

Вопросы ихтиологии, 2021, T. 61, № 4, стр. 384-392
Систематическое положение Dolichopteryx rostrata и ревизованная таблица для определения “длиннотелых” опистопроктид (Opisthoproctidae)
1 Институт проблем экологии и эволюции РАН – ИПЭЭ РАН
Москва, Россия
2 Институт океанологии РАН – ИО РАН
Москва, Россия
* E-mail: prokartster@gmail.com
Поступила в редакцию 12.11.2020
После доработки 10.12.2020
Принята к публикации 11.12.2020
Аннотация
Приведено переописание редкого вида Dolichopteryx rostrata, показана ошибочность сближения этого вида с другим неотеническим представителем опистопроктид – Duolentops minuscula. Несмотря на наличие ряда специфических для “длиннотелых” опистопроктид черт в строении глаз, пигментации и других особенностях, D. rostrata рассматривается в составе рода Dolichopteryx, общей эволюционной чертой которого считается телескопизация глаза и склеротизация его наружной оболочки между передним и задним выростами роговицы. В связи с изменением родовой принадлежности D. rostrata уточнён ключ для определения родов и видов “длиннотелых” опистопроктид.
Вид Dolichopteryx rostrata Fukui et Kitagawa, 2006 был описан по единственному экземпляру SL 66.2 мм, пойманному в Северной Атлантике к западу от Гебридских островов (Fukui, Kitagawa, 2006a). Позже Парин с соавторами (Parin et al., 2009) привели краткое описание еще четырёх экземпляров SL 28.0–52.0 мм из Центральной Атлантики между 10° и 16° с. ш. и сообщили о существовании материалов по этому виду в немецких коллекциях, собранных в районе 8° ю. ш. – 17° с. ш. и 14°–35° з. д. (сами рыбы изучены не были). Ревизуя роды “длиннотелых” опистопроктовых рыб, я отнёс этот вид к роду Duolentops Prokofiev, 2020 (типовой вид – Dolichopteryx minuscula Fukui et Kitagawa, 2006) (Прокофьев, 2020), основываясь исключительно на данных первоописания, так как материал по этому виду у меня отсутствовал. В первоописании наличествовали такие характерные общие признаки с типовым видом данного рода, как пузыревидные глаза с внутренней структурой линзовидной формы непосредственно под хрусталиком (Fukui, Kitagawa, 2006a. Fig. 1), устроенной точно так же, как на рисунке голотипа D. minuscula (Fukui, Kitagawa, 2006b. Fig. 1A); наличие “разлитой” меланофорной пигментации на хвостовом стебле, продолжающейся вперёд в виде двух супротивных полос меланофоров; неотения и близкие значения меристических признаков. Когда ревизия уже находилась в печати, мне посчастливилось обнаружить экземпляр длиннорылого долихоптерикса при разборе содержимого желудков алепизавров Alepisaurus ferox, добытых ярусным ловом в экваториальной Восточной Атлантике. Хотя идентификация этого экземпляра, имевшего очень длинное рыло и при SL 57.0 мм содержавшего зрелую икру, как D. rostrata казалась бесспорной, он имел ряд очень существенных отличий в строении глаз и пигментации тела в сравнении с первоописанием данного вида и определённо не мог быть помещён в род Duolentops. Благодаря любезности Дж. Маклейна (James Maclaine, Natural History Museum (BMNH), London), мне удалось ознакомиться с голотипом D. rostrata, после чего сомнения в конспецифичности моего экземпляра этому виду отпали. Оказалось, что в первоописании (Fukui, Kitagawa, 2006a) был допущен ряд ошибочных интерпретаций ключевых морфологических структур, что не позволяло правильно определить место этого вида в современной системе Opisthoproctidae. Настоящая статья посвящена переописанию D. rostrata и определению его реальной родовой принадлежности. Кроме того, поскольку в опубликованной мною определительной таблице (Прокофьев, 2020) этот вид был подчинён тезе 6(3), характеризующей род Duolentops, потребовалось переделать ключ для определения родов и видов “длиннотелых” опистопроктид.
МАТЕРИАЛ И МЕТОДИКА
Этикеточные данные экземпляров приведены при их описании. Методика изучения и терминология описаны в работе Прокофьева (2020). Следует отдельно остановиться на определениях “телескопического” и “полутелескопического” глаза. При телескопическом глазе роговица сильно выпуклая, зрачок и хрусталик имеют громадные размеры, причём хрусталик целиком охвачен зрачком (“вдвинут” в зрачок), а глазное яблоко имеет удлинённую форму, за счёт чего хрусталик в большей степени удалён от сетчатки, что позволяет распределять собираемые им световые лучи на максимально большей поверхности сетчатки (Brauer, 1908). Полутелескопический глаз, описываемый для D. rostrata, характеризуется пузыревидным (т.е. не удлинённым) глазным яблоком, но при этом выпуклой роговицей, смещённым и направленным вверх хрусталиком, чего не наблюдается в случае нетелескопических глаз (у Bathylychnops, Duolentops и Ioichthys), где хрусталик занимает центральное положение, а роговица плоская. В описании признаки голотипа, отличающиеся от таковых у нового экземпляра, приведены в скобках. В тексте используются следующие сокращения: D, A, P, V, С – соответственно спинной, анальный, грудные, брюшные и хвостовой плавники; sp. br – жаберные тычинки наружного ряда первой дуги, SL – стандартная длина, СРТМ – средний морозильный рыболовный траулер.
РЕЗУЛЬТАТЫ
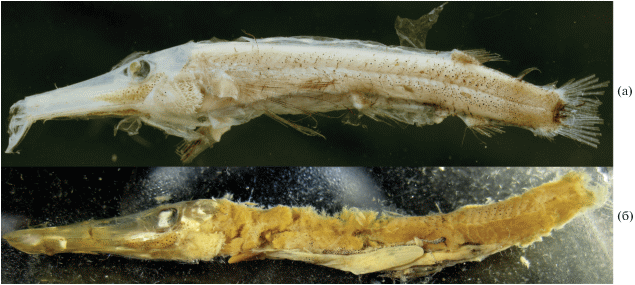
Dolichopteryx rostrata Fukui et Kitagawa, 2006 (рис. 1–5)
Материал. Голотип BMNH 2000.1.14.85, SL 66.2 мм, зрелая самка, 57°29′ с.ш. 11°32′ з.д., 600–0 м, 01.11.1999 (изучен по фотографиям в разных ракурсах). Нетиповой экземпляр ИО РАН б/№, SL 57 мм, зрелая самка, 01°58′ ю.ш., 07°31′ з. д., из желудка Alepisaurus ferox SL 1090 мм, СРТМ-8018, ярус № 2, 24.12.1971 г.
Диагноз. Неотенический вид рода Dolichopteryx (максимальная известная SL 66.2 мм) с полутелескопическими глазами (глазное яблоко пузыревидное при выпуклой роговице и смещённом в зрачок хрусталике) без афокальной зоны, с бобовидными одноразмерными передним (шаровидное тело) и задним выростами роговицы, без дополнительного хрусталика и субсклеральных линз, но с обширной областью склеротизации наружной оболочки роговицы под хрусталиком, с полулунным участком белой (вероятно, светящейся) ткани по нижнему краю орбиты; с длинным рылом (19.6–25.4% SL); с широким скоплением крупных меланофоров под глазом, имеющим форму запятой, продолжающимся вперёд на боковую поверхность рыла; со сплошным скоплением тесно сидящих мелких меланофоров на хвостовом стебле, резко отличных от рассеянных крупных меланофоров в предшествующих отделах тела; с длинным тёмно окрашенным Р и 40–42 позвонками.
Описание. Все плавники сильно повреждены, в D не менее восьми лучей, Р 14, в остальных плавниках даже приблизительный подсчет невозможен ввиду утраты части лучей (у голотипа D 9, A 8, P 14, V 12, C vii + 10 + 9 + ix). В поперечном ряду на уровне начала D у голотипа около семи чешуй. Позвонков (миомеров) 42 (41). Рыло очень длинное (рис. 1), в 1.63 (1.95) раза превышает суммарную длину орбиты и посторбитальной части головы. Край frontale над орбитой образует пластинчатый выступ. Вершина рыла отчётливо выступает перед нижней челюстью (у голотипа вершина рыла сломана). Зубы на vomer мелкие, игловидные, расположены полоской до пяти рядов в ширину, на dentale – ещё мельче, щетинковидные, однорядные. Жаберные тычинки (рис. 2, 3) расположены двумя разделёнными рядами на epi- и ceratobranchiale жаберных дуг. Поверхность жаберных дуг между тычинками покрыта густой массой мукуса. В наружном ряду на первой жаберной дуге 3 + 7 крупных уплощённых тесно сидящих тычинок11, в своём расширенном основании имеющих хрящевую центральную пластинку. Вершины тычинок вытянуты в узкие отростки, загнутые на конце (рис. 2а, 3а). Во внутреннем ряду имеются очень мелкие, трудно различимые тычинки, имеющие вид коротких филаментов или бугорков и расположенные широко разобщёнными группами (1 + 4 на epibranchiale-1, 1 на сочленении epi- и ceratobranchiale, 2 + 1 + 3 на ceratobranchiale-1); у отдельных наиболее развитых из них в основании прослеживается хрящевый центр (рис. 2б–2г, 3б). На hypobranchiale-1 тычинки не обнаружены. На последующих жаберных дугах тычинки наружного ряда мельче, а во внутреннем, напротив, лучше развиты. В наружном ряду на второй жаберной дуге две бугорковидные и одна уплощённая и суженная к вершине (язычковидная) тычинка, на ceratobranchiale-2 четыре язычковидные тычинки с оттянутыми и загнутыми на концах вершинами; на hypobranchiale-2 две бугорковидные тычинки (рис. 2д). Во внутреннем ряду на второй дуге 4 + 8 мелких тычинок (верхние бугорковидные, нижние – короткие язычковидные) (рис. 2е).
Рис. 2.
Dolichopteryx rostrata, экземпляр SL 57 мм, жаберные тычинки первой (а–г) и второй (д, е) дуг: а – наружная сторона; б – внутренняя сторона; в, г – группа тычинок на внутренней стороне нижней части ceratobranchiale-1; д – вид дорсолатерально, е – внутренняя сторона (применён фильтр смещения баланса цвета для лучшей визуализации тычинок). cbri, ebri – соответственно жаберные тычинки внутреннего ряда на epi- и ceratobranchiale второй дуги; cc – хрящевой центр тычинки, hbr – тычинки на hypobranchiale-2, muc – скопления мукуса на жаберных дугах, spbi – тычинки самой нижней группы внутреннего ряда на ceratobranchiale-1. Масштаб, мм: а, б, д – 1; в – 0.5; г, е – 0.25.
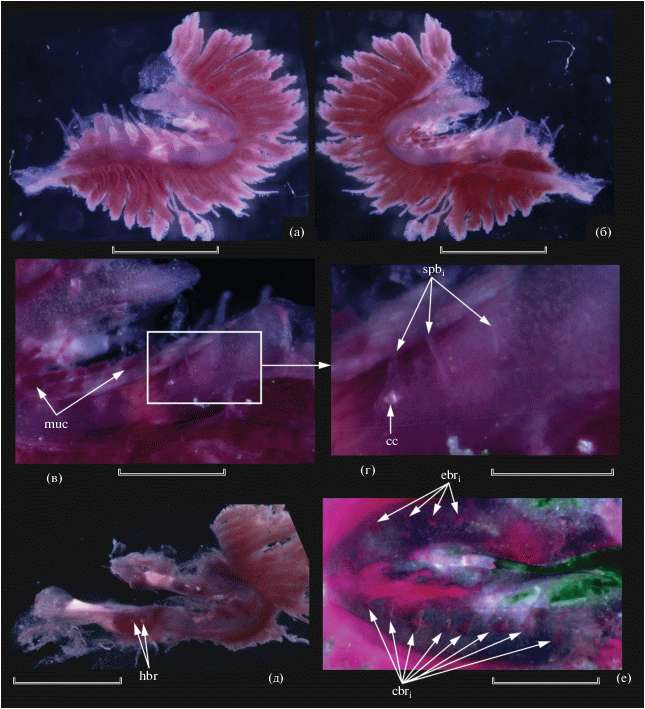
Рис. 3.
Dolichopteryx rostrata, тычинки первой жаберной дуги: а, б – экз. SL 57 мм, схема расположения (а – наружный ряд, б – внутренний ряд); в – голотип, тычинки наружного ряда. Масштаб: 0.5 мм.
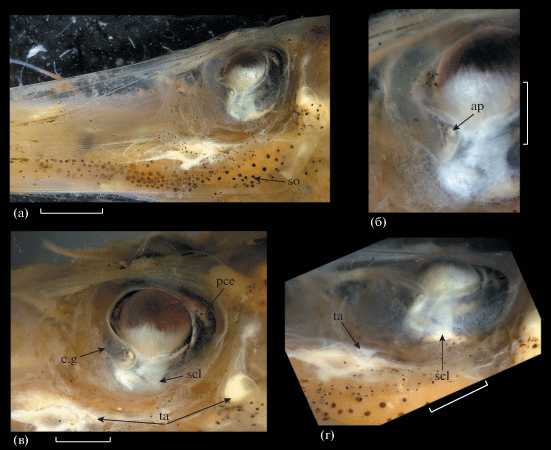
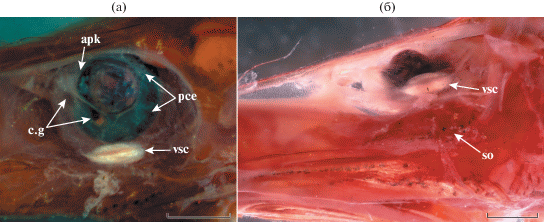
Глаз полутелескопический – глазное яблоко пузыревидное, но роговица сильно выпуклая, хрусталик “вдвинут” в зрачок и направлен вверх; афокальная зона перед хрусталиком отсутствует (рис. 4). Передний вырост роговицы (шаровидное тело) бобовидный, расположен по переднему краю хрусталика, с хорошо выраженным “зеркальцем” (рис. 4б, 4в). Задний вырост роговицы по форме и величине сопоставим с передним (рис. 4в). Наружная оболочка глаза в вентральной части (под видимой снаружи частью хрусталика) с обширным полем наружной склеротизации (между вентральными концами переднего и заднего выростов); другие склеротизированные структуры отсутствуют. У вентрального края орбиты имеется полоска легко отделяющейся молочно-белой (светящейся?) ткани (у голотипа она не определяется, возможно, сорвана) (рис. 4в, 4г).
Рис. 4.
Глаз Dolichopteryx rostrata, экземпляр SL 57 мм: а – общий вид и подглазничное скопление меланофоров; б – передний вырост роговицы, в – вид сбоку, г – вид снизу. ap – “зеркальце”, c.g – передний вырост роговицы, pce – задний вырост роговицы, scl – склеротизированный участок наружной оболочки глаза, so – подглазничное скопление меланофоров, ta – белая (светящаяся?) ткань по нижнему краю орбиты. Масштаб, мм: а – 2, б–г – 1.
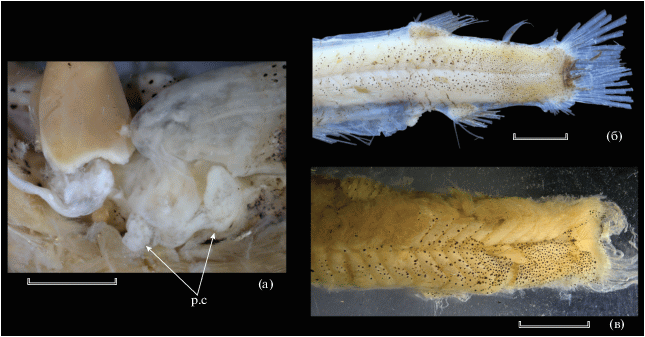
Парные плавники в первоописании изображены одинаково длинными (Fukui, Kitagawa, 2006a. Fig. 1). В настоящее время V у голотипа сильно обломаны; концы лучей верхней половины Р также обломаны, наибольшие лучи в нижней половине достигают начала V. V прикрепляются далеко впереди вертикали начала D (у голотипа – на 10 миомеров), на равном расстоянии от начал Р и А. Начало A расположено за вертикалью конца основания D22. Жировой плавник у экз. SL 57 мм утрачен (у голотипа хорошо развит, прикрепляется позади вертикали конца основания А). Пилорических придатков два, редуцированы до небольших бугорков (рис. 5а). Как и голотип, экземпляр SL 57 мм является самкой со зрелой икрой, с хорошо отделяющимися отдельными икринками, диаметром около 0.8 мм.
Рис. 5.
Пилорические придатки (p.c) (а) и пигментация задней половины тела (б, в) Dolichopteryx rostratа: а, в – экземпляр SL 57 мм; б –голотип. Масштаб, мм: а – 1; б, в – 3.
Измерения экземпляра SL 57 мм, в % SL: длина головы 36.8, длина рыла 22.8, горизонтальный диаметр орбиты 6.6, длина нижней челюсти 7.0, максимальная и минимальная высота тела соответственно 8.8 и 7.0, предорсальное и преанальное расстояния соответственно 77.2 и 82.5.
Пигментация. Мелкоточечная меланофорная пигментация на нижней челюсти, по боковому краю рыла и вдоль костей подвеска, в заглазничной области; в подглазничной области (рис. 4а) обширное скопление в форме запятой, состоящее из довольно крупных тесно расположенных меланофоров, протягивающееся вперёд до середины длины рыла. Участка рефлектирующей ткани под глазом нет. Довольно плотное скопление более мелких меланофоров в гулярной области на уровне глаза. Жаберные дуги не пигментированы. На теле меланофорная пигментация, представленная разноразмерными, в целом довольно крупными меланофорами, до вертикали основания V концентрирующимися вдоль краёв миосепт, далее назад – по всей поверхности миомеров. На хвостовом стебле очень плотная сплошная мелкоточечная меланофорная пигментация по всей поверхности. Пятен серебристой ткани или рефлектирующих участков на коже нет (рис. 5б, 5в). Срединная дорсальная миосепта не пигментирована. У голотипа сохранились остатки тёмноокрашенных чешуйных карманов. Лучи (и перепонка?) Р пигментированы, остальных плавников, по-видимому, нет. В основаниях D, A и Р развит мелкий меланофорный крап. Перитонеум сплошь и густо пигментирован крупными раздельными меланофорами; перитонеальных пятен нет. Желудочно-кишечный тракт не пигментирован.
ОБСУЖДЕНИЕ
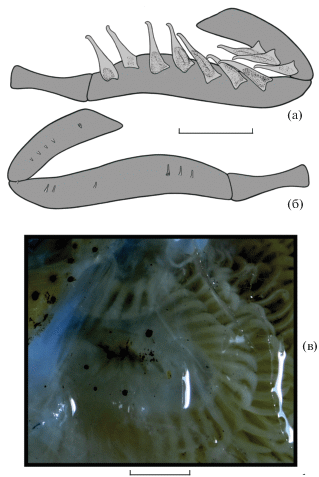
Dolichopteryx rostrata выделяется среди всех представителей группы “длиннотелых” опистопроктид рядом специфических особенностей, а именно строением глаза, сочетающим в себе ряд примитивных и продвинутых черт, исключительно длинным рылом, редукцией пилорических придатков и специфической пигментацией. Среди перечисленных признаков филогенетически наиболее значимыми представляются особенности строения глаза, по которым данный вид не может быть однозначно отнесён к какому-либо из признаваемых в семействе родов. Несмотря на ряд очень примитивных черт, таких как пузыревидное глазное яблоко со слабо развитыми (бобовидными) выростами роговицы, как у наиболее примитивных в этом отношении родов Ioichthys и Duolentops, роговица у этого вида сильно выпуклая и наблюдается смещение хрусталика в зрачок, подобно Dolichopteryx и Dolichopteroides (рис. 4в). Необычной чертой глаза D. rostrata, ранее для “длиннотелых” опистопроктид не отмечавшейся, является наличие под видимой частью хрусталика обширного поля поверхностной склеротизации, соединяющего вентральные концы переднего и заднего выроста роговицы. Гомология этого образования субсклеральной линзе Duolentops (рис. 6) неочевидна, однако оно определенно гомологично утолщению склеры между выростами роговицы у Dolichopteryx (Прокофьев, 2020. Рис. 1е, 1ж), хотя и занимает много бóльшую площадь. Впрочем, последняя особенность может объясняться слабым развитием роговичных выростов, которые у прочих Dolichopteryx сближены и задний из них смещён под передний (Прокофьев, 2020). Наличие у D. rostrata участка молочно-белой поверхностно расположенной ткани по вентральному краю орбиты сближает его с D. anascopa и D. parini. Другие черты сходства с этими видами включают длинные парные плавники (но начало А расположено за концом основания D, а не на его середине, как у D. anascopa и D. parini), наличие жирового плавника и отсутствие перитонеальных пятен (два последних признака, впрочем, отмечены и у некоторых других видов рода Dolichopteryx (Прокофьев, 2020)).
Рис. 6.
Duolentops minuscula SL 30 мм (23°34′ с. ш. 128°35′ в. д., 500 м): а – глаз сбоку; б – глаз и подглазничное скопление меланофоров вентролатерально. apk – афокальное пространство, vsc – субсклеральная линза, ост. обозначения см. на рис. 4. Масштаб: 0.75 мм.
Характер пигментации тела у D. rostrata не имеет аналогов в пределах группы. Он определенно не сходен с пигментацией Duolentops minuscula, несмотря на то что в оригинальных описаниях этих видов (Fukui, Kitagawa, 2006a, 2006b) пигментация изображена очень похоже. Хвостовой стебель D. minuscula несёт разлитую сливающуюся мелкодисперсную пигментацию, далее вперёд распадающуюся на две нечёткие супротивные продольные полосы; эпидермис выше и ниже миокоммы с рефлектирующими серебристыми участками (Прокофьев, 2020. Рис. 5а). Наличие двух супротивных пигментных полос является характерной чертой молоди Dolichopteryx и Dolichopteroides (серебристые рефлектирующие участки у неё либо отсутствуют, либо оформлены в виде чётко отграниченных серебристых пятен, обычно окаймлённых меланофорной пигментацией). У взрослых Dolichopteryx и Dolichopteroides подкожная пигментация отсутствует. Напротив, у D. rostrata подкожная пигментация очень похожа на таковую у молоди Ioichthys, но резко отличается плотным скоплением мелких меланофоров на хвостовом стебле, совершенно не схожих с меланофорами на предшествующих участках тела (рис. 5б, 5в). Вопреки рисунку в первоописании (Fukui, Kitagawa, 2006a. Fig. 1), разобщённые супротивные полосы меланофоров на боках тела у D. rostrata отсутствуют.
Поскольку сходство D. rostrata с D. minuscula в строении глаза ограничивается только двумя плезиоморфиями (пузыревидная форма глазного яблока и слабое развитие роговичных выростов) и совершенно не включает апоморфные признаки (обособленные субсклеральные линзы у Duolentops, выпуклость роговицы, смещение хрусталика и наличие склеротизации наружной оболочки глаза между роговичными выростами у D. rostrata), и кроме того, D. rostrata лишён каких-либо других апоморфий D. minuscula (в строении жаберных тычинок (Прокофьев, 2020. Рис. 3) и пигментации), отнесение D. rostrata к роду Duolentops не может быть принято. Помимо неотении (которая вполне могла возникнуть независимо), единственной необычной чертой сходства между D. rostrata и D. minuscula является похожая форма пилорических придатков, при том что последние у D. rostrata сильно редуцированы (в отличие от D. minuscula: Прокофьев, 2020. Рис. 5в). На мой взгляд, это сходство может быть случайным.
Гораздо больше оснований сближать D. rostrata с Dolichopteryx, а примитивные по сравнению с другими видами данного рода черты строения глаза расценивать как рекапитуляцию к предковому состоянию. Во всяком случае в строении глаза D. rostrata прослеживается характерное для Dolichopteryx направление специализации: выпуклость роговицы и смещение хрусталика, приводящие в итоге к телескопическому глазу, и развитие наружной склеротизации между роговичными выростами при отсутствии каких-либо обособленных склеротизаций, которые могли бы выполнять роль “дополнительного хрусталика”, свойственных родам Bathylychnops и Duolentops. Эту рекапитуляцию нельзя связывать с неотенизацией, поскольку у молоди других Dolichopteryx все характерные черты строения глаза уже сформированы при SL от 20 мм (Прокофьев, 2020).
Конечно, нельзя полностью отвергать возможность выделения D. rostrata в особый род, учитывая его своеобразие в пределах группы “длиннотелых” родов. Однако на настоящем уровне изученности данной группы это представляется преждевременным. Как отмечалось ранее (Прокофьев, 2020), род Dolichopteryx не вполне однороден. Виды D. anascopa и D. parini заметно отличаются от остальных представителей рода по наличию участка поверхностной белой (светящейся?) ткани по нижнему краю орбиты, одинаково длинных парных плавников и взаиморасположению вертикальных плавников, причём по первым двум признакам D. rostrata может быть сближен с этими видами. В составе рода также может быть выделена группа видов “longipes” (D. andriashevi, D. longipes, D. nigripes и D. vityazi), однако другие виды (D. pseudolongipes, D. trunovi) занимают промежуточное положение между этими двумя группами. Поэтому какие-либо таксономические выводы представляются уместными только после поступления дополнительных материалов, которые позволят охарактеризовать онтогенетическую изменчивость для всех видов рода и провести их детальное анатомирование.
Таблица для определения родов и видов “длиннотелых” опистопроктид
1(2) Глаза пузыревидные, без обособленного шаровидного тела и дополнительного хрусталика; хрусталик занимает центральное положение; роговица плоская, афокальная зона имеется; нет склеротизации наружной оболочки глаза между передним и задним роговичными выростами (рис. 6) …………………………............................…..… 3
2(1) Глаза телескопические, а если пузыревидные, то с крупным шаровидным телом, содержащим дополнительный хрусталик, либо с выпуклой роговицей и смещённым хрусталиком, в последнем случае имеется участок склеротизации наружной оболочки глаза между передним и задним роговичными выростами (рис. 4); афокальной зоны нет ……....…………... 7
3(6) Позвонков 53–58; субсклеральное линзовидное уплотнение под хрусталиком отсутствует; жаберные тычинки в два ряда на epi- и ceratobranchiale первой дуги; в наружном ряду удлинённые у молоди и короткие у взрослых рыб, не гипертрофированы; сошник без зубов; краевых лучей С 5–6; SL до 253 мм … (Ioichthys Parin, 2004) ……...........................…………………… 4
4(5) Позвонков 53–55; Индийский океан………………………………...… I. kashkini Parin, 2004
5(4) Позвонков 56–58; Восточная Пацифика……………………………..……….........…… Ioichthys sp.
6(3) Позвонков 45–46; имеется чётко отграниченное субсклеральное линзовидное уплотнение под хрусталиком (рис. 6); жаберные тычинки на первой дуге утолщённые, пальцевидные, в наружном ряду сильно гипертрофированы; две верхние тычинки наружного ряда прикреплены медиальнее линии, соединяющей основания последующих тычинок на ceratobranchiale-1, и наклонены вниз параллельно продольной оси ceratobranchiale-1, лежа позади наружного ряда цератобранхиальных тычинок (Прокофьев, 2020. Рис. 3); передний край сошника с многорядными зубами; краевых лучей С 9–11; неотеническая форма, максимальная известная SL ~ 60 мм … (Duolentops Prokofiev, 2020) ....................................................... .................... D. minuscula (Fukui et Kitagawa, 2006)
7(8) Глаза пузыревидные, с хорошо обособленным шаровидным телом, содержащим дополнительный хрусталик; у рыб SL >112 мм имеются две корнеальные линзы позади шаровидного тела; хрусталик занимает центральное положение; позвонков 67–85; зубы на сошнике однорядные; мальки имеют продольный ряд пятен по дорсальному краю тела … (Bathylychnops Cohen, 1958)....….……..9
8(7) Глаза телескопические, с хорошо обособленным шаровидным телом, но без дополнительного хрусталика33 и без корнеальных линз близ нижнего и нижнезаднего края глаза; если пузыревидные, то передний вырост роговицы бобовидный, не образует обособленного шаровидного тела, а хрусталик смещён и направлен вверх; позвонков 41–60; озубление сошника многорядное; нет дорсальных пигментных пятен у молоди …………13
9(10) Позвонков 65–73 …................................... ................................... B. brachyrhynchus (Parr, 1937)
10(9) Позвонков 77–85 ..………………………..... 11
11(12) V прикрепляются далеко впереди вертикали начала D; жировой плавник расположен над концом А; Юго-Восточная Пацифика ..............…... .......... B. chilensis Parin, Belyanina et Evseenko, 2009
12(11) V и D начинаются близ одной вертикали; жировой плавник расположен над передней третью или серединой А; Северная Пацифика…..........…………….……………. B. exilis Cohen, 1958
13(14) Позвонков 58–60; основания и V, и A хотя бы частично располагаются под основанием D; Р значительно длиннее V … (Dolichopteroides Parin, Belyanina et Evseenko, 2009) ….……………................ ................. Dolichopteroides binocularis (Beebe, 1932)
14(13) Позвонков 40–48; основание V всегда расположено заметно впереди вертикали начала D; Р значительно короче V или примерно одинаковой длины с ними … (Dolichopteryx Brauer, 1901) …………………..…………………………….…….…………. 15
15(16) Глазное яблоко пузыревидное (глаз полутелескопический), передний вырост роговицы бобовидный, не образует обособленного шаровидного тела; рыло длинное (19.6–25.4% SL); подглазничное скопление меланофоров продолжается по боковой поверхности рыла до середины его длины; меланофоры на теле не образуют двух продольных супротивных полос выше и ниже миокоммы; неотеническая форма ……..………. ........................... D. rostrata Fukui et Kitagawa, 2006
16(15) Глазное яблоко цилиндрическое (глаз телескопический), передний вырост роговицы образует обособленное шаровидное тело; рыло короче, 10.6–17.5% SL; у мальков подглазничное скопление меланофоров не продолжается на боковую поверхность рыла, меланофоры на теле формируют две продольные супротивные полосы выше и ниже миокоммы ………………………………17
17(18) Жировой плавник имеется; перитонеальных пятен у молоди нет (кроме D. vityazi; не известно для D. parini)……………….…….……………… 19
18(17) Жировой плавник отсутствует; перитонеальные пятна у молоди имеются .………....……27
19(20) Р и V сильно удлинённые, начало А расположено на вертикали середины длины основания D, у нижнего края глаза имеется участок молочно-белой (не склеротизированной) ткани…..………………………………………..………………… 21
20(19) Р много короче V, начало А расположено перед концом основания D или позади него, нет специфической ткани у нижнего края глаза …………..…………………….………….......................… 23
21(22) Позвонков (миомеров) 41–44, V 12; P, V и D не пигментированы; крупный овальный участок белой ткани у нижнего края глаза; тропическая Индо-Вест-Пацификa..…. D. anascopa Brauer, 1901
22(21) Позвонков (миомеров) 46–47, V 10–11; P, V и D пигментированы; узкий полулунный участок белой ткани у нижнего края глаза; переходная зона Северной Пацифики …………................... ..................... D. parini Kobyliansky et Fedorov, 2001
23(24) Перитонеальные пятна у молоди имеются [sp. br 23–28; тропическая Индо-Вест-Пацифика]...……D. vityazi Parin, Belyanina et Evseenko, 2009
24(23) Перитонеальных пятен у молоди нет ..........................................................................… 25
25(26) sp. br 31–35; Калифорнийский переходный район и экваториальная часть Тихого океана от Новой Каледонии до о-вов Галапагос ……… .......D. pseudolongipes Fukui, Kitagawa et Parin, 2008
26(25) sp. br 11–15; воды Южного полушария между 41° и 53° ю. ш. …......… D. trunovi Parin, 2005
27(28) Четыре перитонеальных пятна в промежутке P–V; жаберные дуги с точечной меланофорной пигментацией [V не окрашены; начало A под концом D] …………………..……............................ ..... D. andriashevi Parin, Belyanina et Evseenko, 2009
28(27) Три перитонеальных пятна в промежутке P–V; жаберные дуги не пигментированы ….. 29
29(30) Начало А немного впереди вертикали конца D; перепонка между третьим–седьмым лучами V интенсивно-чёрная (кроме самого основания); тропическая южная часть Тихого океана ………………………….……... D. nigripes Prokofiev, 2020
30(29) Начало А позади вертикали конца D; V не пигментированы; Атлантический океан …………………..………….…. D. longipes (Vaillant, 1888)
Список литературы
Прокофьев А.М. 2020. Пересмотр родовой классификации “длиннотелых” опистопроктовых рыб (Opisthoproctidae) с описанием новых таксонов и новых находок // Вопр. ихтиологии. Т. 60. № 5. С. 515–543. https://doi.org/10.31857/S0042875220050100
Brauer A. 1908. Die Tiefsee-Fische. II. Anatomischer Teil // Wissenschaftliche Ergebnisse der Deutschen Tiefsee-Expedition auf dem Dampfer “Valdivia” 1898–1899. Bd. 15. Jena: Verlag von Gustav Fischer, 266 S., Taf. XIX–XLIV.
Fukui A., Kitagawa Y. 2006a. Dolichopteryx rostrata, a new species of spookfish (Argentinoidea: Opisthoproctidae) from the eastern North Atlantic Ocean // Ichthyol. Res. V. 53. № 1. P. 7–12. https://doi.org/10.1007/s10228-005-0306-2
Fukui A., Kitagawa Y. 2006b. Dolichopteryx minuscula, a new species of spookfish (Argentinoidei: Opisthoproctidae) from the Indo-West Pacific // Ibid. V. 53. № 2. P. 113–120. https://doi.org/10.1007/s10228-005-0329-8
Parin N.V., Belyanina T.N., Evseenko S.A. 2009. Materials to the revision of the genus Dolichopteryx and closely related taxa (Ioichthys, Bathylychnops) with the separation of a new genus Dolichopteroides and description of three new species (fam. Opisthoproctidae) // J. Ichthyol. V. 49. № 10. P. 839–851. https://doi.org/10.1134/S0032945209100014
Дополнительные материалы отсутствуют.
Инструменты
Вопросы ихтиологии