пакета программ фирмы-производителя CTD-зонда. Для построения карт распределения использовали специализированную программу Surfer ver. 11.
РЕЗУЛЬТАТЫ
В июне−июле 2017 г. G. pistilliger встречался вдоль всей исследованной акватории западнокамчатского шельфа (рис. 2). За исключением южных акваторий, располагающихся между 51°–52° с.ш., частота встречаемости данного вида была высока. Особенно это характерно для участков между 56° и 58° с.ш., однако здесь, в северной части исследованного района, его уловы оказались небольшими (табл. 2). Распределение уловов G. pistilliger по исследованному району характеризовалось значительной неоднородностью. Максимальный улов (300 кг/ч, 2700 экз/ч) получен в координатах 55°06′ с.ш. и 155°50′ в.д. Средний улов за весь период работ составил 32 кг/ч, или 454 экз/ч
Рис. 2.
Распределение нитчатого шлемоносца Gymnocanthus pistilliger на западнокамчатском шельфе в июне–июле 2017 г.: а – масса, кг/ч траления; б – численность, экз/ч траления; (─) – изотермы придонного слоя воды, °С.
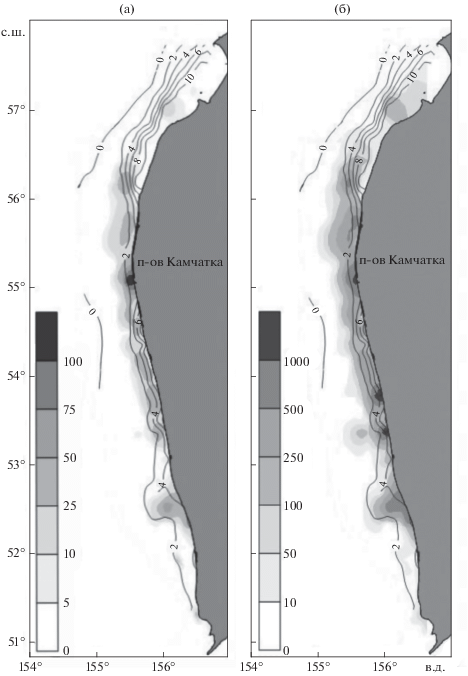
Таблица 2.
Встречаемость, средние значения уловов и массы особи нитчатого шлемоносца Gymnocanthus pistilliger в разных широтных участках западнокамчатского шельфа в июне–июле 2017 г.
| Показатель | Расположение участков траления, ° с.ш. | В среднем | ||||||
|---|---|---|---|---|---|---|---|---|
| 51−52 | 52−53 | 53−54 | 54−55 | 55−56 | 56−57 | 57−58 | ||
| Встречаемость, % | 31.8 | 65.0 | 88.0 | 89.5 | 94.1 | 94.4 | 91.7 | 77.4 |
| Уловы за 1 ч траления: | ||||||||
| – масса, кг | 6.7 | 39.6 | 37.8 | 37.8 | 55.0 | 13.8 | 10.0 | 32.1 |
| – число рыб, экз. | 109 | 547 | 626 | 454 | 685 | 166 | 135 | 454 |
| Средняя масса особи, г | 61 | 72 | 60 | 83 | 80 | 83 | 74 | 71 |
| Число тралений | 22 | 20 | 25 | 19 | 17 | 18 | 12 | 133 |
Районы наибольших концентраций G. pistilliger по массе и численности совпадали лишь частично. При осреднении через 1° наибольшие средние уловы по массе получены между 55° и 56° с.ш., где также отмечалась высокая численность вида. Тогда как максимальные средние уловы по численности отмечены южнее – между 53° и 54° с.ш., однако в весовом выражении они здесь были заметно ниже, чем в вышеуказанном северном диапазоне широт (рис. 2, табл. 2).
Суммарно по двум подзонам учтённая биомасса G. pistilliger составила 11.2 тыс. т, а численность – 146.0 млн экз.; абсолютные и относительные показатели биомассы и численности в Западно-Камчатской подзоне в 1.4−1.5 раза выше, чем в Камчатско-Курильской (табл. 3).
Таблица 3.
Запасы нитчатого шлемоносца Gymnocanthus pistilliger у западного побережья Камчатки, по данным донной траловой съёмки (июнь−июль 2017 г.)
| Показатель | Подзонa | Всего | |
|---|---|---|---|
| Западно-Камчатская | Камчатско-Курильская | ||
| Биомасса, тыс. т | 6.7 | 4.5 | 11.2 |
| Численность, млн экз. | 84.9 | 61.1 | 146.0 |
| Плотность скоплений: | |||
| – биомасса, кг/км2 | 217.3 | 140.6 | 178.5 |
| – численность, экз/км2 | 2745.1 | 1927.2 | 2330.9 |
На западнокамчатском шельфе G. pistilliger встречался на глубинах от 14 до 64 м, хотя наиболее обычен он был до 50-метровой изобаты (табл. 4). Максимальная частота встречаемости и наибольшее число особей в улове за час траления отмечены на глубинах 14–20 м; самый высокий средний улов по массе – в диапазоне 20–30 м; с увеличением глубины все эти показатели снижались. Минимальные уловы (<2 кг/ч) наблюдались на глубинах более 50 м.
Таблица 4.
Встречаемость, средние значения уловов, размеры нитчатого шлемоносца Gymnocanthus pistilliger и температура в разных диапазонах глубин западнокамчатского шельфа в июне–июле 2017 г.
| Показатель | Диапазон глубин, м | В среднем | ||||
|---|---|---|---|---|---|---|
| 14.0–20.0 | 20.1–30.0 | 30.1–40.0 | 40.1–50.0 | 50.1–64.0 | ||
| Встречаемость, % | 90.3 | 96.2 | 82.6 | 85.7 | 40.6 | 77.4 |
| Уловы за 1 ч траления: | ||||||
| – масса, кг | 30.0 | 51.4 | 35.9 | 22.8 | 6.6 | 32.1 |
| – число рыб, экз. | 625 | 620 | 416 | 265 | 81 | 454 |
| Средняя масса особи, г | 48 | 83 | 86 | 86 | 81 | 71 |
| Длина (TL), см: | ||||||
| – самки | $\frac{{17.6\,\,(607)}}{{8{\kern 1pt} - {\kern 1pt} 25}}$ | $\frac{{19.4\,\,(1218)}}{{10{\kern 1pt} - {\kern 1pt} 26}}$ | $\frac{{20.6\,\,(267)}}{{9{\kern 1pt} - {\kern 1pt} 25}}$ | $\frac{{19.6\,\,(215)}}{{13{\kern 1pt} - {\kern 1pt} 26}}$ | $\frac{{20.6\,\,(54)}}{{13{\kern 1pt} - {\kern 1pt} 26}}$ | $\frac{{19.1\,\,(2361)}}{{8{\kern 1pt} - {\kern 1pt} 26}}$ |
| – самцы | $\frac{{14.0\,\,(204)}}{{6{\kern 1pt} - {\kern 1pt} 21}}$ | $\frac{{16.3\,\,(413)}}{{7{\kern 1pt} - {\kern 1pt} 22}}$ | $\frac{{16.9\,\,{\text{(}}187{\text{)}}}}{{10{\kern 1pt} - {\kern 1pt} 23}}$ | $\frac{{16.0\,\,(131)}}{{12{\kern 1pt} - {\kern 1pt} 22}}$ | $\frac{{17.0\,\,(14)}}{{12{\kern 1pt} - {\kern 1pt} 20}}$ | $\frac{{15.9\,\,{\text{(}}949{\text{)}}}}{{6{\kern 1pt} - {\kern 1pt} 23}}$ |
| Температура у дна, °С | 6.7 | 5.2 | 3.1 | 3.2 | 1.8 | 4.4 |
| Число тралений | 31 | 26 | 23 | 21 | 32 | 133 |
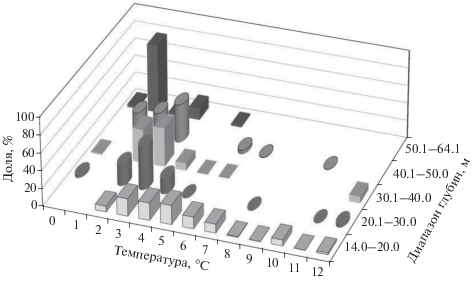
Уловы G. pistilliger отмечены в широком диапазоне температуры придонного слоя воды: 0.4−12.1°С (рис. 2, 3; табл. 4). Стабильно высокие концентрации были приурочены к температурному фону 1.0−3.0°С на глубинах 20–50 м. На малых глубинах (14–20 м) относительно высокие уловы зарегистрированы в температурном диапазоне 3.0–6.0°С. (рис. 3). Наиболее высокие значения температуры придонного слоя наблюдались в районе к северу от 56° с.ш. (рис. 2), где воды северной ветви Западно-Камчатского течения попадают в зал. Шелихова, в результате часть шельфа вокруг м. Южный занята прогретыми и относительно распреснёнными водами антициклонической циркуляции (Фигуркин, 2002). В этом районе небольшие уловы G. pistilliger отмечены в интервале глубин 30–50 м, где температура придонного слоя воды составляла 10–12°С.
Рис. 3.
Доля (% общей массы уловов) нитчатого шлемоносца Gymnocanthus pistilliger в разных диапазонах глубин в зависимости от придонной температуры на западнокамчатском шельфе в июне–июле 2017 г.
Длина особей G. pistilliger в уловах варьировала в пределах 6−26 (в среднем 18.3) см (рис. 4). Самки в уловах были представлены особями TL 8−26 (19.1) см, среди них преобладали рыбы TL 18−22 см (65%). Размерный ряд самцов по сравнению с самками короче, а средняя длина заметно меньше − 6−23 (15.9) см; доминировали особи TL 15−19 см (около 67%). В целом соотношение самцов к самкам равнялось 0.4 : 1. Среди мелких рыб (TL < 15 см) самцы численно преобладали над самками. При увеличении длины соотношение полов выравнивалось, и по достижении TL 18 см доля самок в каждом последующем сантиметровом классе возрастала вплоть до полного отсутствия самцов среди рыб TL > 22 см.
Рис. 4.
Размерный состав (TL) уловов нитчатого шлемоносца Gymnocanthus pistilliger у западного побережья Камчатки в июне–июле 2017 г.: ( ) – самки, (
) – самки, (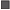 ) – самцы, (─) – оба пола.
) – самцы, (─) – оба пола.
У особей обоих полов наименьшие средние значения длины и массы зарегистрированы в диапазоне малых глубин – 14–20 м (табл. 4). Далее отмечалось увеличение средних размеров самок и самцов. Широкий диапазон варьирования длины у особей обоих полов отмечался на глубинах 14–40 м; с увеличением глубины размерный ряд был заметно короче, особенно среди самцов.
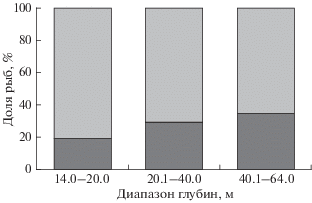
Соотношение полов у исследуемого вида в разных диапазонах глубин было не одинаково (рис. 5). В целом в уловах преобладали самки (73%), их доминирование с увеличением глубины постепенно снижалось с 81 (14−20 м) до 65% (40−64 м).
Рис. 5.
Соотношение самок и самцов нитчатого шлемоносца Gymnocanthus pistilliger в разных диапазонах глубин на западнокамчатском шельфе в июне−июле 2017 г.; обозначения см. на рис. 4.
ОБСУЖДЕНИЕ
На западнокамчатском шельфе в летний период 2017 г. частота встречаемости G. pistilliger была высока во всех широтных диапазонах по всему исследованному району. Он относится к характерным и массовым видам восточной части Охотского моря (Токранов, 1981б; Борец, 1989; Баланов, Матвеев, 2018).
Довольно сложно провести сравнение летнего распределения G. pistilliger на шельфе Западной Камчатки с данными литературы. Предыдущие исследования проводились разными орудиями лова и не всегда в сравниваемые сроки (Токранов, 1981б). Кроме того, имеются обоснованные сомнения в верности видовой идентификации данного вида в отдельных съёмках (Матвеев, Терентьев, 2016). Тем не менее практически во всех работах отмечено, что G. pistilliger встречается по всему западнокамчатскому шельфу на глубине до 60–80 м; его основные концентрации, как и в период наших работ, обычно фиксируются в центральной и северной частях данного района между 53° и 57° с.ш. (Токранов, 1981б; Матвеев, Терентьев, 2016).
Впервые обнаружено несовпадение пространственного распределения уловов G. pistilliger по численности и биомассе (рис. 2). В южной части исследованного района (между 53° и 54° с.ш.) доля мелких рыб (с невысокой средней массой) была значительно больше, чем в его северной части (55°−57° с.ш.) (табл. 2). На данный момент трудно судить о стабильности такого характера распределения G. pistilliger в летний период, поскольку в источниках литературы нет данных для сравнения.
По сведениям Токранова (1981а, 1981б), G. pistilliger на западнокамчатском шельфе летом встречается начиная с минимальных глубин исследования, варьирующих в научных донных траловых съёмках от 10 до 15 м. Мы также отмечали его поимки начиная с минимальных глубин, причём наибольшая плотность рыб оказалась приурочена к глубинам <20 м. Исходя из этого, можно предположить, что часть популяции в летний период обитает и на меньших, не охватываемых съёмками глубинах, в связи с чем часть запаса остаётся недоучтённой. Выявленная нами тенденция уменьшения размеров и массы рыб с уменьшением глубины позволяет констатировать, что недоучёт численности происходит в большей степени, чем недоучёт биомассы.
Можно предположить, что повышенные уловы рыб мелкого размера между 53° и 54° с.ш. вызваны теми же причинами. В северной части исследованного района шельф шире (рис. 1), и, по-видимому, мелкие особи G. pistilliger распределяются по нему и в меньшей степени выходят на глубины >10–15 м, в связи с чем не учитываются в уловах. Южнее 54° с.ш. мелководная часть шельфа более узкая, и молодь вынуждена выходить на глубины >10–15 м в большей степени, что и ведёт за собой увеличение численности в уловах (рис. 2). Нужны дополнительные исследования (в первую очередь, на глубинах 0–15 м), чтобы проверить это предположение.
Представленные в настоящей работе данные о батиметрическом распределении G. pistilliger в охотоморских водах Западной Камчатки в летний период близки к данным Токранова (1981а, 1981б) и Борца (1989). Наиболее заселёнными этим видом на шельфе Западной Камчатки являются глубины от 10–15 до 40–50 м. В летний период особи G. pistilliger в этом районе, как правило, не опускаются глубже 65 м. Лишь отдельные экземпляры встречаются на глубинах 60–80 м (Токранов, 1981б; Борец, 1989). Вместе с тем имеются сведения об обитании G. pistilliger летом в охотоморских прикамчатских водах и глубже. По результатам летней траловой съёмки, выполненной в 1996 г. у западного побережья Камчатки, максимальные глубины поимок этого вида отмечены между изобатами 201 и 300 м (Терентьев, 1998). Однако эти данные значительно отличаются от массива достоверных сведений о вертикальном распределении G. pistilliger (Токранов, 1981a, 1981б, 1987, 1988; Борец, 1989; наши данные). Можно предположить, что это вызвано либо неверным определением вида (за G. pistilliger принимался другой вид рода Gymnocanthus), либо в улове были особи из предыдущего мелководного траления. Подробно эти причины разобраны ранее (Баланов, Матвеев, 2018).
Известно, что в восточной части Берингова моря в летний период G. pistilliger обитает в прибрежных водах, как правило, на глубинах <50 м (Hoff, 2000). По нашим данным, в водах Западной Камчатки вертикальное распределение этого вида очень похоже на то, что известно для восточной части Берингова моря (Токранов, 1981a, 1981б; Борец, 1989). В южной части ареала, в зал. Петра Великого (Вдовин и др., 1994; Панченко, Зуенко, 2009), как и в целом в Японское море (Панченко и др., 2020), G. pistilliger в летний период на глубинах <15 м представлен лишь эпизодическими поимками и концентрируется в батиметрическом диапазоне 40−80 м.
По данным Токранова (1981а, 1981б), G. pistilliger на западнокамчатском шельфе в летний период имеет относительно широкую термопатию – 0–12°C, а наибольшее число особей встречается в диапазоне 6–12°С. Наши данные подтверждают, что G. pistilliger в этом районе в летний период обитает при положительной температуре в тех же пределах, однако предпочитаемый температурный фон был ниже: наибольшие уловы на глубинах > 20 м были получены при температуре 1–3°С, а в мелководной зоне (14–20 м) – 3–6°С (рис. 3). Объективность полученной информации о температурных пределах обитания G. pistilliger для июня–июля подтверждается данными нашего предыдущего исследования, когда в эти же месяцы 2013 и 2014 гг. основные его скопления отмечались при средней температуре воды у дна – соответственно 2.9 и 3.0°С (Матвеев, Терентьев, 2016). Различия в наших данных и данных Токранова (1981а, 1981б), наиболее вероятно, вызваны двумя причинами. Во-первых, Токранов (1981б. Табл. 3) не приводит точные глубины, на которых собраны данные для таблицы в его работе. Если основной материал получен на глубинах до 20 м, то вполне понятно, что ряд предпочтений вида по температуре должен быть сдвинут в сторону более высоких значений. Во-вторых, в 1977–1979 гг. материал собирали в июне–сентябре без разделения на месяцы (Токранов, 1981б). Если предположить, что основные сборы выполняли в августе–сентябре, то данные Токранова (1981а, 1981б) отражают термопатию G. pistilliger в наиболее тёплый период года, тогда как наши данные – в более холодный период начала летнего прогрева вод шельфа Западной Камчатки (Фигуркин, 2002).
Более высокие температурные предпочтения вида в мелководной зоне в значительной степени обусловлены обитанием молоди в прогреваемом мелководье. Напомним, что размеры и масса рыб с глубиной в целом увеличиваются. Вертикальную зональность размерных групп ранее также отмечал Токранов (1981а, 1987). Подобные различия в батиметрическом распределении особей G. pistilliger разного размера описаны и в южной части ареала (Панченко, Зуенко, 2009; Панченко и др., 2020).
Возможно, особенностями гидрологического режима обусловлено и различие в размерах, достигаемых G. pistilliger в разных районах обитания. На юге предельная длина этого вида составляет 33 см (Панченко и др., 2016), в более же суровых условиях восточной части Берингова моря – лишь 20.1 см (Hoff, 2000). По данным Токранова (1987), у охотоморского побережья Камчатки предельный размер G. pistilliger составляет 27 см. В наших уловах этот показатель несколько меньше − 26 см, что объясняется низкой численностью рыб крупного размера, в связи с этим они присутствуют в уловах далеко не в каждой съёмке. Отсутствие в уловах молоди TL < 6 см, по всей видимости, обусловлено тем, что сеголетки обитают на глубинах <14 м. Ранее Токранов (1981а) отмечал, что с увеличением глубины средняя длина особей G. pistilliger возрастает более чем на 6 см. Наши данные подтверждают указанную тенденцию, но этот показатель увеличивается лишь на 3 см: у самцов − с 14.0 до 17.0 см, у самок − с 17.6 до 20.6 см (табл. 4).
Для G. pistilliger характерен половой диморфизм, проявляющийся, в частности, в достижении самками больших, чем самцы, размеров. В разных районах самки численно преобладают над самцами. Это наблюдается в Беринговом (Hoff, 2000) и Японском морях (Панченко, 2012), в водах западнокамчатского шельфа (Токранов, 1987, 2016; наши данные). Происходит это за счёт повышения с возрастанием размеров доли самок в нерестовой части популяции в результате большей продолжительности их жизни (Токранов, 1987; Панченко, 2012; Токранов, Орлов, 2012). Видимо, преобладание самок в популяциях G. pistilliger является составной частью жизненной стратегии вида.
Соотношение полов в разных диапазонах глубин у исследуемого вида не одинаково. Выявлена достаточно выраженная тенденция уменьшения доли самок с возрастанием глубины (рис. 5). Схожая (только более ярко выраженная) картина снижения летом доли самок с увеличением глубины отмечена и в Беринговом море (Hoff, 2000). Вероятно, подобное распределение полов в летний период характерно для северных районов, тогда как на юге ареала, в зал. Петра Великого, отмечена обратная тенденция: летом с возрастанием глубины доля самок увеличивается (Панченко и др., 2020).
Столь резкие различия в вертикальном распределении G. pistilliger (в целом и между полами) между северными районами и Японским морем, по-видимому, обусловлены особенностями гидрологии этих регионов. Если в восточной части Берингова и Охотского морей температура 2–12°С в летний период характерна именно для верхнего 50-метрового слоя (Морошкин, 1966; Арсеньев, 1967), то в Японском море температура верхних 20–30 м может достигать 22–24°С (Зуенко, 2008), что явно неприемлемо для G. pistilliger. На крайнем юге своего ареала данный вид находит условия для нормального существования, уходя на бóльшие глубины.
В прикамчатских водах G. pistilliger нерестится в декабре–январе на глубинах 80–110 м при слабоположительной (1.0–1.2°С) температуре придонных вод (Токранов, 1987). К сожалению, автор не приводит данные о соотношении полов во время нереста. Однако есть сведения из зал. Петра Великого (Японское море), что в преднерестовых скоплениях соотношение полов близко 1 : 1, хотя в нагульный период самок всегда больше (Вдовин и др., 1994). Из этого можно предположить, что летом, в нагульный период, увеличение доли самцов с глубиной в северных областях ареала вызвано смещением части самцов в сторону нерестилищ, куда они подходят раньше самок. В зал. Петра Великого (а, вероятно, и в целом в южной части ареала) нерест, также приуроченный к холодному периоду года, проходит на гораздо меньших глубинах верхней части шельфа (Вдовин и др., 1994; Панченко и др., 2020).
Первые известные оценки биомассы G. pistilliger, обитающего у западного побережья Камчатки, приведены в работах Борца (1985) и Токранова (1988) − соответственно 1.0 и 5.4 тыс. т. По данным Савина с соавторами (2011), в июле–августе 2010 г. биомасса G. pistilliger здесь составляла чуть более 5.1 тыс. т, т.е. была близка к оценкам второго автора. Однако в основном этот показатель был выше и в отдельные годы приближался к 10 тыс. т и более: 9.9 (2007), 11.3 (2002) и 13.7 тыс. т (2005). В среднем для периода 1982–2010 гг. биомасса G. pistilliger западнокамчатского шельфа оценивалась на уровне 7.3 тыс. т. Горбатенко и Савин (2012) приводят оценки биомассы G. pistilliger в качестве кормового объекта для массовых демерсальных видов рыб: среднемноголетнее за период 2005–2010 гг. потребление нитчатого шлемоносца составляет 25.5 тыс. т, соответственно запас должен быть на порядок выше, что, по нашему мнению, определённо завышенная оценка. По данным Шунтова и Темных (2018), среднемноголетний уровень биомассы G. pistilliger западнокамчатского шельфа для 1977−2010 гг. составляет 12.7 тыс. т. Наши усреднённые многолетние оценки биомассы вида для периода 1986−2015 гг. находились на том же уровне, составляя чуть более 10 тыс. т (Матвеев, Терентьев, 2016).
По данным настоящего исследования, летом 2017 г. у западного побережья Камчатки учтённая численность G. pistilliger составляет 146 млн экз., биомасса – 11.2 тыс. т, что соответствует среднемноголетним оценкам и может свидетельствовать о стабильном состоянии популяции исследуемого района.
Список литературы
Андрияшев А.П. 1954. Рыбы северных морей СССР. М.; Л.: Изд-во АН СССР, 566 с.
Андронов П.Ю., Датский А.В. 2014. Закономерности пространственной изменчивости структуры донных сообществ рыб шельфа северо-западной части Берингова моря // Изв. ТИНРО. Т. 177. С. 40–76.
Арсеньев В.С. 1967. Течения и водные массы Берингова моря. М.: Наука, 135 с.
Баланов А.А., Матвеев А.А. 2018. Диагностика Gymnocanthus pistilliger (Pallas, 1814) и G. galeatus Bean, 1881 (Scorpaeniformes: Cottidae) в районах их совместного обитания // Биология моря. Т. 44. № 5. С. 365–368. https://doi.org/10.1134/S0134347518050091
Бизиков В.А., Гончаров С.М., Поляков А.В. 2007. Географическая информационная система “Картмастер” // Рыб. хоз-во. № 1. С. 96−99.
Борец Л.А. 1985. Состав и биомасса донных рыб на шельфе Охотского моря // Биология моря. № 4. С. 54–59.
Борец Л.А. 1989. Закономерности вертикального распределения донных рыб в летний период на западнокамчатском шельфе // Вопр. ихтиологии. Т. 29. Вып. 3. С. 370–376.
Борец Л.А. 1997. Донные ихтиоцены Российского шельфа дальневосточных морей: состав, структура, элементы функционирования и промысловое значение. Владивосток: Изд-во ТИНРО-центр, 217 с.
Борец Л.А. 2000. Аннотированный список рыб дальневосточных морей. Владивосток: Изд-во ТИНРО-центр, 192 с.
Вдовин А.Н., Дударев В.А. 2000. Сравнительная оценка количественных учетов рыбной сырьевой базы Приморья // Вопр. рыболовства. Т. 1. № 4. С. 46–57.
Вдовин А.Н., Швыдкий Г.В., Афанасьева Н.И. и др. 1994. Пространственно-временная изменчивость распределения нитчатого шлемоносца в заливе Петра Великого // Экология. № 4. С. 53–59.
Горбатенко К.М., Савин А.Б. 2012. Состав, биомасса и трофические характеристики рыб на западнокамчатском шельфе // Изв. ТИНРО. Т. 171. С. 40–61.
Зуенко Ю.И. 2008. Промысловая океанография Японского моря. Владивосток: Изд-во ТИНРО-центр, 227 с.
Линдберг Г.У., Красюкова З.В. 1987. Рыбы Японского моря и сопредельных частей Охотского и Желтого морей. Ч. 5. Л.: Наука, 526 с.
Матвеев А.А., Терентьев Д.А. 2016. Промысел, многолетняя динамика биомассы, распределение и размерный состав массовых видов рогатковых Cottidae у западного побережья Камчатки // Исследования водных биологических ресурсов Камчатки и северо-западной части Тихого океана. № 41. С. 17–42. https://doi.org/10.15853/2072-8212.2016.41.17-42
Морошкин К.В. 1966. Водные массы Охотского моря. М.: Наука, 68 с.
Новиков Н.П., Соколовский А.С., Соколовская Т.Г., Яковлев Ю.М. 2002. Рыбы Приморья. Владивосток: Изд-во Дальрыбвтуз, 552 с.
Панченко В.В. 2012. Возраст и рост шлемоносных бычков рода Gymnocanthus (Cottidae) в заливе Петра Великого и прилегающих районах Приморья // Вопр. ихтиологии. Т. 52. № 2. С. 234–247.
Панченко В.В. 2013. Размерно-возрастной состав и динамика численности нитчатого шлемоносца Gymnocanthus pistilliger (Cottidae) в заливе Петра Великого (Японское море) // Вопр. рыболовства. Т. 14. № 2 (54). С. 208–218.
Панченко В.В., Зуенко Ю.И. 2009. Распределение бычков семейства Cottidae в заливе Петра Великого Японского моря в летний период // Там же. Т. 10. № 4. С. 750–763.
Панченко В.В., Калчугин П.В., Соломатов С.Ф. 2016. Уточнение глубин обитания и максимальных размеров донных и придонных видов рыб в российских водах Японского моря // Вопр. ихтиологии. Т. 56. № 3. С. 264–283. https://doi.org/10.7868/S0042875216030152
Панченко В.В., Матвеев А.А., Панченко Л.Л. 2020. Сезонное распределение нитчатого шлемоносца Gymnocanthus pistilliger (Cottidae) в российских водах Японского моря // Там же. Т. 60. № 2. С. 174–182. https://doi.org/10.31857/S0042875220020174
Савин А.Б., Ильинский Е.И., Асеева Н.Л. 2011. Многолетняя динамика в составе донных и придонных рыб на западнокамчатском шельфе в 1982–2010 гг. // Изв. ТИНРО. Т. 166. С. 149–165.
Терентьев Д.А. 1998. Состав и биомасса донных рыб у западного побережья Камчатки по результатам учетной траловой съемки летом 1996 года // Исследования биологии и динамики численности промысловых рыб камчатского шельфа. Вып. IV. С. 28–32.
Токранов А.М. 1981а. Некоторые вопросы биологии нитчатого бычка Gymnacanthus pistilliger (Pallas) прикамчатских вод // Биологические ресурсы шельфа, их рациональное использование и охрана. Владивосток: Изд-во ДВНЦ АН СССР. С. 155–156.
Токранов А.М. 1981б. Распределение керчаковых (Cottidae, Pisces) на западнокамчатском шельфе в летний период // Зоол. журн. Т. 60. Вып. 2. С. 229–237.
Токранов А.М. 1985а. Питание рогатковых рода Gymnacanthus Swainson (Cottidae) прикамчатских вод // Вопр. ихтиологии. Т. 25. Вып. 3. С. 433–437.
Токранов А.М. 1985б. Изменение численности рогатковых (Cottidae) в прикамчатских водах // Тез. докл. Всесоюз. совещ. “Исследования и рациональное использование биоресурсов дальневосточных и северных морей СССР и перспективы создания технически средств для освоения неиспользуемых биоресурсов открытого океана”. Владивосток: Изд-во ТИНРО. С. 69.
Токранов А.М. 1987. О размножении рогатковых рыб рода Gymnocanthus (Cottidae) в прибрежных водах Камчатки // Вопр. ихтиологии. Т. 27. Вып. 6. С. 1026–1030.
Токранов А.М. 1988. Видовой состав и биомасса рогатковых (Pisces: Cottidae) в прибрежных водах Камчатки // Бюл. МОИП. Отд. биол. Т. 93. Вып. 4. С. 61–69.
Токранов А.М. 1993. О половом диморфизме массовых видов рогатковых (Cottidae) прикамчатских вод // Там же. Т. 98. Вып. 6. С. 19–26.
Токранов А.М. 2016. О половом диморфизме у рогатковых рыб (Cottidae, Pisces) прикамчатских вод // Матер. Междунар. конф. “XXX Любищевские чтения–2016. Современные проблемы эволюции и экологии”. Ульяновск: Изд-во УлГПУ. С. 124–131.
Токранов А.М. 2017. Рогатковые рыбы рода Gymnocanthus (Cottidae) прикамчатских вод и проблемы использования их ресурсов // Матер. VIII Всерос. науч.-практ. конф. “Природные ресурсы, их современное состояние, охрана, промысловое и техническое использование”. Ч. 1. Петропавловск-Камчатский: Изд-во КамчатГТУ. С. 176–180.
Токранов А.М., Орлов А.М. 2012. Особенности распределения и экологии двух видов рогатковых рыб рода Gymnocanthus (Cottidae) в тихоокеанских водах северных Курильских островов и юго-восточной Камчатки // Вопр. ихтиологии. Т. 52. № 6. С. 658–671.
Федоров В.В., Черешнев И.А., Назаркин М.В. и др. 2003. Каталог морских и пресноводных рыб северной части Охотского моря. Владивосток: Дальнаука, 204 с.
Фигуркин Л.А. 2002. Развитие океанологических условий западной Камчатки по данным мониторинговых наблюдений 1997 и 2000 гг. // Изв. ТИНРО. Т. 130. Ч. 1. С. 103–116.
Шелехов В.А., Панченко В.В. 2007. Возраст и рост нитчатого шлемоносца Gymnocanthus pistilliger (Cottidae) в водах южного Приморья (Японское море) // Вопр. ихтиологии. Т. 47. № 2. С. 228–237.
Шунтов В.П., Темных О.С. 2018. Среднемноголетняя биомасса и доминирующие виды рыб в донных и придонных биотопах охотского моря. 1. Состав и количественное соотношение видов на шельфе разных районов моря // Изв. ТИНРО. Т. 193. С. 3–19.https://doi.org/10.26428/1606-9919-2018-193-3-19
Allen M.L., Smith G.B. 1988. Atlas and zoogeography of common fishes in the Bering Sea and North-Eastern Pacific // US Dept. Comm. NOAA Tech. Rept. NMFS. № 66. 151 p.
Hoff J.R. 2000. Biology and ecology of threaded sculpin, Gymnocanthus pistilliger, in the eastern Bering Sea // Fish. Bull. V. 98. № 4. P. 711–722.
Mecklenburg C.W., Mecklenburg T.A., Thorsteinson L.K. 2002. Fishes of Alaska. Bethesda, Maryland: Amer. Fish. Soc., XXXVII + 1037 p. + 40 Pls.
Wilson D.E. 1973. Revision of the cottid genus Gymnocanthus, with a description of their osteology: PhD Thesis. Vancouver, Canada: Univ. British Columbia, 223 p.
Yamazaki A., Markevich A., Munehara H. 2013. Molecular phylogeny and zoogeography of marine sculpins in the genus Gymnocanthus (Teleostei; Cottidae) based on mitochondrial DNA sequences // Mar. Biol. V. 160. P. 2581–2589. https://doi.org/10.1007/s00227-013-2250-4
Дополнительные материалы отсутствуют.
Инструменты
Вопросы ихтиологии