

Вопросы ихтиологии, 2021, T. 61, № 5, стр. 591-598
Морфологические изменения эпидермальных клеток у рыб с хронической аносмией и энуклеацией
Г. В. Девицина *
Московский государственный университет
Москва, Россия
* E-mail: gdevicyna@mail.ru
Поступила в редакцию 13.07.2020
После доработки 30.07.2020
Принята к публикации 30.07.2020
Аннотация
С помощью сканирующей электронной микроскопии изучена структура апикальной поверхности эпидермальных клеток в коже головы и в эпителии ротовой полости особей карпа Cyprinus carpio − интактных и с хронической ольфакторной и зрительной деафферентацией. Показано, что сенсорно депривированные рыбы отличаются от интактных значительными изменениями в морфологии апикальной поверхности эпидермальных клеток. На апикальной поверхности клеток нарушается правильный кольцевидный рисунок, но появляются сложные фигуры или пластинчатые разрастания кольцевидных гребней. Характер расположения и ориентации, а также плотность распределения и форма рисунка на апикальной поверхности даже соседних клеток могут быть совершенно различными. Высокая концентрация клеток с модифицированной апикальной поверхностью отмечена в сенсорных зонах вкусовой и тактильной рецепции. Изменения имеют сходный характер у эпидермальных клеток кожи и клеток эпителия ротовой полости. Индивидуальная специфика структурных изменений эпидермальных клеток не выявлена. Обсуждается возможная взаимосвязь изменений апикальной поверхности эпидермальных клеток с активацией системы кожной рецепции и других сенсорных систем.
Разные сенсорные системы рыб функционируют в комплексе, обеспечивая полисенсорность различных форм поведения (Павлов, Касумян, 1990; Devitsina, 2003). Однако в силу определённых условий существования функция отдельных систем может быть ограничена или утрачена. В таких случаях оставшиеся интактными сенсорные структуры могут проявлять признаки морфофункциональной активации, направленной на компенсацию дефицита информации (Девицина, Марусов, 2007; Касумян, Марусов, 2007). Хроническое выключение сенсорных систем дистантного действия (зрительной и обонятельной) у рыб разных видов приводит к сходной адаптивной реакции вкусовой системы. Вкусовые рецепторы у рыб могут располагаться не только в эпителии ротовой полости, но и в наружных покровах головы и тела. Адаптивная реакция выражается в гипертрофии вкусового рецепторного аппарата и активации нейронов вкусовых центров мозга (Девицина, 2006; Девицина и др., 2010, 2015).
Можно полагать, что потеря важнейших для поведения сенсорных каналов дистантного действия не всегда может полностью компенсироваться активацией только одной системой контактного действия – вкусовой. У рыб одним из хорошо развитых сенсорных каналов контактного действия является тактильная рецепция, которая играет большую роль в регуляции поведения многих видов (Kasumyan, 2011). В коже головы карповых рыб (Cyprinidae) обнаружена высокая тактильная чувствительность, которая представлена рецепторами тонального и фазного типов (Девицина, Лапшин, 2016). Тактильная чувствительность на латеральной, дорсальной и вентральной стороне головы карпа Cyprinus carpio распределяется не равномерно, а по зонам. Наиболее чувствительные зоны находятся около ротовой полости. Уровень тактильной чувствительности на голове карпа снижается в рострокаудальном и в вентродорсальном направлении, что имеет определённую биологическую значимость (Девицина, Лапшин, 2020). В связи с этим особый интерес представляет изучение морфологии апикальной поверхности клеток кожи на голове карпа.
У рыб тактильная рецепция совместно с другими видами кожной рецепции образует сложный осязательный комплекс, или сомато-висцеральную систему (Kasumyan, 2011). Общее свойство рецепторов сомато-висцеральной системы рыб заключается в том, что они не являются элементами сенсорных органов, а рассредоточены диффузно по всему телу. Восприятие внешних механических воздействий происходит через непосредственный контакт кожи с внешней средой (Windsor et al., 2008). Структурами, воспринимающими внешние стимулы, являются клетки поверхностного слоя эпидермиса. Поверхностные эпидермальные клетки позвоночных поляризованы, их апикальная часть морфологически и функционально отличается от базальной части (Елисеев и др., 1972). Через апикальную поверхность эпидермальных клеток идёт восприятие внешних стимулов, в том числе и механических.
Специфика кожного канала сенсорной информации, находящегося на границе организма с внешней средой, как у наземных позвоночных, так и у рыб связана и с защитной функцией (Ильинский, 1967; Трубачёв, Василевский, 1976; Kasumyan, 2011). Вопрос о том, какие именно структуры кожи связаны с восприятием тех или иных видов механических воздействий, остаётся пока ещё недостаточно выясненным. Морфологические и функциональные особенности кожной механорецепции и возможности её изменчивости, в том числе в результате полисенсорного дефицита, являются мало исследованным разделом проблемы сенсорных адаптаций.
Цель настоящей работы − изучить влияния хронического выключения сенсорных систем дистантного действия (зрительной и обонятельной) на морфологические особенности апикальной поверхности эпидермальных клеток в тактильно чувствительных зонах, в зонах вкусовой рецепции на поверхности головы и в выстилке ротовой полости рыб.
МАТЕРИАЛ И МЕТОДИКА
Объектом исследования являлись семь особей карпа − (четыре опытные и три контрольные), которые в начале эксперимента имели длину 10–12 см. Опытных рыб подвергли двухсторонней аносмии с помощью термокаутера, а спустя 5 сут − двусторонней энуклеации через небольшой надрез роговицы. Опытные рыбы получили анестезию с гипотермией и легко перенесли обе операции: через 1 сут после операции они свободно плавали в аквариуме в поисках корма. Оперированных и интактных рыб содержали в одинаковых условиях при температуре 15−17°С и кормили личинками комаров (Chironomidae).
Через 7 мес. после операций фиксировали образцы кожи, взятые с одинаковых участков головы у каждой из интактных и опытных рыб − это кожа на латеральной стороне головы между органом обоняния и верхней губой, на верхней губе в её латеральной и в центральной частях, а также на малом усике. На вентральной стороне головы образцы кожи брали в центральной части нижней губы и в гулярной зоне рядом с нижней губой. Всего исследовали 25 образцов кожи от контрольных рыб и 33 от опытных рыб. В ротовой полости образцы эпителия брали в максиллярной и в мандибулярной зонах. У контрольных рыб взяли девять образцов эпителия, у опытных – 12.
Образцы фиксировали 2.5%-ным раствором глютарого альдегида на 0.1 М фосфатном буфере (рН 7.4). Отмывку фиксатора проводили тем же буфером с добавлением сахарозы. Затем образцы обезвоживали в батарее этанол–ацетон и высушивали на аппарате Hitachi HCP-1. Методика обработки описана ранее (Девицина, 1990). Образцы, наклеенные на латунные столики, напыляли смесью золота с палладием. Морфологию апикальной поверхности эпидермальных клеток изучали с помощью электронных сканирующих микроскопов CamScan S2 или JSM-6380LA.
РЕЗУЛЬТАТЫ
Интактные рыбы. На поверхности кожи головы карпа в выделенных ранее сенсорных зонах по критерию тактильной чувствительности (Девицина, Лапшин, 2020) эпидермальные клетки имеют одинаковый характерный кольцевидный рисунок, образованный рёбрами на их апикальной поверхности (рис. 1). Характерный кольцевидный рисунок на поверхности клеток присутствует в эпителии губ, в коже каудальнее губ, на челюстном усике, в том числе вблизи пор вкусовых почек (рис. 1а, 1б, 1г). На поверхности эпителия околоротовых зон видны вершины многочисленных одиночных хемосенсорных клеток.
Рис. 1.
Апикальная поверхность эпидермальных клеток у интактных особей карпа Cyprinus carpio: а – между верхней губой и органом обоняния, б – около вкусовой почки в средней части малого усика, в – около вкусовой почки в максиллярной зоне ротовой полости, г − на каудальном краю верхней губы. 1 – микровиллы одиночных хемосенсорных клеток, 2 – обычный кольцевидный рисунок, 3 – вкусовая почка. Масштаб, мкм: а – 30, б – 2, в – 20, г – 10.
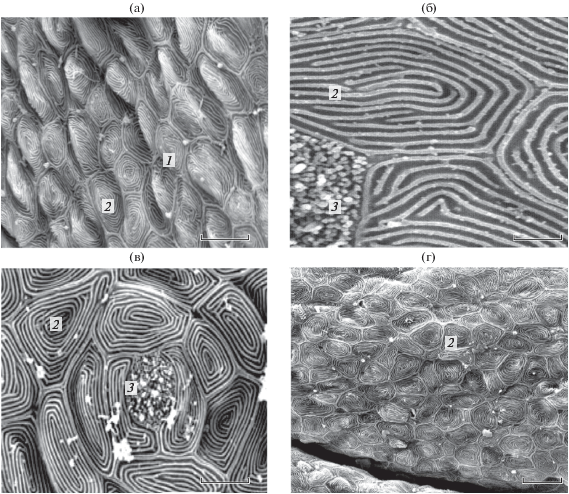
В ротовой полости карпа эпидермальные клетки максиллярной и мандибулярной зон образуют небольшие возвышения и окружают вкусовые почки. Апикальная поверхность эпидермальных клеток, расположенных вокруг вкусовых почек и между ними, имеет хорошо выраженный характерный кольцевидный рисунок − такой же, как у клеток снаружи (рис. 1в).
Сенсорно депривированные рыбы. В коже головы карпа на апикальной поверхности эпидермальных клеток кольцевидный рисунок нарушен. От кольцевидных гребней сохраняются многочисленные фрагменты разной длины и формы. В целом на апикальной поверхности клеток образуется сложный и варьирующий по форме рисунок (рис. 2). Расположение и ориентация, а также плотность распределения, форма и размеры фрагментов гребней, даже у соседних клеток, могут быть совершенно различными (рис. 2а). Наиболее распространённые формы модификации апикальной поверхности клеток: фрагментарный и точечный рисунки (рис. 2б, 2г), пластинчатые разрастания (рис. 3а, 3б).
Рис. 2.
Разнообразие морфологических изменений апикальной поверхности эпидермальных клеток во вкусовых зонах головы карпа Cyprinus carpio с хронической ольфакторной и зрительной деафферентацией: а – четыре эпидермальные клетки с разным рисунком апикальной поверхности в коже верхней губы; б – вкусовая почка на верхней губе, окружённая эпидермальными клетками с модифицированной апикальной поверхностью; в – вкусовая почка в максиллярной зоне ротовой полости, окружённая клетками с модифицированной апикальной поверхностью; г – вкусовая почка в базальной части малого усика, окружённая эпидермальными клетками с модифицированной апикальной поверхностью. 4 – точечный рисунок, 5 – фрагментарный рисунок, ост. обозначения см. на рис. 1. Масштаб, мкм: а – 5, б – 20, в – 3, г – 10.

Рис. 3.
Модифицированная апикальная поверхность эпидермальных клеток кожи в тактильно чувствительных зонах головы карпа Cyprinus carpio с хронической ольфакторной и зрительной деафферентацией: а – клетки с кольцевидными разрастаниями гребней и с изменением формы апикальной поверхности на латеральной стороне головы каудальнее верхней губы, б – апикальная поверхность эпидермальных клеток с пластинчатыми кольцевидными разрастаниями, в – клетки с различными модификациями апикальной поверхности на вентральной стороне головы в гулярной зоне, г – апикальная поверхность эпидермальных клеток с точечным и закрученным рисунком. 6 – пластинчатый кольцевидный рисунок; ост. обозначения см. на рис. 2. Масштаб, мкм: а – 30, б – 5, в – 20, г – 5.

Высокие плоские пластинчатые разрастания заменили гребни апикальной мембраны клеток. В отличие от других форм модификации, они сохранили кольцевидное распределение, соответствующее расположению гребней. Пластины могут варьировать по высоте на соседних клетках и в пределах одной клетки. Апикальная мембрана этих клеток с высокими кольцевидно расположенными пластинами может выгибаться, образуя возвышения, или прогибаться внутрь, образуя углубления (рис. 3а, 3б). Клетки с пластинчатыми разрастаниями образуют скопления в коже латеральной и вентральной поверхности головы. Клетки с пластинчатым рисунком не перемежаются с клетками, имеющими фрагментарный или точечный рисунок (рис. 3).
Клетки с фрагментарным рисунком особенно многочисленны в зонах кожи с вкусовыми почками. Разнообразные клетки этого типа окружают все выявленные вкусовые почки погружённого и непогружённого типов. Клетки с модифицированной апикальной мембраной составляют эпителий вкусовых сосочков и эпителий кожи, окружающей вкусовые сосочки. Скопления этих модифицированных клеток многочисленны на поверхности верхней и нижней губ, в коже около губ, в эпителии усика (рис. 2б, 2г). В коже гулярной зоны и около губ часто встречаются скопления эпидермальных клеток с модификацией точечного типа. Многочисленные клетки с фрагментарным и точечным рисунком часто располагаются рядом (рис. 3в, 3г).
Скопления клеток с пластинчатыми структурами часто располагаются в коже около и каудальнее ротовых зон, где встречаются редкие вкусовые почки, а также в зонах без вкусовых почек. Это значительные площади кожи латеральной стороны головы ростральнее обонятельного мешка, кожа каудального края губ, кожа вентральной стороны рядом с гулярной зоной. В коже головы сохранились также значительные площади с клетками обычного типа с обычным характерным кольцевидным рисунком на апикальной поверхности.
В ротовой полости оперированных рыб эпителий претерпел те же изменения, что и наружный эпителий. Обкладочные клетки вкусовых почек, клетки вкусовых сосочков и клетки эпителия между вкусовыми сосочками имеют модифицированную апикальную поверхность с фрагментарным рисунком (рис. 2в). Клетки с пластинчатыми разрастаниями не выявлены. В зонах эпителия без вкусовых почек находятся скопления клеток с обычным кольцевидным рисунком.
Сравнение эпидермальных клеток у четырёх оперированных рыб показало отсутствие индивидуальных различий в структурных изменениях на апикальной поверхности во всех исследованных участках кожи головы и эпителия ротовой полости.
ОБСУЖДЕНИЕ
Эпидермальные клетки кожи рыб разных видов имеют однотипную классическую структуру апикальной поверхности, которая несёт многочисленные кольцевидные гребни (Devitsina, 2005; Девицина, Головкина, 2008, 2011, 2018). Такая структура поверхности эпидермальных клеток генетически закреплена и у наземных позвоночных животных (Елисеев и др., 1972; Трубачёв, Василевский, 1976). Эпидермальные клетки кожи, в частности их апикальная поверхность, выполняют пограничные функции, защищая организм от непосредственных воздействий внешней среды. Важным свойством эпидермальных клеток является также их способность передавать к многочисленным рецепторным структурам (свободные нервные окончания, клетки Меркеля и др.), находящимся в коже, внешние механические воздействия различного происхождения (Уолл, 1964; Whitear, 1971; Kasumyan, 2011).
Экспериментально показано, что у карповых рыб тактильная рецепция особенно хорошо развита в коже головы, где она имеет зональное распределение. Околоротовые зоны, которые включают поверхность губ и кожи каудальнее губ, обладают наиболее высокой тактильной чувствительностью (Девицина, Лапшин, 2020). Морфологическое изучение поверхности клеток кожи головы в зонах с различной тактильной чувствительностью у интактного карпа показало, что эпидермальные клетки на своей апикальной поверхности во всех зонах имеют сходное кольцевидное расположение гребней. Однако эпидермальные клетки в тех же тактильных зонах головы у сенсорно депривированных рыб видоизменены. На апикальной поверхности клеток появились кольцевидно расположенные пластины разного размера. Появление высоких пластинчатых разрастаний, которые увеличивают площадь апикальной поверхности и могут быть подвижны, вероятно, способствует повышению чувствительности кожи к стимулам касательного и гидродинамического характера. Особенно высокий уровень осязательной чувствительности кожи обнаружен у астианакса Astyanax fasciatus − обитателя пещерных водоёмов (Windsor et al., 2008). Подобные морфологические изменения эпидермальных клеток обнаружены и у слепых пещерных особей астианакса A. mexicanus. В отличие от карпа у астианакса пластинчатые разрастания на эпидермальных клетках встречаются как на поверхности кожи, так и во вкусовых зонах ротовой полости (Девицина, Головкина, 2018). По-видимому, в условиях сенсорного дефицита у рыб разных видов в апикальной мембране эпидермальных клеток могут происходить сходные морфологические изменения и, в том числе, изменения адаптивной механосенсорной направленности.
Вместе с тем распределение и характер модификации апикальной мембраны клеток могут иметь и видовую специфику. Так, сенсорно депривированные карпы отличаются от слепого астианакса изменениями в коже вкусовых зон наружной и ротовой локализации. Отличия заключаются в полном исчезновении у карпа обычного кольцевидного рисунка на эпидермальных клетках во вкусовых зонах и замене его на рисунок фрагментарного или точечного типа. У слепого астианакса чаще появляются пластинчатые модификации поверхности эпидермальных клеток. Столь существенные изменения структуры апикальной мембраны могут изменять и её свойства, такие как упругость, эластичность, подвижность, связанные с выполнением различных функций кожей у рыб (Романенко, 2001).
У карпа зоны эпидермиса с различным характером чувствительности и, возможно, с различной иннервацией, имеют различное распределение модифицированных эпидермальных клеток. Так, в зонах с повышенной тактильной чувствительностью и рядом с ними находятся большие скопления клеток с пластинчатой модификацией апикальной поверхности (Девицина, Лапшин, 2020). В зонах, содержащих скопления вкусовых почек, эпидермальные клетки имеют на апикальной поверхности модификации фрагментарного рисунка. Особенности распределения эпидермальных клеток с различными структурными изменениями апикальной поверхности в коже головы и в ротовой полости карпа дают основания предполагать их функциональную и, возможно, адаптивную направленность.
Ранее было показано, что хроническая депривация сенсорных систем дистантного действия у рыб приводит к морфологической гипертрофии рецепторного аппарата контактного действия – вкусовых рецепторов наружной и ротовой локализации (Девицина, 2006; Девицина и др., 2015). Обнаружено также, что у карпа с хронической аносмией повышается чувствительность кожной хеморецепции и вкусовой рецепции (Касумян, Марусов, 2007; Chervova, Lapshin, 2010; Kasumyan, Sidorov, 2010).
Результаты настоящей работы показали, что у карпа хроническая депривация основных систем дистантного действия (зрительной и обонятельной) приводит к значительным морфологическим изменениям структуры апикальной поверхности эпидермальных клеток в коже головы. При этом разные зоны головы различаются характером морфологических изменений эпидермальных клеток. Это даёт основание предполагать существование и функциональных различий между зонами разной локализации. Известно, что околоротовые зоны кожи у карпа имеют особенно интенсивную иннервацию со стороны тройничного нерва, который является проводником полисенсорной информации, в том числе и тактильной рецепции (Whitear, 1971; Belousova et al., 1983; Puzdrowski, 1987). Одновременная активация рецепторных структур и центральных проекций вкусовой и тактильной чувствительности свидетельствует о взаимодействии этих систем и подтверждается рядом морфологических и физиологических исследований (Herrick, 1906; Kiyohara et al., 1986; Marui et al., 1988; Devitsina, 2003; Patterson, 2007).
Значительные изменения структуры апикальной мембраны эпидермальных клеток в условиях зрительного и ольфакторного дефицита можно рассматривать как отражение сложной компенсаторной реакции со стороны тактильной рецепции. По-видимому, у рыб эпидермальные клетки обладают высоким потенциалом морфологической адаптивной изменчивости. Морфологические признаки активации тактильных функций кожи у зрительно и ольфакторно депривированных рыб свидетельствуют о взаимодействии кожной тактильной системы с сенсорными зрительной и обонятельной системами. Взаимодействие этих сенсорных систем дистантного и контактного действия проявляется и у наземных позвоночных (Смит, 2005; Patterson, 2007). У слепых рыб при отсутствии зрительной рецепции ориентация в пространстве осуществляется с помощью боковой линии, а также возможна при активации осязательной, тактильной чувствительности эпидермальных клеток с различными формами модификации их апикальной мембраны.
Таким образом, можно полагать, что зрительная и обонятельная депривация рыб вызывает значительные морфологические и, возможно, функциональные изменения эпидермальных клеток разной локализации. Изменения могут иметь адаптивный характер и сохраняться в течение длительного времени. Выявленные изменения эпидермальных клеток проявляются в коже наружных сенсорных зон и в эпителии ротовой полости. Изменения морфологии апикальной поверхности эпидермальных клеток в условиях хронического выключения зрительной и обонятельной рецепции у рыб отражают компенсаторную активацию системы кожной тактильной рецепции.
Список литературы
Девицина Г.В. 1990. Исследование морфологии хемосенсорных систем // Сенсорная физиология морских рыб (методические аспекты). Мурманск: Апатиты. С. 7–10.
Девицина Г.В. 2006. Адаптивная изменчивость рецепторного отдела вкусовой системы карпа (Cyprinus carpio L.) после хронической аносмии // Журн. эволюц. биохимии и физиологии. Т. 42. № 6. С. 589–594.
Девицина Г.В., Головкина Т.В. 2008. Морфология вкусового аппарата ротовой полости наваги Eleginus navaga L. и трески Gadus morhua maris albi Der. (Gadidae, Teleostei) // Сенсорные системы. Т. 22. № 1. С. 70−86.
Девицина Г.В., Головкина Т.В. 2011. Вкусовой аппарат у молоди радужной форели Parasalmo (= Oncorhynchus) mykiss Walb. // Вопр. ихтиологии. Т. 51. № 1. С. 113−123.
Девицина Г.В., Головкина Т.В. 2018. Структурная организация вкусового аппарата у харациновых рыб (Characidae, Teleostei) // Вопр. ихтиологии. Т. 58. № 1. С. 53–66. https://doi.org/10.7868/S0042875218010071
Девицина Г.В., Лапшин Д.Н. 2016. Тактильная рецепция наружных покровов головы серебряного карася (Carassius auratus gibelio) // ДАН. Т. 466. № 4. С. 493–495. https://doi.org/10.7868/S0869565216040253
Девицина Г.В., Лапшин Д.Н. 2020. Неинвазивное электрофизиологическое исследование тактильной чувствительности у карповых рыб (Cyprinidae) // Журн. эволюц. биохимии и физиологии. Т. 56. № 4. С. 303−313. https://doi.org/10.31857/S004445292004004X
Девицина Г.В., Марусов Е.А. 2007. Взаимодействие сенсорных систем и пищевое поведение рыб // Успехи соврем. биологии. Т. 127. № 4. С. 387–395.
Девицина Г.В., Ружинская Н.Н., Гдовский П.А. 2010. Влияние хронической аносмии на морфофункциональные показатели первичных центров вкусовой системы карпа (Cyprinus carpio L.) // Журн. эволюц. биохимии и физиологии. Т. 46. № 4. С. 321−329.
Девицина Г.В., Головкина Т.В., Лобырев Ф.С. 2015. Взаимодействие интраоральной вкусовой субсистемы с сенсорными системами дистантного действия у рыб // Сенсорные системы. Т. 20. № 3. С. 201– 212.
Елисеев В.Г., Афанасьева Ю.И., Копаева Ю.Н., Юрина Н.А. 1972. Гистология. М.: Медицина, 616 с.
Ильинский О.Б. 1967. Вопросы физиологии сенсорных систем: механорецепторы. Л.: Наука, 84 с.
Касумян А.О., Марусов Е.А. 2007. Хеморецепция у хронически аносмированных рыб: феномен компенсаторного развития вкусовой системы // Вопр. ихтиологии. Т. 47. № 5. С. 684–693.
Павлов Д.С., Касумян А.О. 1990. Сенсорные основы пищевого поведения рыб // Там же. Т. 3. Вып. 5. С. 720–732.
Романенко Е.В. 2001. Гидродинамика рыб и дельфинов. М.: Тов. науч. изд. КМК, 411 с.
Смит К. 2005. Биология сенсорных систем. М.: Бином. Лаборатория знаний, 583 с.
Трубачёв В.В., Василевский Н.Н. 1976. Кожная сенсорная система // Физиология сенсорных систем / Под ред. Батуева А.С. М.: Медицина. С. 232–260.
Уолл П. 1964. Две системы передачи кожных ощущений // Теория связи в сенсорных системах. М.: Наука. С. 166–180.
Belousova T.A., Devitsina G.V., Malyukina G.A. 1983. Functional peculiarities of fish trigeminal system // Chem. Senses. V. 8. № 2. P. 121–130.
Chervova L.S., Lapshin D.N. 2010. The effect of olfactory deprivation on the thresholds of sensitivity of skin chemoreceptors of carp (Cyprinus carpio L.) // J. Ichthyol. V. 50. № 11. P. 1030−1035. https://doi.org/10.1134/S0032945210110081
Devitsina G.V. 2003. On the chemosensory systems interaction in fish // Ibid. V. 43. Suppl. 1. № 8. P. S214–S227.
Devitsina G.V. 2005. Comparative morphology of intraoral taste apparatus in fish // Ibid. V. 45. Suppl. 2. S286–S306.
Herrick C.J. 1906. On the centers for taste and touch in the medulla oblongata of fishes // J. Comp. Neurol. V. 16. P. 403–439.
Kasumyan A.O. 2011. Tactile reception and behavior of fish // J. Ichthyol. V. 51. № 11. P. 1035–1103.
Kasumyan A.O., Sidorov S.S. 2010. Behavior of gustatory testing of food objects in the carp (Cyprinus carpio) in the norm and at chronic anosmia // Ibid. V. 50. № 11. P. 1043–1059.
Kiyohara S., Houman H., Yamashita S. et al. 1986. Morphological evidence for a direct projection of trigeminal nerve fibers to the primary gustatory center in the catfish, Plotossus anguilaris // Brain Res. V. 379. P. 353–368.
Marui T., Caprio J., Kiyohara S., Kasahara Y. 1988. Topographical organization of taste and tactile neurons in the facial lobe of the sea catfish, Plotosus lineatus // Ibid. V. 446. P. 178–182.
Patterson M. 2007. Seeing with the hands, touching with the eyes: vision, touch and the enlightenment spatial imaginary // Senses Soci. V. 1. № 2. P. 225–243.
Puzdrowski R.L. 1987. The peripheral distribution and central projections of the sensory rami of the facial nerve in the goldfish, Carassius auratus // J. Comp. Neurol. V. 259. № 3. P. 382–392. https://doi.org/10.1002/cne.902590306
Whitear M. 1971. Cell specialization and sensory function in fish epidermis // J. Zool. V. 163. P. 237–264.
Windsor S.P., Tan D., Montgomery J.C. 2008. Swimming, kinematics and hydrodynamics imaging in the blind Mexican cave fish (Astyanax fasciatus) // J. Exp. Biol. V. 211. P. 2950–2959. https://doi.org/10.1242/jeb.020453
Дополнительные материалы отсутствуют.
Инструменты
Вопросы ихтиологии